
辣椒胶孢炭疽菌遗传转化体系的建立
2019-09-04 09:34曾泉满益龙陈岳张鑫张卓李成刚杜娇张德咏刘勇谭新球
植物保护 2019年4期
曾泉 满益龙 陈岳 张鑫 张卓 李成刚 杜娇 张德咏 刘勇 谭新球


摘要 由辣椒胶孢炭疽菌Colletotrichum gloeosporioides导致的辣椒炭疽病是辣椒生产上最为严重的真菌病害之一。本文以辣椒胶孢炭疽菌CSLL11为供试菌株,采用PEG-CaCl2介导的原生质体转化法,将含有潮霉素B抗性基因和eGFP表达基因的DNA片段成功转入辣椒胶孢炭疽菌的原生质体中,获得了稳定表达绿色荧光的转化子,从而成功建立了辣椒胶孢炭疽菌的遗传转化体系。试验结果表明,可有效筛选阳性转化子的潮霉素B浓度为500 mg/L;PCR及Southern blot结果显示,eGFP表达基因已单拷贝整合至辣椒胶孢炭疽菌转化子的基因组中;使用荧光显微镜观察第一代及继代培养后的转化子,菌丝与分生孢子均表现出强烈的绿色荧光信号,说明GFP能在转化子中稳定遗传;将转化子与野生型菌株相比,菌落形态、生长速率及致病力水平无明显差异。本研究建立了辣椒胶孢炭疽菌遗传转化体系并获得了稳定表达绿色荧光蛋白的转化子,对辣椒炭疽菌与寄主互作的研究及病害防治具有重要意义。
关键词 辣椒炭疽病菌; 遗传转化; GFP; 生长; 致病性
中图分类号: S 436.418
文献标识码: A
DOI: 10.16688/j.zwbh.2019037
辣椒为茄科辣椒属的一年生草本植物[1],是我国种植面积最大的蔬菜作物之一,目前种植面积已经超过146.7万hm2[2]。辣椒炭疽病是辣椒生产上最为严重的真菌病害之一,分布广,危害重,传播快,可引起辣椒幼苗死亡、落叶和果实腐烂,严重影响辣椒的产量和品质[35]。
辣椒炭疽病的病原菌为胶孢炭疽菌Colletotrichum gloeosporioides,属于半知菌亚门刺盘孢属真菌[6]。类似于其他真菌,炭疽菌在冬天或者环境胁迫条件下能产生大量的微菌核在土壤中休眠,待条件适宜时,微菌核产生大量孢子,经过雨水传播到寄主表面,一旦孢子接触寄主后,萌发形成附着胞,病原菌借助其巨大的膨压穿透寄主表皮,然后在寄主细胞内形成侵染菌丝再扩展到相邻细胞,形成病斑,同时在病斑上产生大量的孢子,开始下一轮的侵染[7]。尽管炭疽病菌的侵染循环已然明了,但由于炭疽菌种类繁多,同属不同种的炭疽病菌间生物学性状差异巨大,胶孢炭疽菌各寄主专化型与相应寄主间的互作机制复杂,因此辣椒胶孢炭疽菌的致病机理还不清楚,其与寄主的互作机理尚不明确,因此,有必要针对辣椒胶孢炭疽菌建立一套有效的研究体系。遗传转化是研究病原菌生长发育与致病分子机制的基本方法和手段。在前期对炭疽菌的研究中,包括PEG-CaCl2介导的原生质体转化技术[89]、根癌农杆菌介导的孢子转化技术[1011],以及电穿孔转化法[12]在内的遗传转化操作已得到广泛的应用,但是由于胶孢炭疽菌各寄主专化型间培养性状方面存在较大差异,因此以上转化体系在辣椒炭疽病菌上还未得到成功应用。因此,建立一套高效、稳定的辣椒炭疽病菌遗传转化体系,有助于进一步深入研究其生物学、致病分子机理以及挖掘潜在的药剂分子靶标,为辣椒炭疽病的防治提供理论依据。
1 材料与方法
1.1 试验材料
1.1.1 菌株及质粒
辣椒炭疽病菌菌株CSLL11分离自湖南省长沙县榔梨镇,采集典型症状标本,经单孢分离,病原鉴定后保存备用。含潮霉素B抗性的质粒SK1044由江苏省农业科学院刘永锋研究员馈赠。
1.1.2 培养基
PDA培养基:购自索莱宝Solarbio公司制备的不含抗生素的马铃薯葡萄糖培养基,46 g于1 L ddH2O,溶解,121℃高压灭菌20 min。
TB3培养基:酵母提取物3 g,酸水解酪蛋白3 g,蔗糖200 g,加ddH2O定容至1 L,121℃高压灭菌20 min,固体培养基加琼脂1.5 g/200 mL(m/V)。
1.1.3 试剂
裂解酶(lysing enzyme)、聚乙二醇(PEG)和潮霉素(hygromycin B)购自美国Sigma aldrich公司。
1×STC缓冲液:蔗糖100 g,Tris-HCl(50 mmol/L) 3.028 5 g(pH=8.0),CaCl2(50 mmol/L)2.774 5 g;加ddH2O至500 mL,121℃高压灭菌20 min。
PTC缓冲液:称量PEG 4000 120 g,溶解于1×STC缓冲液中,定容至200 mL,121℃高压灭菌20 min。
5×YEG:酵母提取物5 g,葡萄糖10 g,加ddH2O溶解,定容至1 L,121℃高压灭菌20 min。
酶解液配制:细胞壁破壁酶0.3 g,完全溶解于0.7 mol/L NaCl后定容至30 mL,用0.22 μm细菌过滤器过滤除菌。
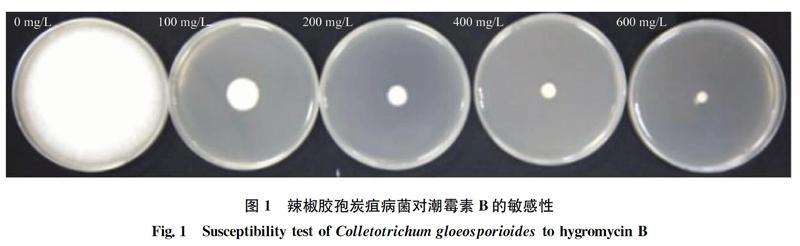
1.2 辣椒胶孢炭疽病菌对潮霉素的敏感性测定
将辣椒胶孢炭疽病菌菌株CSLL11接种在PDA培养基培养5 d后,用打孔器沿菌落边缘打取菌丝块,转接到潮霉素浓度分别为0、100、200、400 mg/L和600 mg/L的PDA平板上, 26℃恒温培养箱中培養7 d,观察生长情况,每个浓度设置3次重复。
1.3 辣椒胶孢炭疽病菌的遗传转化
1.3.1 原生质体的制备
培养由单个分生孢子再生的菌落,切取菌丝块于PDA平板上生长1~2 d,切取菌丝块,尽量切碎,置于约80 mL 5 × YEG的液体培养基中,28℃、150 r/min振荡培养16 h;用1~2层Miracloth过滤收集菌丝,无菌水冲洗两遍,用吸水纸将残留水分吸干,将菌丝置于盛有25 mL酶液的50 mL离心管中酶解。离心管水平放置,30℃、60 r/min振荡1.5~2 h酶解;向酶解液内加少许0.7 mol/L NaCl溶液,轻轻摇晃,使用三层灭菌擦镜纸过滤,用0.7 mol/L NaCl溶液轻轻冲洗1~2次,去残渣,收集滤液于50 mL离心管中,3 600 r/min、4℃离心10 min;小心弃上清,加10~20 mL 1×STC悬浮沉淀,用剪去尖头的枪头轻轻吹打,然后3 000 r/min、4℃离心5 min,重复上述步骤两次,加适量(约300 μL)1×STC溶液悬浮,计数,使原生质体终浓度为108个/mL,分装为150 μL/管,立即进行转化操作。
2.4 GFP轉化菌株及野生型菌株的生物学特性差异
2.4.1 GFP转化菌株及野生型菌株的生长情况
采用菌饼法检测菌株生长速率,并利用摇菌法进行产孢量试验。试验结果表明转化菌株与野生型菌株菌落形态及生长速率无明显差异(表1)。转化菌株平均产孢量为0.8×105~1.8×105个/mL,野生型菌株的产孢量为0.8×105~1.4×105个/mL,没有明显差异。
2.4.2 GFP转化菌株的致病性测定
以5×105个/mL 分生孢子接种辣椒果实进行致病性测定,接种6 d后调查致病情况。结果表明,转化菌株及野生型菌株接种的辣椒均出现向内凹陷的病斑,而未接种病原菌的对照未发病(图6)。
3 讨论
PEG-CaCl2介导的原生质体转化方法具有转化效率高、操作简便、体系成熟等特点,在稻瘟病菌、禾谷镰刀菌等丝状真菌遗传转化研究中应用非常成熟[1314],在杨树、苹果、梨炭疽菌的遗传操作中也有报道,但是在辣椒炭疽病菌中还未报道。胶孢炭疽菌寄主繁多,病菌根据寄主范围而分化为各种类型,各专化型间培养性状、形态及致病力水平存在较大差异。本研究在前期胶孢炭疽菌转化体系的基础上,进一步对辣椒炭疽病菌的转化条件摸索优化,通过PEG-CaCl2介导的原生质体转化方法把含GFP序列的DNA片段转入到辣椒炭疽菌中,并且获得稳定的绿色荧光表达菌株,从而在辣椒炭疽菌中成功建立了高效、稳定的遗传转化体系,为后续研究奠定了基础。
原生质体的制备在辣椒炭疽菌遗传转化的过程中起着关键作用,其质量和数量直接影响转化的效率,而菌株专化型、菌株培养时间等因素均决定着原生质体的质量。辣椒炭疽菌在固体平板及液体培养条件下均易产生分生孢子,当大量分生孢子混杂在原生质体中时,不仅会导致转化效率降低,还会影响转化子的再生。在梨炭疽病菌中,分生孢子在CM液体培养基生长至24 h后,继续转接到新的CM液体培养基中扩培24 h[9],而本研究缩短了培养时间,新鲜菌丝块置于5×YEG的液体培养基中振荡培养16 h后即用于转化,从而避免了分生孢子对转化过程的影响。
目前,真菌遗传操作过程中的选择标记有潮霉素、博莱霉素、G418等,为真菌的基因转入、敲除提供了多种选择[1516]。本研究在尚不明确辣椒炭疽菌对潮霉素B敏感性的前提下,首先开展了潮霉素B的敏感性测定,经检测发现当潮霉素B浓度为500 mg/L时,能显著抑制辣椒炭疽菌的生长,表明此浓度适合作为遗传转化选择浓度。
本研究所用的转化片段以农杆菌质粒SK1044为模板扩增获得,含有增强型绿色荧光蛋白GFP及来源于异旋孢腔菌的强启动子GPDh,目前SK1044表达载体多在镰刀菌中使用[17],本研究首次在辣椒炭疽菌中使用了该启动子,获得的转化菌株荧光强度高且较一致,说明GPDh启动子可以在辣椒炭疽菌中高水平启动目的基因的表达。此外,本研究转化过程中使用的是含有T-DNA上下臂、GFP基因及hygR基因的DNA片段,相比转化质粒更加方便且阳性率达到了100%。
在辣椒炭疽病菌遗传转化体系成功建立的同时发现,转化子通过3代培养仍可以表达出明亮绿色荧光,说明GFP基因已整合至转化子基因组中并且能够稳定遗传。同时,考虑到PEG介导的转化方法具有随机插入的特点,我们对获得的转化子进行了Southern blot验证,所选的6个转化子均为单拷贝插入,单拷贝插入比例达到100%。在对其生长和致病过程中的研究发现,尽管转入了外源片段,但是转化子的生物学表型与野生型较为一致,表明转入的GFP片段并不影响辣椒炭疽菌自身的生长发育,因此可用于后续研究辣椒炭疽菌与寄主互作关系。
本研究摸索了辣椒炭疽病菌的原生质体转化条件,建立了高效、稳定的PEG-CaCl2介导的辣椒炭疽病菌遗传转化体系,通过转入绿色荧光表达基因获得带有绿色荧光标记的转化菌株,并且其生长形态、生长速率及致病性与原始菌株没有差异,该结果将为挖掘辣椒炭疽病菌致病相关基因提供强大的技术支持,从而为后续的辣椒炭疽菌与寄主互作的机制研究奠定了良好基础。
参考文献
[1] 张莉.促进辣椒产业进一步发展——我国辣椒制品行业现状及发展趋势[J].产品可靠性报告,2008,10:3637.
[2] 黄任中,黄启中,吕中华,等.我国干制辣椒产业现状及发展对策[J].中国蔬菜,2015(2):911.
[3] 汪爱娥,丁克坚,马珂.辣椒炭疽病的研究进展[J].安徽农业科学,2005,33(3):508509.
[4] 郭明程,李保同,汤丽梅,等.爵床提取物对辣椒炭疽病菌的抑制作用及其苗期防效[J].植物保护,2013,39(4):119123.
[5] 胡彬,王晓青,梁铁双.辣椒主要病虫害化学防治技术[J].中国蔬菜,2017(4):8792.
[6] 孙春英,毛胜利,张正海,等.辣椒抗炭疽病遗传与育种研究进展[J].园艺学报,2013,40(3):579590.
[7] THAN P P, PRIHASTUTI H, PHOULIVONG S, et al. Chilli anthracnose disease caused by Colletotrichum species [J]. Journal of Zhejiang University-Science B, 2008, 9(10):764778.
[8] 李思蒙,王永林,黄冬辉,等.杨树炭疽病菌原生质体遗传转化的建立及绿色荧光蛋白的表达[J].林业科学,2013,49(5):121127.
[9] 杜艳,刘永锋,常有宏,等.梨炭疽病菌原生质体遗传转化体系的建立及GFP标记菌株的获得[J].江苏农业学报,2017,33(2):295300.
[10]林春花,劉先宝,蔡吉苗,等.橡胶树尖孢炭疽菌绿色荧光蛋白(GFP)标记转化株的获得[J].热带作物学报,2009,30(10):14951500.
[11]LIN C H, CAI Z Y, SHI T, et al. The use of T-DNA tagging to isolate mutants of Colletotrichum gloeosporioides and Colletotrichum acutatum with reduced virulence against Hevea brasiliensisa[J]. Forest Pathology, 2013, 43(4):289296.
[12]ARMESTO C, MAIA F G M, DE ABREU M S, et al. Genetic transformation with the gfp gene of Colletotrichum gloeosporioides isolates from coffee with blister spot [J]. Brazilian Journal of Microbiology, 2012, 43(3):12221229.
[13]SWEIGARD J A, CHUMLEY F G, VALENT B. Disruption of a Magnaporthe grisea cutinase gene [J]. Molecular & General Genetics, 1992, 232(2):183.
[14]GU Qin, ZHANG Chengqi, LIU Xin, et al. A transcription factor FgSte12 is required for pathogenicity in Fusarium graminearum [J].Molecular Plant Pathology,2015,16(1):113.
[15]ZHOU Xiaoying, LI Guotian, XU Jinrong. Efficient approaches for generating GFP fusion and epitope-tagging constructs in filamentous fungi [J]. Methods in Molecular Biology, 2011, 722:199212.
[16]ZHOU Xiaoying, LIU Wende, WANG Chenfang, et al. A MADS-box transcription factor MoMcm1 is required for male fertility, microconidium production and virulence in Magnaporthe oryzae [J]. Molecular Microbiology, 2011, 80(1):3353.
[17]赵芹,张向向,邓渊钰,等.茎基腐亚洲镰孢菌侵染抗感小麦品种的组织病理学观察[J].植物病理学报,2017(2):6272.
(责任编辑:田 喆)
猜你喜欢
中学生天地(A版)(2020年3期)2020-04-10
故事作文·高年级(2020年3期)2020-03-17
山东农业科学(2016年11期)2016-12-17
江苏农业科学(2016年7期)2016-10-20
红领巾·萌芽(2016年1期)2016-09-10
中国动物保健(2015年4期)2015-10-21
天津农业科学(2015年9期)2015-09-02
英语学习·新锐空间(2013年3期)2013-05-23
少年科学(2009年12期)2009-07-07
时代风采(2009年14期)2009-03-23
