
西藏东达山3种嵩草属植物叶片解剖结构的比较
2019-08-30 07:07孙晓红杨春娇张大才
西南林业大学学报 2019年5期
孙晓红 杨春娇 张大才
( 西南林业大学西南地区生物多样性保育国家林业与草原局重点实验室,云南 昆明 650233)
青藏高原气候恶劣,低温,昼夜温差较大,降水具有明显的季节性、地域性差异[1]。嵩草属(Kobresia)隶属于莎草科(Cyperaceae),是藏东南地区高寒草甸的建群种和优势种[2]。因长期生长在青藏高原地区,嵩草属植物为适应高山环境,形成了明显的形态差异,并呈现出一定的变化机制,从而形成了特定的形态特征[3]。目前有关嵩草属植物的研究主要集中在种子发芽率和形态特征[4-5]、遗传多样性[6-7]、植物多样性的海拔分布格局[8-9]、植物群落特征[10-11]等方面,而对其叶片形态解剖结构方面的研究相对缺乏。叶片是植物进行气体交换、水分蒸腾、运输的门户,也是高等植物进行光合作用的重要器官[12]。在植物进化过程中,叶片形态解剖结构对环境因子的变化极为敏感[13]。叶片不同部位的解剖结构存在差异,而这些差异有可能与环境变化有关[14]。嵩草属植物为单子叶植物,叶片较长且生长气候恶劣,其完成叶片的生长需经历的气候变化较大。生长在严酷环境下的嵩草叶片不同部位解剖结构可能存在显著变化。东达山高寒草甸以大花嵩草(K. macrantha)、高山嵩草(K. pygmaea)和线叶嵩草(K. capillifolia)为优势种[15-16],因此,本研究拟比较3种嵩草属植物叶片解剖结构的差异性,以期为嵩草属植物形态学和分类学提供分类依据和参考资料,同时总结叶片不同部位解剖结构的差异和变化规律,进一步揭示嵩草属植物对高山环境的内在适应机理。
1 研究区概况
采样地位于西藏自治区昌都市左贡县东南部东达山垭口(28°30′~30°28′N,97°06′~98°36′E)附近。东达山是川藏318国道第2高垭口,海拔标高5 008 m,有“生命禁区”之称。该研究区域属于高原季风半湿润气候,气温年差较小,年平均气温为0~10 ℃。全年降水季节分布不均匀,蒸发量较大,相对湿度较小,年平均降水量400~600 mm,主要集中在5—10月份,约占全年降水量的80%,其余月份降水稀少且干燥寒冷。由于受到过度放牧的影响,高寒草甸趋于生境干旱化,具有耐辐射、耐旱、耐贫瘠特点的嵩草属植物成为青藏高原高寒草甸的优势种和建群种,同时其对该地区的生态系统平衡起着重要作用[17]。
2 材料与方法
2.1 研究材料
选取分布于东达山高寒草甸的大花嵩草、线叶嵩草、高山嵩草作为研究材料。在植物生长最旺盛的2017年8月中旬采样,选取分布生境相同的3种嵩草各15株,采集成熟叶片15片,就地截取的叶片迅速放入已准备好的固定液FAA中保存,带回实验室进一步处理。
2.2 研究方法
2.2.1 制片方法
采用常规石蜡连续切片法,将样品从FAA固定液中取出,然后用由蒸馏水、酒精、叔丁醇配制而成的脱水液,按照75%、85%、95%、100%的浓度梯度依次脱水,每级脱水2 h。抽气6 h左右,再用100%叔丁醇透明3次,每次3 h。最后用石蜡包埋、切片,切片厚度为12 μm,切片脱蜡后用苏木精染色,加拿大树胶封片[18]。
2.2.2 显微测量与观察
每种植物叶片按叶尖、叶中、叶基分别制成15张临时装片,每片叶片共45张临时装片,每个种共计675张临时装片,所有观察采用LeicaDM750型光学显微镜(徕卡公司,德国)。每张制片随机选取1个视野,观察并测量叶片厚度和叶片横切面面积。叶片厚度测量位置为近轴面主脉凹陷处与主脉方向对应的垂直距离(μm)。表皮细胞的大小以表皮细胞的长度和宽度来衡量。每张制片随机选取1个视野,每个视野选取连续10个表皮细胞测量;计数泡状细胞数量,并测量其面积。每张制片随机选取1个视野,测量1个维管束外鞘面积和内鞘面积,按照公式(1)~(2)计算机械组织占叶片横切面面积的比例(PML)和维管束占叶片横切面面积的比例(PBM)。

2.2.3 数据分析
基础数据采用Microsoft Excel 2010进行预处理,然后采用SPSS 21.0软件进行统计分析。嵩草属种间解剖结构差异选取叶基的测量值进行两两比较分析,采用单因素方差分析叶尖、叶中、叶基解剖结构的差异。
3 结果与分析
3.1 叶片解剖结构特征比较分析
由图1和表1可知,高山嵩草和线叶嵩草叶片横切面呈新月形,大花嵩草叶片横切面呈“V”字形;3种嵩草属植物叶片厚度为186.72~623.674 μm,其中线叶嵩草最厚,大花嵩草次之,高山嵩草最薄。解剖结构均由表皮、叶肉、叶脉组成,主要差异表现为大花嵩草近轴表皮主脉凹陷处存在泡状细胞,数量约为5个,面积为8 990.03 μm2,而高山嵩草和线叶嵩草叶片表皮细胞上无泡状细胞。
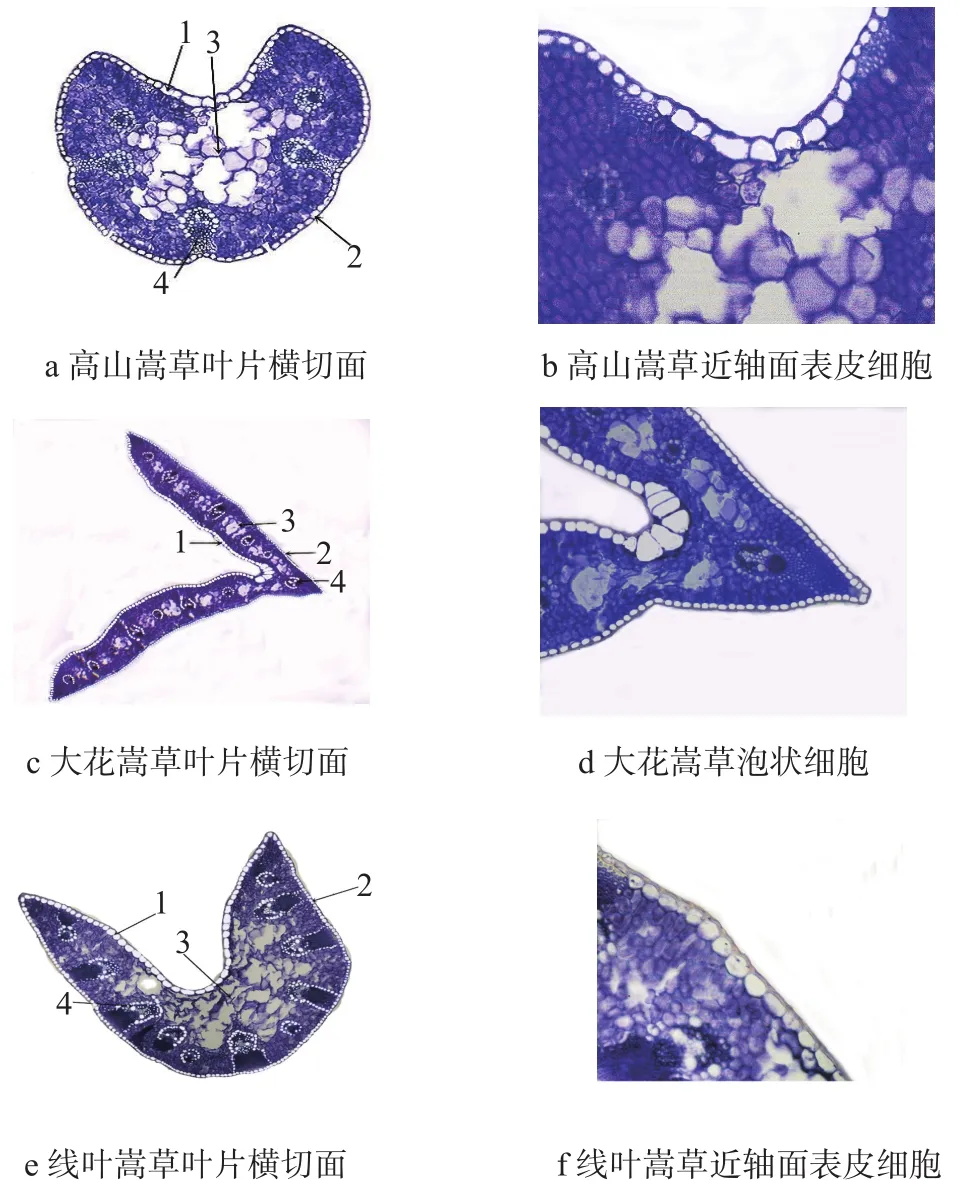
1为近轴面表皮细胞;2为远轴面表皮细胞;3为气腔;4为维管束。图 1 3种嵩草属植物叶片横切面结构Fig. 1 Leaf cross section structure of 3 species of Kobresia
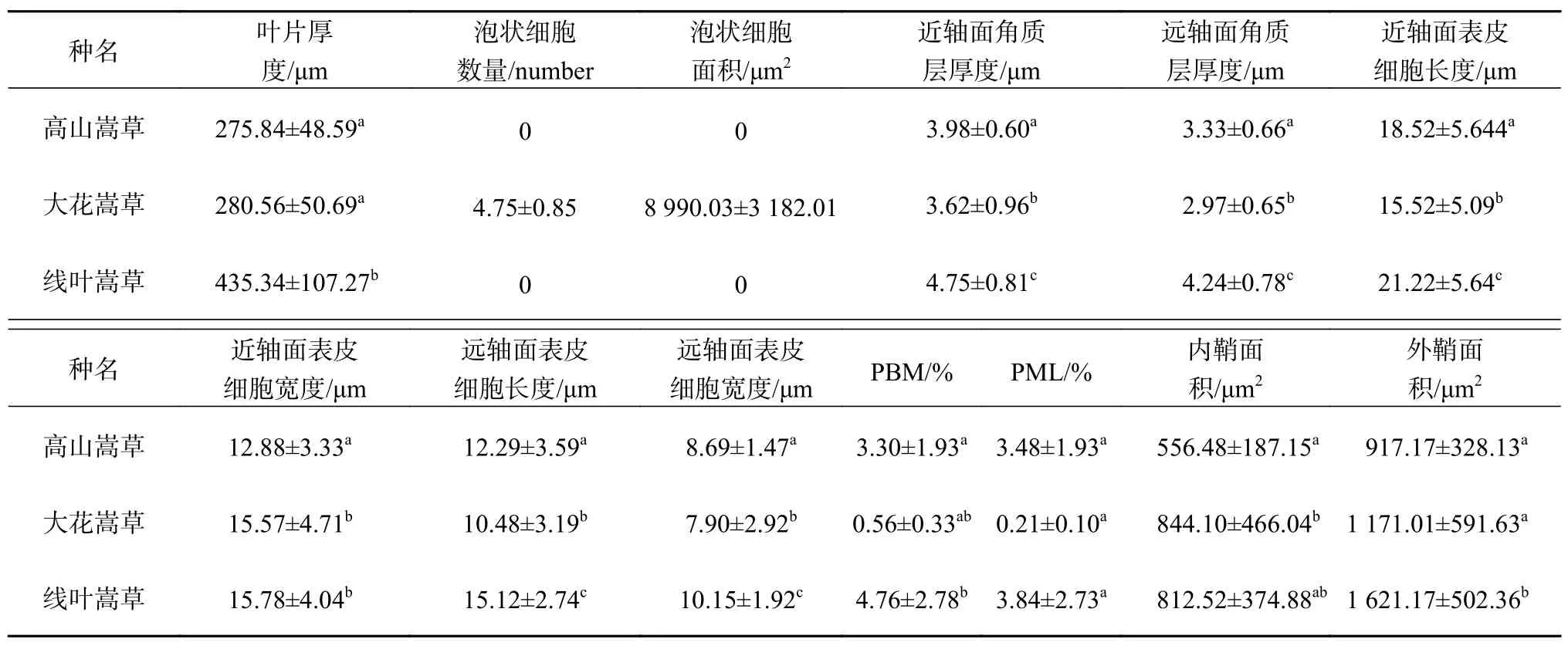
表 1 3种嵩草属植物叶片解剖结构特征Table 1 Characteristics of leaf anatomical structure of 3 species of Kobresia
3种嵩草属植物叶片近轴面表皮细胞角质层厚度均大于远轴面;线叶嵩草叶片近、远轴面角质层厚度最大,其次是高山嵩草,大花嵩草最小,且它们之间的差异显著(P < 0.05)。近、远轴面表皮细胞均由1层细胞组成,其中高山嵩草和大花嵩草近、远轴表皮细胞形状分别为长方形和近圆形,线叶嵩草近、远轴表皮细胞形状分别为长方形和椭圆形。近轴面表皮细胞均大于远轴面表皮细胞。3种嵩草叶片远轴面表皮细胞长度和宽度、近轴面表皮细胞长度差异显著(P <0.05),大花嵩草与线叶嵩草近轴面表皮细胞宽度差异不显著,与高山嵩草差异显著(P < 0.05)。
3种嵩草属植物叶肉没有栅栏组织与海绵组织的分化,但具有发育良好的气腔。高山嵩草和线叶嵩草在叶片中央形成气腔,大花嵩草的气腔和维管束相间排列,较小且形状不规则。高山嵩草和线叶嵩草主脉维管束在叶中位置靠近远轴面一侧,侧脉维管束有大、小之分。主脉与较大侧脉下方有成片机械组织,较小侧脉上下方均无机械组织;大花嵩草的维管束和气腔相间分布。线叶嵩草维管束和机械组织占叶片横切面面积的比例均最大,分别达到4.76%和3.84%;高山嵩草次之,分别为3.30%和3.48%;大花嵩草最小,分别为0.56%和0.21%。3种嵩草植物机械组织占叶片横切面面积的比例种间差异均不显著;大花嵩草与线叶嵩草维管束占叶片横切面面积的比例差异不显著,与高山嵩草差异显著(P < 0.05)。3种嵩草属植物维管束内鞘面积变化范围为 144.27~1 623.77 μm2,大花嵩草最大,其次是线叶嵩草,高山嵩草最小;3种嵩草属植物维管束外鞘面积变化范围为316.35~2 515.16 μm2,线叶嵩草最大,其次是大花嵩草,高山嵩草最小。
3.2 叶片不同部位解剖结构差异分析
由图2可知,大花嵩草和高山嵩草叶尖、叶中、叶基的叶片厚度差异极显著(P < 0.01),而线叶嵩草这3个部位叶片厚度差异不显著。3种嵩草3个部位叶片横切面面积变化一致,即叶中 > 叶基 > 叶尖。

图 2 3种嵩草植物叶片不同部位叶片厚度和叶片横切面面积的变化Fig. 2 Variation of leaf thickness and cross section of leaf area in different parts of leaves of 3 species of Kobresia
由图3可知,3种嵩草叶尖、叶中、叶基的近、远轴面角质层厚度差异极显著(P < 0.01);大花嵩草和线叶嵩草近、远轴面角质层厚度变化一致,即叶尖 > 叶中 >叶基,而高山嵩草近、远轴面角质层厚度表现为叶中 > 叶尖 > 叶基;大花嵩草和高山嵩草不同部位角质层厚度均值相近,而线叶嵩草叶尖部位角质层厚度显著大于叶中、叶基部位。
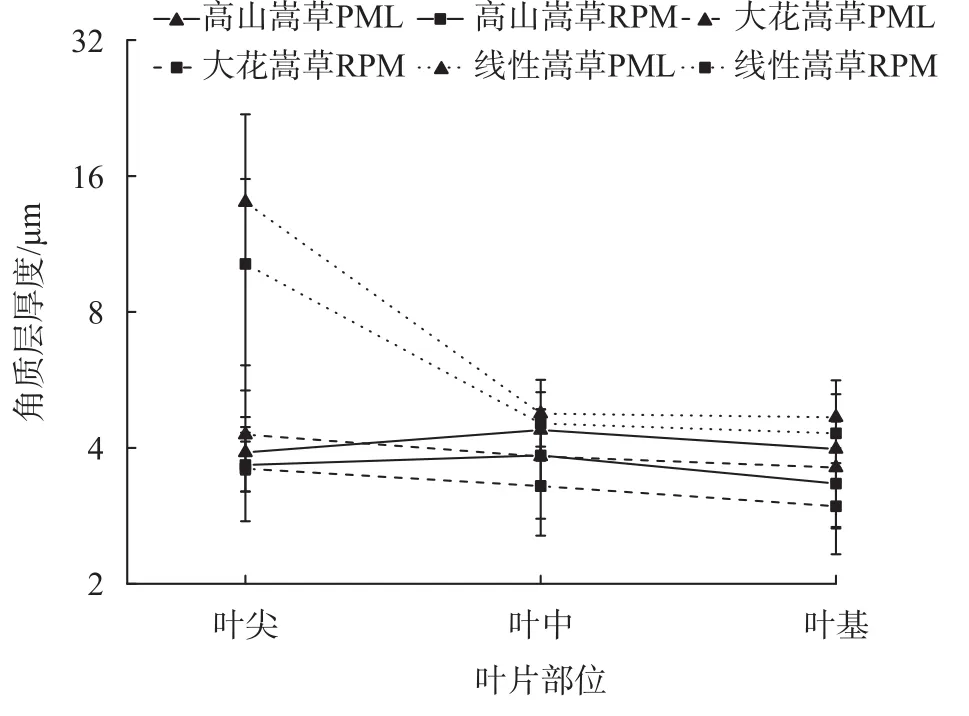
图 3 3种嵩草植物叶片不同部位角质层厚度的变化Fig. 3 Changes in the thickness of the stratum corneum in different parts of the leaves of 3 species of Kobresia
由图4可知,高山嵩草、线叶嵩草叶尖、叶中、叶基3部位的近、远轴面表皮细胞长度、宽度差异极显著(P < 0.01),而大花嵩草3部位近、远轴面表皮细胞仅长度差异显著(P < 0.05),宽度差异不显著。
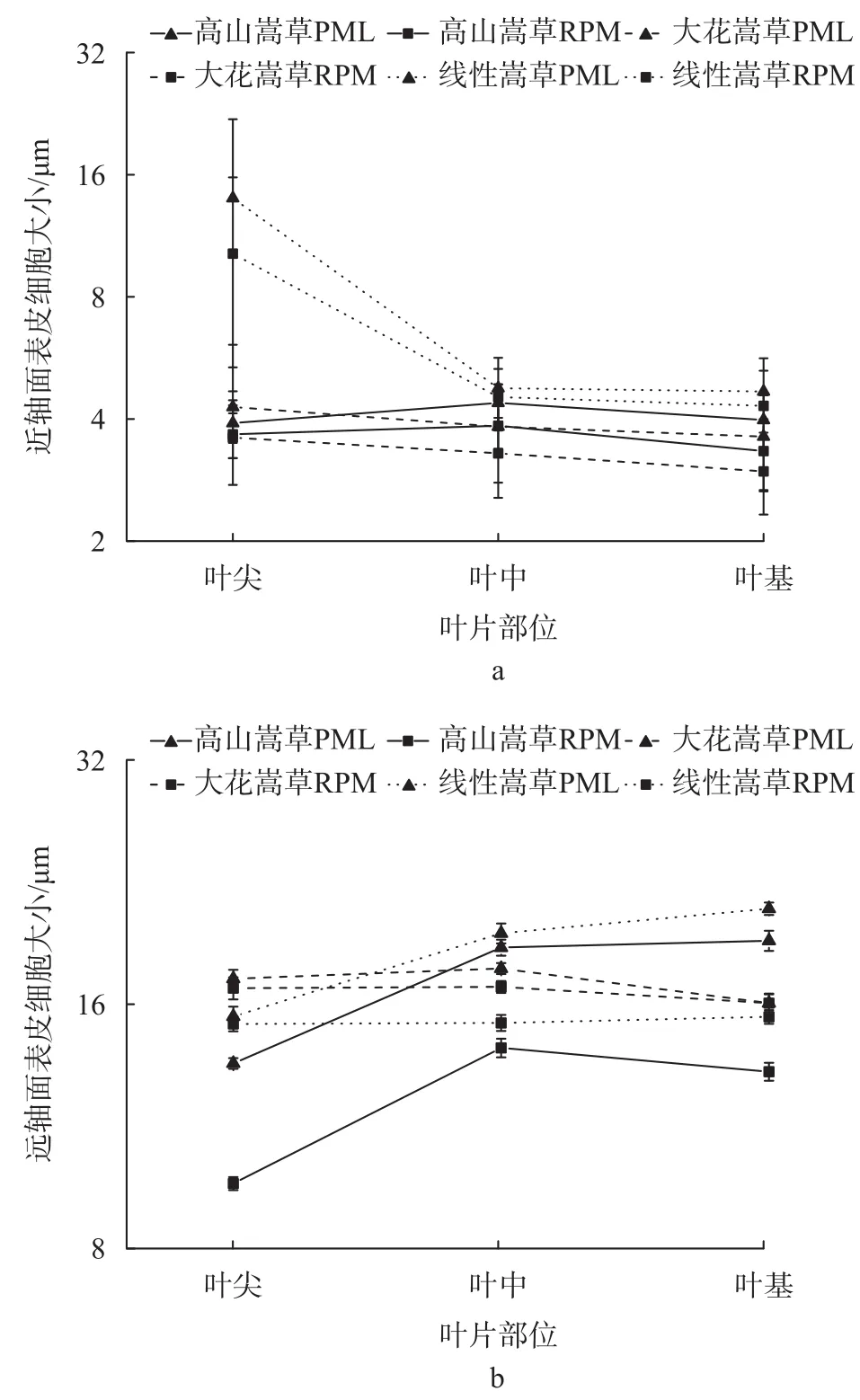
图 4 3种嵩草植物叶片不同部位表皮细胞的变化Fig. 4 Changes of epidermal cells in different parts of leaves of 3 species of Kobresia
由图5可知,高山嵩草和线叶嵩草叶尖、叶中、叶基的机械组织占叶片横切面面积的比例差异极显著(P < 0.01),而大花嵩草差异不显著。高山嵩草、大花嵩草叶尖、叶中、叶基机械组织占叶片横切面面积的比例变化一致,即叶尖>叶中>叶基,而线叶嵩草表现为叶中>叶尖>叶基。高山嵩草和线叶嵩草叶尖、叶中、叶基的维管束占叶片横切面面积的比例差异不显著,大花嵩草叶尖、叶中、叶基的维管束占叶片面积的比例差异极显著(P < 0.01),且叶尖部位显著高于叶中、叶基部位。高山嵩草和线叶嵩草叶尖、叶中、叶基维管束内鞘和外鞘面积差异均不显著,而大花嵩草3部位的内鞘面积和外鞘面积差异均极显著(P < 0.01)。
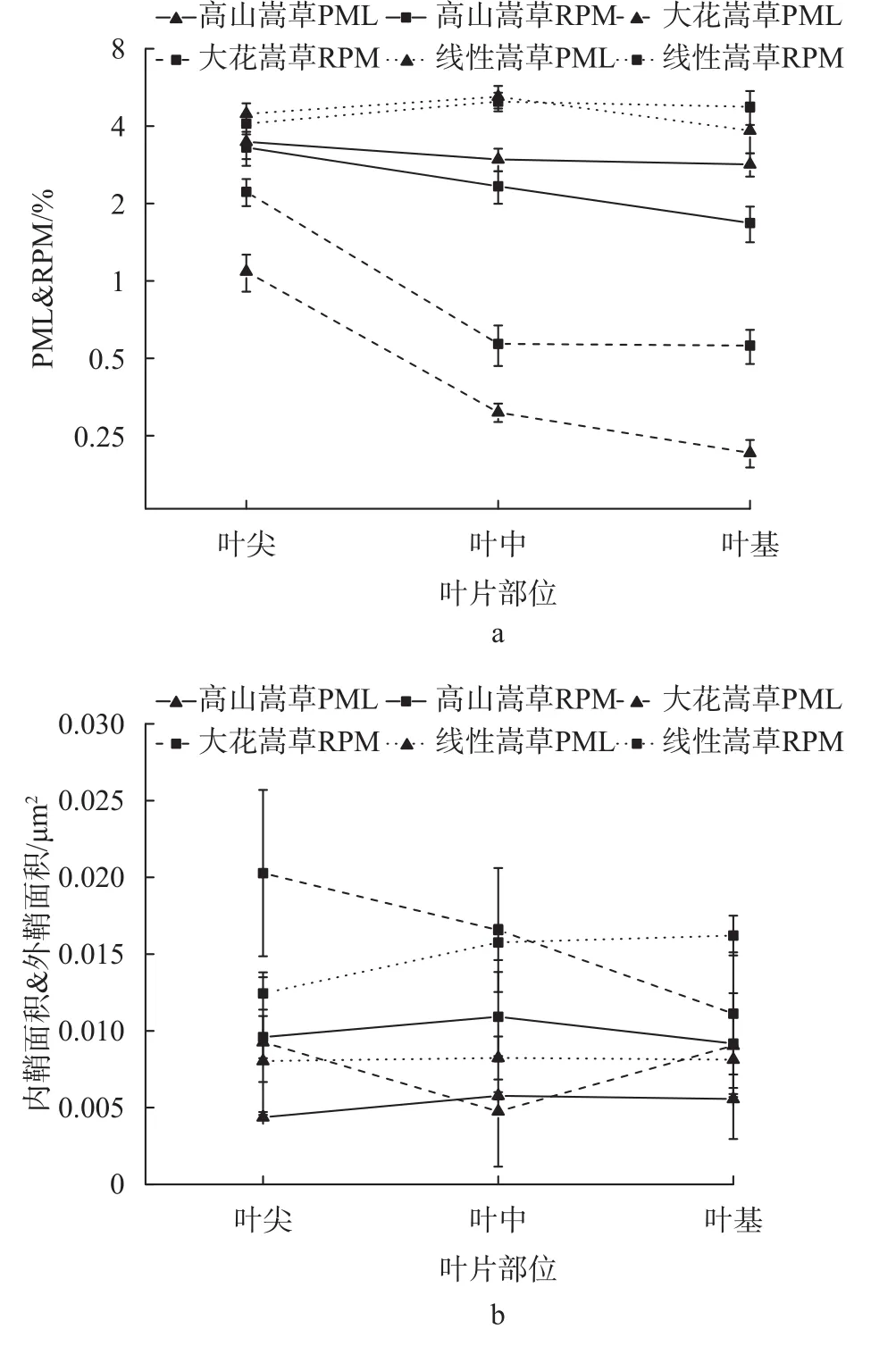
图 5 3种嵩草植物叶片不同部位维管束/叶片横切面面积、机械组织/叶片横切面面积、内鞘面积和外鞘面积变化Fig. 5 Variation of bundle/cross section of leaf area and mechanical tissue/cross section of leaf area between inter sheath area and outer sheath area in different parts of leaves for 3 species of Kobresia
4 结论与讨论
4.1 嵩草属植物叶片解剖结构的分类学意义
本研究发现,3种嵩草叶片解剖结构中近、远轴面表皮均由一层细胞组成;近轴面表皮细胞长度、宽度均大于远轴面。但是,3种嵩草属植物叶片解剖结构也存在显著差异。一是大花嵩草近轴面表皮细胞存在泡状细胞,而高山嵩草和线叶嵩草叶片表皮细胞上无泡状细胞。很多研究显示泡状细胞在种间分类研究上具有重要意义[19-20],因此研究结果可以为嵩草属植物分类提供重要依据。二是高山嵩草和大花嵩草近、远轴表皮细胞形状分别为长方形和近圆形,线叶嵩草近、远轴表皮细胞形状分别为长方形和椭圆形。这些解剖差异可以为种间分类依据[21]。
4.2 嵩草属植物叶的解剖结构与生境适应关系
3种嵩草叶片解剖结构不同程度地表现出旱生植物的典型解剖结构,说明不同植物叶片解剖结构对干旱生境的适应性不同。研究表明叶片厚度与抗旱性成正比,叶片越厚,抗旱性越强,这是因为叶片越厚,储水能力越强[22]。3种嵩草的叶片厚度均较厚,表现出显著的抗旱特性。将叶片厚度作为植物抗旱指标与柠条锦鸡(Caragana korshin-skii)、达乌里胡枝子(Lespedeza davurica)的研究结果一致[23-24]。其中线叶嵩草叶片厚度最厚,大花嵩草次之,高山嵩草最小。说明线叶嵩草具有更强的储水能力,能更好地应对干旱胁迫的影响。
3种嵩草植物均具有较厚的角质层,与虎榛子(Ostryopsis davidiana)、黄刺玫(Rosa xanthina)、白刺华(Sophora viciifolia)、沙棘(Hippophaerhamnoides)、文冠果(Xanthoceras sorbifolia)[25]、樱花(Prunus serrulata)、月季(Rosa chinese)、国槐(Sophora japonica)[26]等植物叶片在干旱生境中角质层较厚的研究结果一致。一般认为发达的角质层可以降低叶片蒸腾,防止体内水分散失,从而提高植物的抗旱性能[27-28]。其中,线叶嵩草角质层厚度最厚,高山嵩草次之,大花嵩草最小。关于表皮细胞对植物抗旱性的影响,有研究认为较厚的表皮细胞能增强植物的抗旱性[29-30],而其他研究认为表皮细胞越小,抗旱性越大[31-32]。本研究结果与后者一致,认为较小的表皮细胞有利于减少蒸腾,控制失水的能力越强。其中大花嵩草表皮细胞长度、宽度最小。除表皮细胞外,表皮细胞分化出的泡状细胞同样影响植物的抗旱性。一般认为发达的泡状细胞对失水具有灵敏的反应,可根据外部环境的变化随时调整叶片卷曲程度,减少水分的散失,克服水分供应不足,增强抗旱能力[33]。本研究结果显示3种嵩草植物中只有大花嵩草表皮细胞分化出泡状细胞,加之较小的表皮细胞,说明大花嵩草表皮细胞对生境干旱化的响应更为敏感。
通常情况下生长在干旱环境下的植物维管束和机械组织较为发达,因为发达的维管束能够增强植物运输水分的能力,发达的机械组织则可以减少植物萎蔫时所造成的损伤[34-35]。本研究中3种嵩草植物维管束和机械组织占叶片横切面面积比例较小,与黑褐苔草(Carex atrofusca)、华扁穗草(Blysmus sinocompressus)[36]、“阿坝”垂穗披碱草(Elymus natans)、“阿坝”硬秆仲彬草(Kengvilia rigidula)[37]等叶片具有发达的维管组织和机械组织的研究结果存在差异。其中,线叶嵩草维管束和机械组织占叶片横切面面积比例最大,大花嵩草显著低于高山嵩草和线叶嵩草,产生这种差异的原因很多,可能由于植物生育期短,生长缓慢,所以维管束和机械组织不发达,或者植物应对环境的策略不同。3种嵩草维管束都具有维管束鞘,这也是植物抗旱能力的体现[21]。与圆囊苔草(Carex orbicularis)[38]叶片通过维管束鞘增强抗旱能力的研究结果一致。线叶嵩草维管束鞘总面积最大,高山嵩草次之,大花嵩草最小。综上所述,3种嵩草植物相比较,线叶嵩草具有更高的储水能力、输送水分和营养的能力。说明线叶嵩草对干旱环境的适应能力最强。另外,3种嵩草植物适应干旱生境的能力与策略不同。线叶嵩草和高山嵩草主要利用较厚的角质层,降低水分蒸腾,从而对叶片进行保护,以及利用较厚的叶片提高其储水能力,限制叶片蒸腾,增强其抗旱性。而大花嵩草通过较小的表皮细胞减少蒸腾,尤其靠表皮细胞分化出的泡状细胞,来应对失水过多,即使水分供应不足,也能忍受干旱。
4.3 嵩草属植物叶片不同部位解剖结构对环境的响应
目前有关叶片不同部位的研究主要针对气孔密度和大小[39-40]、及生理差异[41]等方面,而关于叶片不同部位的解剖结构的研究还未见研究。本研究中3种嵩草叶片不同部位的解剖结构表现出一定的差异,其中,高山嵩草叶片不同部位中存在极显著差异的结构最多,在近、远轴面叶片厚度、角质层厚度、近、远轴面表皮细胞长度、宽度、机械组织占叶片横切面面积的比例共8项指标呈极显著差异。大花嵩草次之,在叶片厚度、近、远轴面角质层厚度、近轴面表皮细胞长度、维管束占叶片横切面面积的比例、内鞘面积和外鞘面积共7项指标呈极显著差异。而线叶嵩草只有5项指标呈极显著差异,即近、远轴面角质层厚度、远轴面表皮细胞长度、宽度、机械组织占叶片横切面面积的比例。说明同属不同种间叶片不同部位解剖结构存在差异及其对生境干旱化的响应不同。能否推测叶片不同部位解剖结构与生境敏感性之间存在关系,即叶片不同部位解剖结构差异越大,说明对生境干旱化越敏感,植物对环境的适应能力越强,这还需要今后进一步深入分析和探讨。
[ 参 考 文 献 ]
[1]李林, 王振宇, 徐维新, 等. 青藏高原典型高寒草甸生态系统对气候冻土环境变化的响应 [J]. 青海气象,2010(1): 15-22.
[2]李巧峡, 赵庆芳, 马世荣, 等. 嵩草属植物研究进展 [J]. 西北师范大学学报(自然科学版), 2006, 42(6):78-82.
[3]王勋陵, 马骥. 从旱生植物叶结构探讨其生态适应的多样性 [J]. 生态学报, 1999, 19(6): 787-792.
[4]张国云, 裴国亮, 姬燕. 利用扫描电镜研究嵩草属牧草种子形态和解剖学特征 [J]. 分析仪器, 2011(1):69-73.
[5]张国云, 何晓华, 郭付振. 扫描电镜在嵩草种子结构及发芽研究中的应用 [J]. 安徽农业科学, 2014,42(28): 9670-9673.
[6]李巧峡. 青藏高原东部嵩草属五种植物的遗传多样性研究 [D]. 兰州: 西北师范大学, 2005
[7]赵庆芳. 青藏高原东部嵩草属植物遗传多样性研究 [D]. 兰州: 兰州大学, 2006
[8]卢慧, 丛静, 刘晓, 等. 三江源区高寒草甸植物多样性的海拔分布格局 [J]. 草业学报, 2015, 24(7): 197-204.
[9]张大才, 李双智, 张锐. 高寒草甸湿润与干旱化生境中植物多样性的海拔分布格局 [J]. 草业科学, 2018,35(5): 1013-1021.
[10]卓玉璞. 不同利用方式对山地草甸线叶嵩草型草地植物群落特征的影响 [J]. 中国草食动物, 2011, 31(6):44-47.
[11]李英年, 赵亮, 徐世晓, 等. 祁连山海北高寒湿地植物群落结构及生态特征 [J]. 冰川冻土, 2006, 28(1):76-84.
[12]李芳兰, 包维楷. 植物叶片形态解剖结构对环境变化的响应与适应 [J]. 植物学通报, 2005, 40(S1): 118-127.
[13]李全发, 王宝娟, 安丽华, 等. 青藏高原草地植物叶解剖特征 [J]. 生态学报, 2013, 33(7): 2062-2070.
[14]张婧雯. 2种园林植物树冠不同部位叶片结构的变化 [J]. 黑龙江科学, 2017, 8(2): 82-83.
[15]朱玉怀, 张大才, 李双智. 西藏东达山3种嵩草属植物气孔特征沿海拔的变化 [J]. 西北植物学报, 2017,37(4): 728-736.
[16]张大才, 朱玉怀, 李双智. 东达山高寒草甸8种植物气孔特征沿土壤水分梯度的变化 [J]. 草业学报,2018, 27(7): 36-46.
[17]沈渭寿, 刘波, 颜长珍, 等. 青藏高原典型区生态状况时空变化及气候变化响应研究 [M]. 北京: 中国环境出版社, 2013.
[18]何艳艳. 石蜡制片的方法、技巧与注意事项 [J]. 农业研究与应用, 2014(4): 54-57.
[19]曲波, 朱明星, 陈旭辉, 等. 22种禾本科植物叶片泡状细胞形态特征的初步研究 [J]. 西北植物学报, 2010,30(8): 1595-1601.
[20]崔艳. 青藏高原东部嵩草属植物的生态解剖学研究 [D]. 兰州: 西北师范大学, 2006
[21]山宝琴, 贺学礼. 毛乌素沙地12种蒿属植物叶的解剖特征 [J]. 西北农林科技大学学报(自然科学版),2007, 35(6): 211-217.
[22]蒲文彩, 许云蕾, 余志祥, 等. 元江干热河谷典型耐旱植物叶片解剖结构特征及抗旱性分析 [J]. 西南林业大学学报(自然科学), 2019, 39(1): 58-68.
[23]韩刚, 李少雄, 徐鹏, 等. 6种灌木叶片解剖结构的抗旱性分析 [J]. 西北林学院学报, 2006, 21(4): 43-46.
[24]赵祥, 董宽虎, 张垚, 等. 不同居群达乌里胡枝子茎解剖结构与其抗旱性的关系 [J]. 草地学报, 2010, 18(5):615-621.
[25]薛智德, 韩蕊莲, 侯庆春, 等. 延安地区5种灌木叶旱性结构的解剖研究 [J]. 西北植物学报, 2004, 24(7):1200-1206.
[26]刘红茹, 冯永忠, 王得祥, 等. 延安城区10种阔叶园林植物叶片结构及其抗旱性评价 [J]. 西北植物学报,2012, 32(10): 2053-2060.
[27]朱栗琼, 徐艳霞, 招礼军, 等. 喀斯特地区莎叶兰的解剖构造及其环境适应性 [J]. 广西植物, 2016, 36(10):1179-1185.
[28]许云蕾, 蒲文彩, 余志祥, 等. 金沙江干热河谷同一群落不同生活型植物叶片解剖结构差异分析 [J]. 西南林业大学学报(自然科学), 2018, 38(6): 74-82.
[29]朱栗琼, 李吉跃, 招礼军. 六种阔叶树叶片解剖结构特征及其耐旱性比较 [J]. 广西植物, 2007, 27(3):431-434.
[30]徐扬, 陈小红, 赵安玖. 川西高原4种苹果属植物叶片解剖结构与其抗旱性分析 [J]. 西北植物学报,2015, 35(11): 2227-2234.
[31]王耀芝, 王勋陵, 李蔚. 荒漠化草原常见植物叶内部结构的观察 [J]. 兰州大学学报, 1983, 19(3): 87-96.
[32]孟庆杰, 王光全, 董绍锋, 等. 桃叶片组织结构与其抗旱性关系的研究 [J]. 西北林学院学报, 2005, 20(1):65-67.
[33]康萨如拉, 牛建明, 张庆, 等. 短花针茅叶片解剖结构及与气候因子的关系 [J]. 草业学报, 2013, 22(1):77-86.
[34]郭改改, 封斌, 麻保林, 等. 不同区域长柄扁桃叶片解剖结构及其抗旱性分析 [J]. 西北植物学报, 2013,33(4): 720-728.
[35]潘昕, 邱权, 李吉跃, 等. 基于叶片解剖结构对青藏高原25种灌木的抗旱性评价 [J]. 华南农业大学学报,2015, 36(2): 61-68.
[36]郭敏. 3种莎草科植物叶片形态解剖学及发育过程中细胞程序性死亡现象 [D]. 兰州: 甘肃农业大学, 2006
[37]肖冰雪, 杨满业, 陈琴, 等. 川西北高寒草地5种牧草叶片解剖结构与抗旱性关系研究 [J]. 草业与畜牧,2013(4): 1-5.
[38]强科斌, 郭敏, 张晓庆, 等. 圆囊苔草叶片的形态解剖学研究 [J]. 草业学报, 2007, 16(2): 76-83.
[39]Poole I, Weyers J D B, Lawson T, et al. Variations in stomatal density and index: implications for palaeoclimatic reconstructions [J]. Plant, Cell and Environment,1996, 19(6): 705-712.
[40]Salisbury E J. On the causes and ecological significance of stomatal frequency, with special reference to the wood land flora [J]. Philosophical Transactions of the Royal Society of London, 1927, 21(6): 1-65.
[41]潘泠, 樊瑞萍, 周琴, 等. 高羊茅越冬期叶片不同部位生理生化特性研究 [J]. 草业科学, 2010, 27(7): 5-9.
猜你喜欢
河北农业大学学报(2021年5期)2021-11-10
放射学实践(2021年9期)2021-09-22
建材发展导向(2021年14期)2021-08-23
新疆农业科学(2020年9期)2020-10-13
河南农业科学(2019年9期)2019-09-24
节水灌溉(2019年8期)2019-08-30
祝您健康·文摘版(2019年4期)2019-06-11
中国中西医结合皮肤性病学杂志(2016年4期)2016-07-18
中国房地产业(2016年9期)2016-03-01
儿童故事画报·发现号趣味百科(2015年7期)2015-10-23
