
草地贪夜蛾的飞行能力测定
2019-09-04 09:34葛世帅何莉梅和伟徐瑞斌孙晓婷吴孔明
植物保护 2019年4期
关键词:草地贪夜蛾
葛世帅 何莉梅 和伟 徐瑞斌 孙晓婷 吴孔明


摘要 迁飞是草地贪夜蛾Spodoptera frugiperda (J. E. Smith) 快速扩散的生物学基础。本研究对草地贪夜蛾1~13日龄成虫飞行参数的测定表明:1~5日龄的个体具有较强的飞行能力,3日龄达到峰值后逐渐下降。3日龄个体吊飞24 h,平均飞行距离、飞行时间和飞行速度分别为(29.21±1.54)km、(11.00±0.47)h和(2.69 ±0.10)km/h。1~11日龄成虫的振翅频率无显著差异,平均振翅频率范围为42.39~44.64 n/s。相同日龄雌、雄蛾的飞行距离和飞行时间均无显著差异,但1日龄雄蛾的平均飞行速度显著大于雌蛾。补充营养组成虫的飞行距离和飞行时间显著高于未补充营养组。研究结果为深入阐明草地贪夜蛾的飞行生物学和迁飞机制提供了科学基础。
关键词 草地贪夜蛾; 迁飞; 振翅频率; 飞行能力
中图分类号: S 435.132
文献标识码: A
DOI: 10.16688/j.zwbh.2019322
草地贪夜蛾Spodoptera frugiperda(J.E.Smith)原生于美洲的热带和亚热带地区,是当地重要的农业害虫[1]。该虫繁殖量大、生活周期短、寄主范围广,可为害玉米、水稻、棉花、高粱等多种农作物[24]。草地贪夜蛾自2016年入侵非洲以来,已发展成为一种世界性的重大农业害虫。其于2019年1月侵入我国云南省西南地区[5],然后迅速向东向北蔓延,截至6月中旬我国已有19个省(市、自治区)发现草地贪夜蛾为害玉米等作物。
草地贪夜蛾之所以能在我国甚至世界范围内快速蔓延,与其远距离迁飞行为密不可分。迁飞是昆虫长期进化过程中的一种生存策略,是对外界环境的适应行为,也是迁飞性害虫经常大面积暴发成灾的原因[67]。国外专家采用海捕、轨迹分析、雷达监测等多种研究技术,系统地证明了草地贪夜蛾的远距离迁飞行为[810]。研究迁飞昆虫的专家Nagoshi等利用草地贪夜蛾基因的单倍型种类和比值鉴定其虫源性质[1112],推测出草地贪夜蛾在美国的两条迁飞路线,明确了其在美国的迁飞规律。随后,Nagoshi等又利用草地贪夜蛾单倍型分子标记技术发现南美洲国家草地贪夜蛾具有非常高的种群遗传相似性,可能为同质种群[13]。利用昆虫雷达对草地贪夜蛾迁飞模式的监测表明,与多数迁飞性昆虫相似,迁飞活动主要与空中温度和风速相关,迁飞方向主要受到风向的影响,但并非完全随风迁移而是与风向成一定夹角[14]。吴秋琳等基于WRF模式的三维轨迹分析模拟了缅甸草地贪夜蛾虫源迁入我国的路径和可能降落区域,对该虫在我国的蔓延范围进行了准确有效的预测[1516]。
草地贪夜蛾已在缅甸等东南亚国家定殖,这些国家是草地贪夜蛾迁入我国的境外虫源地。在国内,云南、广西、广东、海南等适合草地贪夜蛾终年繁殖的热带和亚热带地区也形成了稳定的向北方广大地区迁飞为害的虫源基地。为了阐明草地贪夜蛾在我国的迁飞规律,有必要研究明确其飞行生物学习性。测定昆虫的飞行能力是研究昆虫飞行生物学的基础[17],普遍采用的方法是利用昆虫飞行磨吊飞[1819]。通过控制吊飞环境和试虫状态比较不同环境因素和昆虫生理因素下昆虫的飞行能力。昆虫的振翅频率与飞行能力有密切的关系,目前的昆虫雷达已能准确测定高空迁飞昆虫的振翅频率,并以此作为鉴别昆虫种类的一个重要参数[20]。对昆虫振翅频率的研究可促进对其飞行生物学的了解,还可为雷达自动识别昆虫种类提供数据参考。本文研究了草地贪夜蛾的飞行能力和振翅频率,旨在深入了解草地贪夜蛾的飞行生物学,为阐明迁飞机制提供科学依据。
1 材料与方法
1.1 供试虫源
2019年1月在云南省德宏州鲜食玉米田采集草地贪夜蛾2~5龄幼虫,带回养虫室内饲喂新鲜玉米叶片至其化蛹。待成虫羽化后饲喂10%的白糖水补充营养,产卵后收集卵块,幼虫孵化后置于养虫盒内用人工饲料饲养建立实验种群。实验种群幼虫化蛹后,区分雌雄并放入不同的培养皿中,培养皿内垫入卫生纸喷水保持湿度。成虫羽化后,将雌雄虫分别置于不同的养虫笼中,贴上标签注明羽化日期和性别供试验使用。幼虫和成虫饲养条件均为:温度25℃± 1℃,相对湿度60% ±10%,光周期为L∥D=16 h∥8 h。
1.2 仪器设备
使用的仪器设備包括昆虫飞行信息系统(FXMD-24-USB型,佳多科工贸股份有限公司)、频闪仪(phaser-strobe PBX型,美国蒙那多)、加湿器(Bear JSQ-C50X5)、空调(小天鹅)、温湿度计(HTC-1)。
1.3 试验方法
吊飞试验参考郭江龙等的方法并做部分调整[21]。选取健康、能正常飞行的成虫,装于离心管内并编号。试虫称重后用少量乙醚轻微麻醉,然后放于有凹槽的泡沫板上展翅,用小毛笔轻轻除去胸腹连接处的鳞毛,用自制吊环蘸取少许502胶粘到试虫胸腹交接处的体壁上,保持吊环与试虫虫体垂直,轻轻吹气使胶快速凝固。试虫苏醒后,轻轻吹气观察其振翅情况,剔除不能正常振翅或振翅不均匀的个体。粘虫完毕后,用大头针连接吊环,按照编号顺序插在泡沫板上置于吊飞室,适应10~15 min后用频闪仪测试每头试虫的振翅频率,重复测试3次。测试完成后将试虫按照顺序套接于飞行磨吊臂末端,保持吊环垂直于吊臂向下,使试虫绕飞行磨中心轴在水平方向上做切线旋转。在电脑飞行磨系统上登记试虫信息并且设置飞行磨采集终止时间后启动系统。吊飞期间保持室内完全黑暗,吊飞室温度25℃±1℃,相对湿度70%±5%。
1.3.1 测定不同日龄和性别成虫的飞行参数
成虫羽化后,每日饲喂10%蜂蜜水补充营养,选取1、3、5、7、9、11、13日龄的未交配雌、雄蛾测定其振翅频率和飞行能力,吊飞时间为24 h,不同日龄雌、雄蛾分别吊飞25~33头。
1.3.2 测定补充营养对成虫飞行参数的影响
选取3日龄未交配雌、雄蛾,设置10%蜂蜜水和清水两个处理,测定振翅频率和飞行能力,吊飞时间为24 h,雌、雄蛾各25~33头。
1.4 数据处理
使用统计分析软件DPS 7.05进行数据分析,所有数据均用平均值±标准误表示。使用单因素方差分析法对不同日龄成虫的飞行参数进行分析,若差异显著进行Tukeys HSD多重比较;使用双因素方差分析法分析日龄和性别对飞行的影响,然后进行Tukeys HSD多重比较。相同日龄、不同性别以及是否补充营养成虫之间的飞行参数采用均值t检验进行分析。数据分析前,先对原始数据进行异常值检验,剔除异常值后进行正态性检验,若不符合正态分布,将数据进行对数转换后再做统计分析,分析的显著水平为P<0.05,极显著水平为P<0.01。
2 结果与分析
2.1 不同日龄草地贪夜蛾成虫的飞行参数
总体上,不同日龄草地贪夜蛾成虫的振翅频率存在显著差异(F=4.38, df1=6, df2=384, P<0.01)。试虫振翅频率最高可达每秒61.33次,最低仅为每秒29.84次(表1)。1~11日龄成虫的振翅频率无显著差异,但13日龄成虫振翅频率明显降低。
24 h连续吊飞条件下,不同日龄成虫的飞行距离存在显著差异(F=60.77,df1=6,df2=376,P<0.01),最长飞行距离可达62.98 km(表1)。1~5日龄成虫飞行能力较强,3日龄达到峰值,平均飞行距离(29.21±1.54)km。此后随日龄增加飞行距离逐渐减少,至13日龄时平均飞行距离降到(5.58±0.69)km。
24 h连续吊飞条件下,不同日龄草地贪夜蛾成虫的飞行时间存在显著差异(F=56.77, df1=6, df2=380, P<0.01),最长飞行时间可达20.56 h(表1)。1日龄成虫即可进行较长时间的飞行,至3日龄达到峰值,平均飞行(11.00±0.47)h;3日龄以后,飞行时间随日龄增加而逐渐降低,至13日龄时飞行时间降为(2.91±0.36)h。
24 h连续吊飞条件下,不同日龄草地贪夜蛾成虫的飞行速度存在显著差异(F=4.64,df1=6,df2=384,P<0.01)。1~11日龄成虫的飞行速度无显著差异,以5日龄飞行速度最快(2.72±0.10)km/h,此后飞行速度降低,13日龄降至(2.15±0.08)km/h(表1)。
2.2 草地贪夜蛾雌蛾和雄蛾飞行能力比较
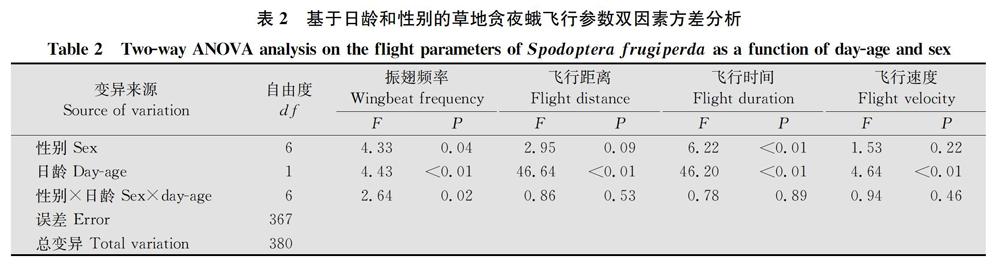
方差分析表明,不同性别草地贪夜蛾的振翅频率和飞行时间均有显著差异(振翅频率:F=4.33,P=0.04;飞行时间:F=6.22,P<0.01),且性别和日龄对振翅频率具有显著的交互作用(F=2.64,P=0.02)(表2)。11日齡雌蛾振翅频率显著高于雄蛾(t=4.63,P<0.01),但其他同一日龄雌、雄蛾间无显著差异。相同日龄雌、雄蛾的飞行距离和飞行时间均无显著差异,但1日龄雄蛾的平均飞行速度显著大于雌蛾(t=2.21,P=0.03)(图1)。
2.3 补充营养对草地贪夜蛾飞行参数的影响
连续吊飞24 h,取食10%蜂蜜水与清水的草地贪夜蛾成虫在振翅频率和飞行速度方面无显著差异,但飞行距离和飞行时间有显著性差异。取食蜂蜜水的成虫其飞行距离和飞行时间分别增加了1.29 km和0.85 h(表3)。
3 讨论
多种迁飞性的鳞翅目夜蛾科昆虫被证实有较强的飞行能力。昆虫的飞行能力受到多种因素的影响,包括温湿度、光照、营养、交配状态、振翅频率、日龄和性别等[2224]。迁飞性昆虫飞行生物学的研究一直是国内外的研究热点[6],准确获取迁飞性昆虫的飞行生物学参数对于阐明迁飞机制具有重要意义。
本研究通过对草地贪夜蛾实验种群飞行能力和振翅频率的测定证实了草地贪夜蛾具有较强的飞行潜力。进行24 h连续吊飞,草地贪夜蛾个体飞行时间最长可达20.56 h,飞行距离最远可达62.98 km。总体飞行能力与宽胫夜蛾Protoschinia scutosa相近,而略低于黄地老虎Agrotis segetum和甘蓝夜蛾Mamestra brassicae Linnaeus[21,2526]。不同日龄草地贪夜蛾的飞行能力有明显的差别,其1日龄成虫就有较强的飞行能力,3日龄达到峰值,之后随日龄增加其飞行能力下降,这与多种迁飞性昆虫包括棉铃虫Helicoverpa armigera、黄地老虎、甘蓝夜蛾、二点委夜蛾Athetis lepigone等的飞行能力变化规律相似[2122, 2627]。1日龄成虫飞行能力较3日龄稍弱的原因可能是飞行肌发育过程尚未完全结束。3日龄以后其飞行能力降低的原因可能是体内能量使用的重心开始转向生殖,供飞行用的能源物质不断减少且飞行肌干重下降使其不能维持较长时间的飞行[2829]。根据草地贪夜蛾1~5日龄具有较强飞行能力的现象,推测其迁飞发生在卵巢稚嫩期,符合“卵子发生飞行拮抗综合征”规律。虽然日龄是决定草地贪夜蛾飞行能力的一个因素,但除13日龄外,草地贪夜蛾的平均飞行速度并无显著差异,这表明日龄主要通过影响飞行时间影响飞行距离。1~11日龄草地贪夜蛾成虫的振翅频率没有显著性差异,此结果与不同日龄稻纵卷叶螟振翅频率变化规律一致[30],与棉铃虫、黏虫Mythimna separata和小地老虎Agrotis ipsilon振翅频率随日龄先上升后下降的趋势不同[31]。
24 h连续吊飞,相同日龄雌雄蛾的飞行距离和飞行时间均无显著差异。飞行速度方面,除1日龄雄蛾飞行速度显著大于雌蛾外,其他日龄雌雄间均无显著差异;振翅频率方面,除11日龄雌蛾振翅频率显著大于雄蛾外,其他日龄雌雄之间无显著差异。雌雄之间飞行能力变化趋势大致为初羽化时雄蛾比雌蛾拥有更强的飞行能力,此后二者飞行能力无显著差异。但3日龄以后,雌蛾在飞行距离和飞行时间上均优于雄蛾,可能随吊飞时间的增加其差异会逐渐显著[27]。1日龄雄蛾具有较强飞行能力的原因可能是自然界雌蛾比雄蛾先羽化1~2 d,雄蛾为了与雌蛾同时迁飞至相同区域或者为了更好地寻觅配偶进行交配,而需要在较短时间内拥有较强的飞行能力。
多项研究表明,成虫期取食不仅对昆虫的生殖活动有影响,而且对昆虫的飞行活动也具有显著的影响[3235]。本研究结果同样表明,补充营养后会显著增强草地贪夜蛾的飞行能力,主要体现在飞行距离和飞行时间的增加。另外,本研究还发现补充营养对增强草地贪夜蛾雄蛾飞行能力的作用更加明显,说明与雌蛾相比,雄蛾更依赖取食来增强自身的飞行能力,这可能是因为雌蛾体内储存较多的脂肪等能源物质,在没有取食的情况下仍然能够维持较强的飞行能力。
通过对草地贪夜蛾飞行生物学参数的测定,明确了不同日龄、性别和营养对草地贪夜蛾振翅频率和飞行能力的影响。但影响草地贪夜蛾飞行能力的因素还包括温度、湿度、交配和生殖等。此外,草地贪夜蛾飞行对其生殖力的影响,以及是否同褐飞虱Nilaparvata lugens、白背飞虱Sogatella furcifera和稻纵卷叶螟Cnaphalocrocis medinalis等迁飞性昆虫一样具有再迁飞的能力[3638]等,都还需要进一步研究。本研究所用虫源为室内实验种群,测试方法为室内飞行磨吊飞,这不能完全代表草地贪夜蛾野外种群在自然界的飞行活动。自然种群的迁飞活动更多地受到环境和行为因素的综合影响,这就需要采取室内、室外研究相结合,利用雷达监测等方法开展更为深入的研究工作。
参考文献
[1] SPARKS A N. A review of the biology of the fall armyworm[J]. The Florida Entomologist, 1979, 62(2): 8287.
[2] LUGINBILL P. The fall army worm [R]. USDA Technology Bulletin, 1928, 34: 91.
[3] MURU'A G, MOLINA-OCHOA J, COVIELLA C. Population dynamics of the fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae) and its parasitoids in northwestern Argentina [J]. Florida Entomologist, 2006, 89(2): 175182.
[4] MONTEZANO D. Host plants of Spodoptera frugiperda (Lepidoptera: Noctuidae) in the Americas [J]. African Entomology, 2018, 26(2): 286300.
[5] 楊学礼,刘永昌,罗茗钟,等.云南省江城县首次发现迁入我国西南地区的草地贪夜蛾[J].云南农业,2019(1):72.
[6] 程遐年.中国迁飞昆虫的研究进展[J].昆虫知识,1992,29(3):146149.
[7] DINGLE H, DRAKE V A. What is migration?[J]. BioScience, 2007, 57: 113121.
[8] SPARKS A N, JACKSON R D, CARPENTAR J E, et al. Insects captured in light traps in the Gulf of Mexico [J]. Annals of the Entomological Society of America, 1986, 79(1): 132139.
[9] WESTBROOK J K, NAGOSHI R N, MEAGHER R L, et al. Modeling seasonal migration of fall armyworm moths [J]. International Journal of Biometeorology, 2016, 60(2): 255267.
[10]WOLF W W, SPARKS A N, PAIR S D, et al. Radar observations and collections of insects in the Gulf of Mexico[M]∥DANTHANARAYANA W. Insect flight. Springer-Verlag, 1986: 221234.
[11]NAGOSHI R N, MEAGHER R L, FLANDERS K, et al. Using haplotypes to monitor the migration of fall armyworm (Lepidoptera: Noctuidae) corn-strain populations from Texas and Florida [J]. Journal of Economic Entomology, 2008, 101(3): 742749.
[12]NAGOSHI R N, FLEISCHER S, MEAGHER R L. Texas is the overwintering source of fall armyworm in central Pennsylvania: implications for migration into the northeastern United States [J]. Environmental Entomology,2009,38(6):15461554.
[13]NAGOSHI R N, GABRIELA MURU'A M, HAY-ROE M, et al. Genetic characterization of fall armyworm (Lepidoptera: Noctuidae) host strains in Argentina [J]. Journal of Economic Entomology, 2012, 105(2): 418428.
[14]WESTBROOK J K. Noctuid migration in Texas within the nocturnal aeroecological boundary layer[J]. Integrative and Comparative Biology, 2007, 48(1): 99106.
[15]吳秋琳,姜玉英,吴孔明.草地贪夜蛾缅甸虫源迁入中国的路径分析[J].植物保护,2019,45(2):16.
[16]吴秋琳,姜玉英,胡高,等.中国热带和南亚热带地区草地贪夜蛾春夏两季迁飞轨迹的分析[J].植物保护,2019,45(3):19.
[17]CAMPOSE W G, SCHOEREEDER J H, SPERBER C F. Does the age of the host plant modulate migratory activity of Plutella xylostella?[J]. Entomological Science, 2004, 7(4): 323329.
[18]KEES A M, HEFTY A R, VENETTE R C, et al. Flight capacity of the walnut twig beetle (Coleoptera: Scolytidae) on a laboratory flight mill[J]. Environmental Entomology, 2017, 46(3): 633641.
[19]刘莎,吕召云,高欢欢,等.昆虫飞行能力研究进展[J].环境昆虫学报,2018,40(5):3643.
[20]封洪强.雷达在昆虫学研究中的应用[J].植物保护,2011,37(5):113.
[21]郭江龙,付晓伟,赵新成,等.黄地老虎飞行能力研究[J].环境昆虫学报,2016,38(5):888895.
[22]吴孔明,郭予元.棉铃虫的飞翔活动[J].生态学报,1996,16(6):612617.
[23]袁瑞玲,杨珊,冯丹,等.温度、湿度、光照对桔小实蝇飞行能力的影响[J].环境昆虫学报,2016,38(5):903911.
[24]程登发,田喆,李红梅,等.温度和湿度对麦长管蚜飞行能力的影响[J].昆虫学报,2002,45(1):8085.
[25]赵胜园.宽胫夜蛾在渤海地区的迁飞行为研究[D].北京:中国农业科学院,2018.
[26]吴晓,付晓伟,赵新成,等.甘蓝夜蛾Mamestra brassicae Linnaeus飞行能力研究[J].应用昆虫学报,2016,53(3):595603.
[27]郑作涛,江幸福,张蕾,等.二点委夜蛾飞行行为特征[J].应用昆虫学报,2014,51(3):643653.
[28]JOHNSON C G. Migration and dispersal of insects by flight[M]. London: Methuen, 1969: 745763.
[29]SUN Beibei, JIANG Xingfu, ZHANG Lei, et al. Methoprene influences reproduction and flight capacity in adults of the rice leaf roller, Cnaphalocrocis medinalis (Guenée) (Lepidoptera: Pyralidae)[J]. Archives of Insect Biochemistry and Physiology, 2012, 82(1): 113.
[30]黄加诚.稻纵卷叶螟生殖和飞行的生理生态研究[D].武汉:华中农业大学,2014.
[31]施翔宇,封洪强,李建东,等.黏虫、棉铃虫和小地老虎振翅频率的比较[J].植物保护,2013,39(2):3135.
[32]江幸福,罗礼智,胡毅.成虫期营养对甜菜夜蛾生殖和飞行的影响[J].植物保护学报,2000,27(4):327332.
[33]吴孔明,郭予元.营养和幼期密度对棉铃虫飞翔能力的影响[J].昆虫学报,1997,40(1):5157.
[34]党志红,李耀发,安静杰,等.成虫期补充不同营养对二点委夜蛾实验种群繁殖力的影响[J].中国植保导刊,2017,37(8):1316.
[35]贾佩华,曹雅忠.小地老虎成虫的飞翔活动[J].昆虫学报,1992,35(1):5965.
[36]封传红,翟保平,张孝羲.褐飞虱的再迁飞能力[J].中国水稻科学,2001,15(2):125130.
[37]江远昆,翟保平.白背飞虱的再迁飞能力[J].昆虫学报,2004,47(4):467473.
[38]王凤英,张孝羲,翟保平.稻纵卷叶螟的飞行和再迁飞能力[J].昆虫学报,2010,53(11):12651272.
(责任编辑:杨明丽)
猜你喜欢
农家科技下旬刊(2019年10期)2019-11-29
植物保护(2019年5期)2019-11-27
植物保护(2019年5期)2019-11-27
现代农业科技(2019年19期)2019-11-21
现代农业科技(2019年19期)2019-11-21
农家科技(2019年10期)2019-11-13
农家科技(2019年7期)2019-10-08
农家科技(2019年7期)2019-10-08
农家科技(2019年8期)2019-09-19
南方农业·下旬(2019年4期)2019-08-26
