
外源一氧化氮供体硝普钠对番茄幼苗盐胁迫伤害的缓解作用
2019-08-29 07:26孙德智杨恒山张庆国苏雅乐其其格韩晓日
浙江农业学报 2019年8期
孙德智,杨恒山,张庆国,范 富,苏雅乐其其格,彭 靖,韩晓日
(1.内蒙古民族大学 农学院,内蒙古 通辽 028000; 2.沈阳农业大学 土地与环境学院,土肥资源高效利用国家工程实验室,辽宁 沈阳 110866)
盐害是限制作物生产潜力发挥的主要非生物逆境因子之一,是威胁生态安全、制约农业可持续发展的全球性问题。据报道,世界农业生产总量的30%以上来自于约占全球17%的灌溉可耕地和灌溉农业[1]。目前,全世界现有灌溉农业耕地面积的20%已经遭到了不同程度的盐渍化危害,受不合理灌溉、过度施肥和荒漠化进程等多因素的影响,次生盐渍化土地面积还将以年均1%~2%的增速继续扩大[1-2]。遏制盐渍土地蔓延、发展耐盐农业生产迫在眉睫。
番茄(SolanumlycopersicumL.)属茄科番茄属,为一年生或多年生草本植物,具有适应范围广、产量高、营养丰富、用途广泛、栽培方式多样以及栽培季节较长等特点[3-4]。番茄虽中度耐盐[4],但生长介质中盐分浓度过高,也会干扰株体正常的生理代谢,致使其光合生产潜力难以充分发挥,并最终导致产量降低,商品性变差,给生产造成严重的经济损失。
一氧化氮(nitric oxide,NO)是植物体内普遍存在的一种小分子气态活性信号物质[5-6]。它可广泛参与植物生长、发育及其相关生理代谢过程的调节,包括种子萌发、根叶的伸展、根系重力感应、光形态建成、呼吸作用、气孔关闭、细胞程序性死亡、组织的成熟和衰老以及各种逆境胁迫的防御反应等[6-8]。NO对植物的生理效应与其浓度密切相关[5,8-9]。李巧自等[10]研究了不同浓度SNP对铁皮石斛原球茎生长和多糖积累的影响,结果发现,低浓度SNP处理利于原球茎的生长,而高浓度SNP处理可促进多糖的积累。盐胁迫下,低浓度SNP可促进水稻种子的萌发和幼苗的生长,而SNP较高时则表现出抑制作用[11]。通过激活抗氧化酶活性和促进渗透调节物质的合成,适当浓度SNP处理的番茄[12]、黄瓜[13]幼苗叶片膜结构稳定性得到了有效保护,进而增强了植株盐渍逆境的适应性。杜卓涛等[14]在研究苦瓜幼苗耐受低温胁迫时也获得了相似的研究结果。目前,外源NO供体SNP调节植物适应非生物逆境胁迫的适宜浓度筛选研究多集中于对其植株生长、根叶抗氧化保护系统与渗透调节物质积累等方面的探讨,有关同步利用气体交换、叶绿素荧光技术和测定抗氧化生理指标系统分析不同浓度SNP影响植物光合生理特征的研究鲜见报道。本实验以秦丰保冠番茄幼苗为材料,通过NaCl模拟盐害环境,研究不同浓度的外源NO供体硝普钠(SNP)对NaCl胁迫下幼苗生长、叶片光合及抗氧化相关生理指标的影响,旨在探究SNP对番茄幼苗盐胁迫伤害的缓解效应,并筛选出适宜的SNP使用浓度,为进一步探明外源NO调节番茄幼苗耐盐的生理生态机制奠定基础。
1 材料与方法
1.1 材料培养与处理
供试材料为番茄品种秦丰保冠,由西安秦丰蔬菜研究所提供。采用营养钵育苗,选取均一、饱满的种子浸种催芽(55 ℃温水中浸泡3~4 h后放在铺有湿润纱布的培养皿内,置于29 ℃恒温箱中),露白后将芽势相近的种子播于直径10 cm、高10 cm的塑料营养钵中,每钵2粒,以蛭石作基质,当幼苗破心后每钵保留1株。在日光温室内培育,待第一片真叶展平后每2 d浇1/8浓度Hoagland营养液1次,每钵浇50 mL,当幼苗长至4~5片真叶时,挑选长势较好且一致的植株洗净根部育苗基质后,定植于长60 cm、宽40 cm、高20 cm的水培箱中,每箱6株,株行距均为15 cm,用1/4浓度Hoagland营养液栽培,利用充气泵24 h不间断补充营养液中氧气,每2 d更换1次营养液。
定植后恢复生长10 d,开始进行试验。共设7个处理:1)对照(CK),1/4 Hoagland营养液;2)处理1(T1),1/4 Hoagland营养液+100 mmol·L-1NaCl;3)处理2(T2),1/4 Hoagland营养液+100 mmol·L-1NaCl+50 μmol·L-1SNP;4)处理3(T3),1/4 Hoagland营养液+100 mmol·L-1NaCl+100 μmol·L-1SNP;5)处理4(T4),1/4 Hoagland营养液+100 mmol·L-1NaCl+200 μmol·L-1SNP;6)处理5(T5),1/4 Hoagland营养液+100 mmol·L-1NaCl+400 μmol·L-1SNP;7)处理6(T6),1/4 Hoagland营养液+100 mmol·L-1NaCl+800 μmol·L-1SNP。每处理3次重复,水培箱随机排列,为保证处理浓度的稳定性,处理期间每天更换1次处理液。处理第6天,取幼苗上数第2片完全展开叶进行各项生理指标的测定,第8天结束处理后进行生长量和干物质积累的测定。
1.2 测定项目与方法
1.2.1 生长指标的测定
用直尺测量幼苗株高(茎基部到生长点高度);用游标卡尺测量茎粗(子叶下部2/3处);将植株分离根、茎、叶,分别洗净(自来水冲洗3次,蒸馏水冲洗2次,用吸水纸吸干)后立刻称鲜质量,之后置于105 ℃鼓风烘箱中杀青15 min,再于75 ℃下烘干,称干质量。计算壮苗指数,壮苗指数=(茎粗/株高+根干质量/茎叶干质量)×全株干质量。各指标测定均为3次重复,每个重复随机取样调查6株,取平均值。
1.2.2 光合色素含量的测定
称取0.2 g剪碎、混匀的叶片(去除中脉)放入研钵中,加少许石英砂、CaCO3粉及2~3 mL 80%的丙酮研磨成匀浆后定容至25 mL棕色容量瓶中,摇匀,暗处保存24 h,备用待测。用紫外-可见分光光度计(UV-1750,Shimadzu,Japan)在25℃下分别测定663、645、470 nm波长下的光密度(D值),然后按Lichtenthaler等[15]方法分别计算叶绿素a(Chla)、叶绿素b(Chlb)、类胡萝卜素(Car)和叶绿素a+b(Chla+b)值。
1.2.3 气体交换参数的测定
利用LI-6400XT便携式光合仪(LI-COR,Lincoln,NE,USA)测定叶片的净光合速率(Pn)、气孔导度(Gs)和胞间CO2浓度(Ci),每处理重复测定6株。测定时使用开放式气路、内置LED红蓝光源(6400-02B),设定空气流速为500 μmol·s-1,控制光强为800 μmol·m-2·s-1、叶室温度为(28±2)℃、CO2浓度为(360±20)μL·L-1。
1.2.4 叶绿素荧光参数的测定
采用JUNIOR-PAM便携式脉冲调制叶绿素荧光仪(Heinz Walz GmbH,Effeltrich,Germany)测定叶片的叶绿素荧光参数,每处理重复测定6株。将叶片暗适应30 min后,首先开启检测光,测得初始荧光(Fo),再由饱和脉冲光激发,得到最大荧光(Fm)。随后打开内源光化光,10 min后获得光下的稳态荧光(Fs),并再次照射饱和脉冲光以获得光下最大荧光(Fm′)。关闭光化光的同时快速遮光测定叶片,并开启远红光测定光下最小荧光(Fo′)。PSⅡ最大光化学效率(Fv/Fm)、光化学猝灭系数(qP)、非光化学猝灭系数(qN)和实际光化学效率(ΦPSⅡ)均由仪器自动给出。
1.2.5 相对电导率、丙二醛(MDA)含量和抗氧化酶活性的测定
参照张宪政[16]的方法测定电解质渗出率;采用硫代巴比妥酸(TBA)法[17]测定丙二醛(MDA)含量。超氧化物歧化酶(SOD)活性测定采用氮蓝四唑(NBT)还原法[17];过氧化物酶(POD)活性测定采用愈创木酚比色法[18];过氧化氢酶(CAT)活性采用紫外吸收法[17]测定;抗氧化酶活性测定时采用同一提取液体系,提取液为pH 7.8(50 mmol·L-1)磷酸缓冲液。各项指标的测定均重复3次。
1.3 数据分析与作图
用Excel 2003和Origin Pro 8.5软件整理实验数据和作图,用 SPSS 19.0统计软件进行单因素方差分析,采用Duncan新复极差法进行差异显著性检验(P<0.05)。
2 结果与分析
2.1 外源NO供体SNP对NaCl胁迫下番茄幼苗生长的影响
由表1可见,与CK相比,NaCl胁迫处理(T1)后的番茄幼苗生长受到显著抑制,其株高、茎粗、生物量和壮苗指数分别下降了25.16%、22.37%、38.31%和26.74%。与T1相比,施加不同浓度SNP处理(T2~T6)的番茄幼苗株高、茎粗、生物量和壮苗指数均有不同程度增加,且随SNP处理浓度的增加呈先上升后下降的变化趋势,在T3处理时增幅达到最大,其株高、茎粗、生物量和壮苗指数分别比T1处理提高了23.79%、13.38%、44.21%和25.40%,但以上指标与CK相比仍显著偏低。由此可见,外源NO处理可有效缓解NaCl胁迫对番茄幼苗生长的抑制作用,且具有剂量效应,其中以100 μmol·L-1SNP处理效果最好,但仍未能恢复至CK的水平。
2.2 外源NO供体SNP对NaCl胁迫下番茄幼苗叶片光合色素含量的影响
番茄幼苗叶片叶绿素a(Chla)、叶绿素b(Chlb)、叶绿素a+b(Chla+b)总量和类胡萝卜素(Car)含量在各处理下表现出相似的变化趋势(图1)。其中,NaCl胁迫处理(T1)幼苗叶片的以上各光合色素含量分别较CK显著降低了37.39%、28.78%、35.37%和19.81%(P<0.05)。各浓度SNP处理(T2~T6)均能不同程度提高NaCl胁迫下叶片各光合色素的含量,且随SNP浓度增加,均呈现先升高后降低的变化趋势。在T3处理时各指标增幅均达到最大,Chla、Chlb、Chla+b和Car含量依次分别比T1显著提高了35.92%、26.19%、33.45%和14.15%(P<0.05)。以上结果说明,外源NO可能通过提高叶片光合色素含量来促进幼苗的光合作用,从而增强其对盐胁迫的抗性,并以100 μmol·L-1SNP处理效果最好。
表1 硝普钠对NaCl胁迫下番茄幼苗生长的影响
Table 1 Effect of sodium nitroprusside on the growth of tomato seedlings under NaCl stress
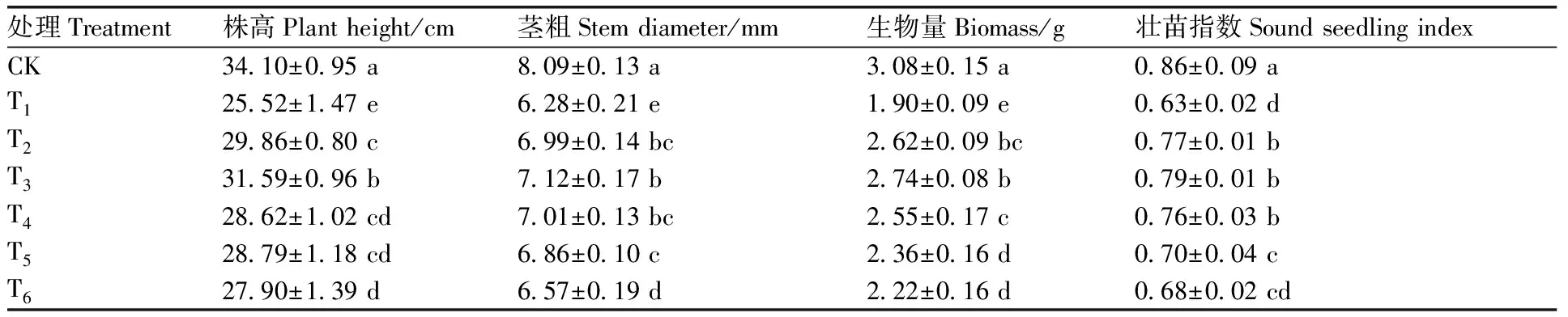
处理Treatment株高Plant height/cm茎粗Stem diameter/mm生物量Biomass/g壮苗指数Sound seedling indexCK34.10±0.95 a8.09±0.13 a3.08±0.15 a0.86±0.09 aT125.52±1.47 e6.28±0.21 e1.90±0.09 e0.63±0.02 dT229.86±0.80 c6.99±0.14 bc2.62±0.09 bc0.77±0.01 bT331.59±0.96 b7.12±0.17 b2.74±0.08 b0.79±0.01 bT428.62±1.02 cd7.01±0.13 bc2.55±0.17 c0.76±0.03 bT528.79±1.18 cd6.86±0.10 c2.36±0.16 d0.70±0.04 cT627.90±1.39 d6.57±0.19 d2.22±0.16 d0.68±0.02 cd
同列数据后没有相同字母表示在0.05水平存在显著性差异。
Different letters in the same column indicated significant differences at the 0.05 level.

图中不同小写字母表示不同处理间的差异显著(PDifferent letters indicated the significant differences among various treatments at P图1 硝普钠对NaCl胁迫下番茄幼苗叶片光合色素含量的影响Fig.1 Effect of sodium nitroprusside on photosynthetic pigment content in leaves of tomato seedlings under NaCl stress
2.3 外源NO供体SNP对NaCl胁迫下番茄幼苗叶片光合作用气体交换参数的影响
由图2可知,与CK相比,NaCl胁迫处理(T1)下的叶片净光合速率(Pn)和气孔导度(Gs)分别显著降低了60.63%和39.56%,而胞间CO2浓度(Ci)却显著升高了18.97%(P<0.05)。与T1相比,施加不同浓度SNP处理(T2~T6)的番茄幼苗叶片Pn和Gs均显著升高,Ci则不同程度降低,且呈现出随SNP浓度增加,Pn和Gs先升高后降低,Ci与上述2个参数表现出完全相反的趋势变化。在T3处理时变幅达到最大,其Pn和Gs分别较T1处理显著提高了85.91%和39.69%,Ci则比T1显著降低了9.38%(P<0.05)。以上结果说明,NaCl胁迫显著影响了番茄幼苗的光合作用效率,外源NO可以有效缓解盐胁迫的伤害,并以100 μmol·L-1SNP处理效果最好。

图2 硝普钠对NaCl胁迫下番茄幼苗叶片气体交换参数的影响Fig.2 Effects of sodium nitroprusside on gas exchange parameters in leaves of tomato seedlings under NaCl stress
2.4 外源NO供体SNP对NaCl胁迫下番茄幼苗叶片叶绿素荧光参数的影响
由图3可知,与CK相比,NaCl胁迫处理(T1)的幼苗叶片初始荧光(Fo)与非光化学淬灭系数(qN)分别显著升高了38.40%和27.45%,最大荧光(Fm)、PSⅡ最大光化学效率(Fv/Fm)、实际光化学效率(ΦPSⅡ)和光化学淬灭系数(qP)则分别显著降低了22.83%、18.04%、72.15%和54.44%(P<0.05)。经SNP处理后,叶片Fo与qN均表现为先降低后升高的变化趋势,而Fm、Fv/Fm、ΦPSⅡ和qP表现为先升高后下降的变化趋势。与T1相比,各浓度SNP处理(T2~T6)的各指标变化均达显著水平,除Fm外,各参数均在T3处理时变幅达到最大,其Fo和qN分别比T1处理降低了16.53%和13.85%,Fv/Fm、ΦPSⅡ和qP则分别比T1处理提高了13.02%、125.98%和61.72%。Fm虽在T4处理时变幅达到最大,但其值与T3处理相比差异并不显著,与T1相比显著提高了13.98%(P<0.05)。以上结果说明,外源NO处理提高了番茄幼苗叶片的PSⅡ光化学活性,减轻了光抑制对类囊体膜的破坏,并有效地降低了天线色素的非光化学热耗散。
2.5 外源NO供体SNP对NaCl胁迫下番茄幼苗叶片抗氧化酶活性的影响
在NaCl胁迫条件下,不同浓度SNP处理(T2~T6)对番茄幼苗叶片SOD、POD和CAT的活性均具有显著影响,且随SNP处理浓度的增加,3种酶活性均呈先升高后下降的趋势变化(图4)。在单独NaCl胁迫处理(T1)时,叶片CAT活性与CK相比变化不显著(P>0.05),SOD与POD活性显著升高(P<0.05),2指标分别比CK高出了36.74%和23.83%。与T1相比,T2~T6处理均可显著提高叶片SOD和CAT的活性,对POD活性的影响较为复杂,T2~T4处理时,POD活性显著升高;T5和T6处理时,POD活性显著降低(P<0.05)。在T3处理时,SOD、POD和CAT活性均获得最大增幅,3指标分别比T1处理依次提高了62.92%、47.80%和58.28%。可见,外源NO对NaCl胁迫下番茄幼苗叶片抗氧化保护酶活性的影响亦存在剂量效应,各处理条件下以100 μmol·L-1SNP处理效果最佳,浓度过高不利于促进抗氧化酶活性的升高。

图3 硝普钠对NaCl胁迫下番茄幼苗叶片叶绿素荧光参数的影响Fig.3 Effects of sodium nitroprusside on chlorophyll fluorescence parameters in leaves of tomato seedlings under NaCl stress
2.6 外源NO供体SNP对NaCl胁迫下番茄幼苗叶片膜脂过氧化和膜透性的影响
番茄幼苗叶片MDA含量和电解质渗出率在各处理下表现出相似的变化趋势(图5)。其中,NaCl胁迫处理(T1)幼苗叶片的MDA含量和电解质渗出率分别较CK显著高出了158.15%和137.10%(P<0.05)。各浓度SNP处理(T2~T6)均能显著降低NaCl胁迫下叶片的MDA含量和电解质渗出率,且随SNP浓度增加,两参数均呈现先降低后升高的变化特征。在T3处理时降幅达到最大,MDA含量和电解质渗出率分别比T1显著降低了35.31%和34.65%(P<0.05)。可见,NaCl胁迫引发了膜脂过氧化,导致膜结构受损,外源NO能显著缓解这种伤害程度,并以100 μmol·L-1SNP处理效果最佳,浓度过高反而会削弱缓解效果。
3 讨论
盐胁迫通过扰乱生理代谢阻碍植物正常的生长发育[19],继而导致植株生物积累量下降,难以形成壮苗。SNP是一种较为常用的外源NO供体,Delledonne等[20]证明了0.5 mmol·L-1的SNP能够产生约2 μmol·L-1的NO。非逆境条件下,恰当施用一定浓度SNP处理有利促进番茄种子萌发和幼苗的生长发育[21]。盐渍逆境下,外源施用50~800 μmol·L-1SNP处理起到了缓解番茄幼苗生长抑制的作用(使植株株高、茎粗、生物量和壮苗指数均不同程度升高),这与吴雪霞等[12]的研究结果一致。说明适当浓度外源NO供体SNP处理可以调节植物生理代谢活动向着有利于促进植物生长发育的方向变化。

图4 硝普钠对NaCl胁迫下番茄幼苗叶片抗氧化酶活性的影响Fig.4 Effects of sodium nitroprusside on antioxidase activity in leaves of tomato seedlings under NaCl stress

图5 硝普钠对NaCl胁迫下番茄幼苗叶片丙二醛含量和电解质渗出率的影响Fig.5 Effects of sodium nitroprusside on malondialdehyde content and electrolyte leakage in leaves of tomato seedlings under NaCl stress
光合作用是植物生长发育,形成生物产量的前提。盐胁迫下,不同浓度SNP处理对番茄幼苗生长抑制的缓解最终归因于叶片光合功能的改善。光合色素作为植物吸收、传递和转换光能,进行光合作用的物质基础,其含量的高低直接关系到叶片光合功能的充分发挥。许多研究证实,环境因子的改变可以引起光合色素含量的变化,进而引发光合功能的改变[22]。本研究结果表明,盐胁迫下番茄幼苗叶片Chla、Chlb、Chl(a+b)和Car含量均显著降低,而施加SNP处理可使胁迫下幼苗叶片的各色素含量获得不同程度的提高,说明外源NO可通过提高盐胁迫下光合色素的含量而影响番茄幼苗的生长发育,进而增强了植株的耐盐性。
盐胁迫下,植物光合作用受阻是多种因素共同作用的结果,除光合色素含量降低外,还包括因渗透胁迫引发气孔部分关闭而产生的气孔限制,以及由叶肉细胞光合活性降低而导致的非气孔限制。根据Farquhar等[23]提出气体交换模型理论判断,如果胁迫使气孔导度(Gs)减小,而叶肉细胞仍能活跃地进行光合作用时,细胞间隙CO2浓度(Ci)应有明显的降低,这种情况是典型的气孔限制;如果叶肉细胞本身光合能力显著降低,即使在Gs较低的情况下,Ci也可能升高或保持不变,此时非气孔限制就成了光合速率(Pn)降低的主导因素。本实验中,NaCl胁迫处理的第6天,番茄幼苗叶片Pn和Gs均显著降低、而Ci却显著增加,这表明Pn降低的主导因素是非气孔限制。与单独盐胁迫处理相比,50~800 μmol·L-1的SNP处理不同程度缓解了NaCl胁迫下叶片Pn、Gs的降低和Ci的升高,说明适当浓度外源NO处理可以有效抑制NaCl胁迫对番茄幼苗光合作用产生的非气孔限制,进而增强了叶肉细胞对CO2同化利用,使植株叶片在盐胁迫下依然可以维持较高的光合能力,相似的研究结果也出现在常青山等[5]对夏枯草幼苗的报道中。
盐逆境下,细胞叶绿体和线粒体电子传递中泄露的电子积累会诱发活性氧自由基(ROS)的大量产生,过量的ROS将直接攻击生物膜,引起或加剧了膜脂过氧化作用[28]。丙二醛(MDA)作为膜脂过氧化产物,能与蛋白质结合造成膜蛋白变性,从而直接危害膜结构的稳定性与完整性,致使细胞膜透性增大,进一步导致电解质的大量外渗[29]。为防御ROS的氧化伤害,植物在长期进化过程中形成了一整套行之有效的ROS清除系统。其中,超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)作为这一系统内的3种主要抗氧化酶,在保护植物体免受ROS氧化损伤和防御膜结构破坏方面起着至关重要的作用[5,8,12-14]。本实验结果显示,NaCl胁迫处理的第6天,施加不同浓度SNP处理的番茄幼苗叶片MDA含量和电解质渗出率显著降低;SOD和CAT活性显著升高;POD活性在50~200 μmol·L-1SNP处理下显著升高,在400、800 μmol·L-1SNP处理下显著降低,并均以100 μmol·L-1SNP处理时效果最好,这与吴雪霞等[12]的研究结果一致,表明适宜浓度的外源NO可通过提高抗氧化酶活性,增强叶片ROS的清除能力,使细胞膜脂过氧化程度降低、膜结构稳定性增强,进而维持了番茄幼苗较高的盐渍耐受性。
猜你喜欢
舰船科学技术(2022年11期)2022-07-15
全科护理(2022年14期)2022-05-16
中国医药导报(2021年35期)2022-01-20
中国土壤与肥料(2021年5期)2021-12-02
载人航天(2021年5期)2021-11-20
农业资源与环境学报(2021年5期)2021-10-06
实用器官移植电子杂志(2020年4期)2020-08-31
吉林农业科技学院学报(2020年2期)2020-05-15
电子制作(2017年20期)2017-04-26
农家科技中旬版(2016年7期)2016-10-20
