
水稻缺铁应答机制研究进展
2019-08-20 14:58邓凌韦王利军王永力李琬马军韬张丽艳邹世辉王英
江苏农业科学 2019年8期
关键词:烟酰胺
邓凌韦 王利军 王永力 李琬 马军韬 张丽艳 邹世辉 王英
摘要:铁是植物生长发育必需的微量元素,也是人体不可或缺的微量元素之一。水稻作为世界上50%人口的主食,可有效地从土壤中吸收铁供人类食用,有助于改善人类铁缺乏的现状。本文综述了水稻铁吸收及转运的调控机制,并对铁在水稻根、茎部运输的最新研究进展进行了阐述,为水稻铁生物强化提供参考,为水稻富铁品种的培育提供思路。
关键词:铁生物强化;铁缺乏;脱氧麦根酸;烟酰胺
中图分类号: S511文献标志码: A
文章编号:1002-1302(2019)08-0012-06
人体缺铁是世界上最严重的营养缺乏症之一,危害严重。人体主要从食物中摄入铁。水稻是重要的粮食作物,全世界50%以上的人口以水稻为主食。发展中国家人们主要从稻米中摄入铁元素,例如在菲律宾(包括高收入家庭),人体50%的铁摄入来自于水稻、玉米[1]。水稻生长所需可溶性铁浓度在10-9~10-4 mol/L之间[2],但即使是通气良好、pH值适宜的土壤中,可溶性铁浓度不超过10-15 mol/L,碱性土壤中可溶性铁浓度只有10-17 mol/L[3]。土壤可溶性铁离子不足降低了水稻铁含量,因此水稻铁生物强化育种成为农作物国际挑战计划HarvestPlus项目中的一个重要研究内容,2008年哥本哈根会议(http://www.copenhagenconsensus.com/)更是明确指出,生物强化是解决生物体缺铁这一世界营养缺乏症最经济、有效的途径[4]。
虽然近年来水稻铁生物强化取得了很多进展,但水稻传统育种周期长、工作量大,铁元素在稻米中含量又低,迫切须要研究出一种简便、快捷、经济的铁含量检测方法供育种者使用,并根据缺铁应答机制制定育种方案,调节相关基因,培育富铁品系,加速育种进程。本文综述水稻缺铁应答机制研究进展,并对未来的研究工作进行展望,旨在为水稻铁生物强化育种提供参考。
1水稻缺铁应答机制的特殊性
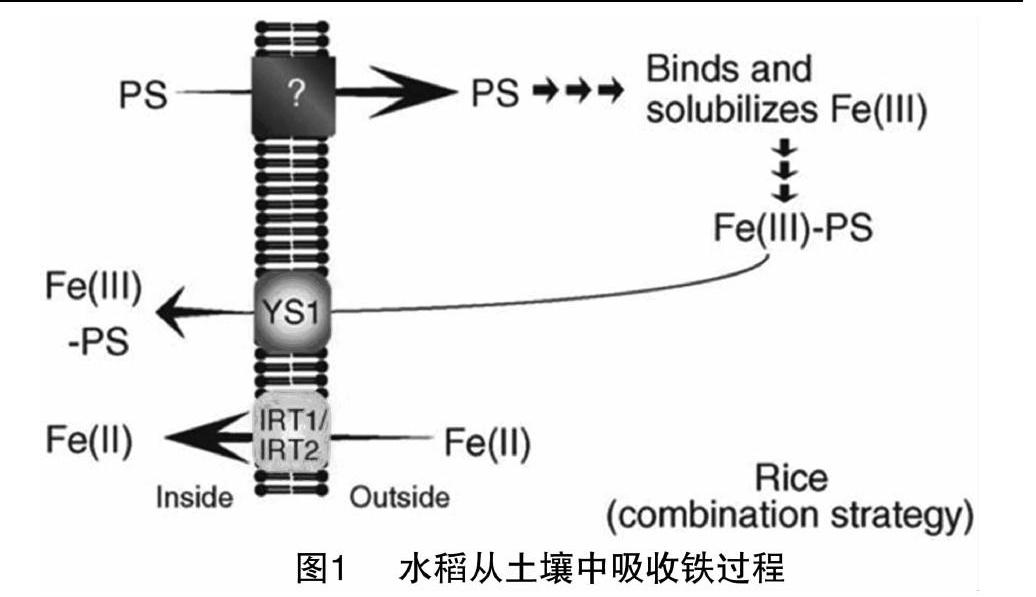
1986年,R-mheld等根据应答方式不同,将植物缺铁应答机制分为机制Ⅰ(还原机制)和机制Ⅱ(螯合机制)2种方式[5]。机制Ⅰ植物(双子叶植物和非禾本科单子叶植物)先将Fe3+还原成Fe2+再吸收利用;机制Ⅱ植物(仅限于禾本科植物)吸收Fe3+的鳌合物,不能还原Fe3+。ys1突变体不能吸收Fe3+-PS复合体(Fe3+-植物铁载体复合体),导致苗期叶片失綠,最终死亡[6],说明Fe3+-PS复合体吸收是机制Ⅱ植物缺铁应答的必要步骤[6-7]。
水稻是特殊的机制Ⅱ植物。OsIRT1、OsIRT2基因被定位在根表皮细胞质膜上,具有类似机制Ⅰ植物Fe2+运载体IRT1功能[8]。naat1突变体不能合成铁载体(PS),该突变体在Fe3+源条件下生长缓慢,叶片失绿;但在Fe2+源条件下正常生长[9]。这些试验结果说明,水稻虽然不能还原Fe3+,但能吸收Fe2+[10-11],所以有些学者直接把水稻定义为混合机制植物(图1)。
2缺铁感应
2.1缺铁感应元件
为了避免自由基的毒害作用,水稻进化出精密的铁吸收调控系统[12]。水稻缺铁感应元件包括IDEF1(iron deficiency-responsive element-binding factor 1)[13]、HRZ[hemerythrin motif-containing really interesting new gene (RING)- and zinc-finger proteins][14]。水稻HRZ是拟南芥BTS的同源蛋白质[15]。
2.1.1IDEF1
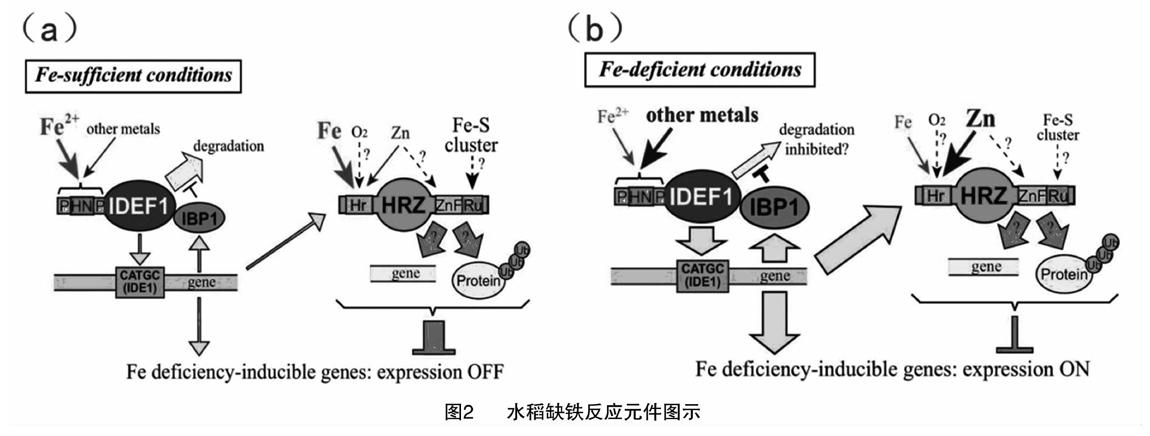
与其他缺铁应答相关基因不同,IDEF1转录本不受铁有效浓度影响,因而推测IDEF1位于缺铁应答级联反应的上游,是缺铁感应元件[16-17]。IDEF1通过结合基因序列普遍存在的CATGC序列[18]精密调控缺铁应答顺式元件基因[19]。IDEF1的组氨酸-天冬酰胺重复序列(histidine-asparagine repeat)、脯氨酸富集区域(proline-rich regions)(图2)是二价金属离子结合域,根据细胞内铁离子浓度在总金属离子浓度中的比例调节水稻缺铁应答反应[13]。
在铁充足条件下,26S蛋白酶系统降解IDEF1,使之维持本底表达[20]。在缺铁条件下,IBP1(bowman-birk trypsin inhibitor designated IDEF1-binding protein 1)抑制26S蛋白酶系统活性,水稻体内IDEF1浓度增加,当IDEF1积累到阈值浓度时,启动缺铁应答基因[21]。
2.1.2HRZs
HRZs是水稻缺铁诱导基因[14,22],[JP3]该基因所编码的蛋白质含有保守的hemerythrin、CHY-、CTCHY-、RING-Zn-fingers、rubredoxin-type fold[14]。Hemerythrin结合二价金属,RING-Zn-fingers是26S蛋白酶系统泛素化位点。即使在铁充足条件下,HRZs敲除植株体内缺铁应答基因都表达,说明HRZs通过负向调节缺铁应答系统基因,维持植株体内铁动态平衡[14]。
2.2调节缺铁应答的信号分子
IDEF1根据细胞内铁离子浓度在总金属离子浓度中的比例调节水稻缺铁应答反应[13],HRZs的Hemerythrin结构域能够结合2个铁分子,其中第2个位点能结合氧分子,且这个过程可逆。Kobayashi等的试验结果说明,铁分子和氧分子都是调节缺铁应答的信号分子[23]。为了避免自由基毒害,水稻体内的铁大多以螯合或铁蛋白质形式存在。HRZs蛋白质的rubredoxin-type fold结构域含有Fe-S簇,烟酰胺(nicotianamine)和麦根酸(deoxymugineic acid)是铁螯合剂,可影响铁在水稻体内的移动性和可利用性,因而推测Fe-S、烟酰胺、麦根酸是缺铁信号[23]因子。
缺铁诱导机制Ⅰ植物合成乙烯,但不能诱导机制Ⅱ植物合成乙烯[3]。水稻是混合机制植物(既可以吸收Fe2+,也可以吸收Fe3+)。缺铁诱导水稻合成乙烯,乙烯合成的1-氨基环丙烷1-羧酸(1-Aminocyclopropane-1-Carboxylic,ACC)能够增加水稻对缺铁的耐受性;乙烯抑制剂C2+和STS抑制缺铁应答基因的表达。进一步研究发现,乙烯调节OsIRO2的表达,OsIRO2调节OsNAS1/2和OsYSL15的表达[24]。目前还不清楚缺铁诱导植物合成乙烯的分子机制[3]。
[JP3]生长素和一氧化氮能够正向调节缺铁应答反应[25]。水稻中的生长素响应转录因子OsARF12(auxin response factor,ARF)突变后,根部构型发生改变,突变体对生长素不敏感,铁含量低于野生型[26]。细胞分裂素和茉莉酸负向调控水稻缺铁应答[3]。
3铁在根部的吸收及运输
3.1铁的吸收机制——还原机制
3.1.1Fe3+的还原
机制Ⅰ植物的三价铁还原酶FRO(ferric-chelate reductase)存在于根部,负责还原根际Fe3+[27]。虽然水稻有2个FRO基因,但在根部检测不到,只有在叶片能检测到这2个基因。OsFRO1基因受锌、锰、铜信号调节,OsFRO2基因参与缺铁应答。这些试验结果说明,OsFROs基因并不参与根际Fe3+的还原过程[8]。因此,有学者认为,水稻缺铁应答不需要还原Fe3+。
3.1.2Fe2+的吸收
机制Ⅰ植物二价铁转运体(iron-regulated sporter,IRT)负责Fe2+由根际到根内细胞的运输。目前水稻中只克隆到2个OsIRT基因,这2个基因都定位在质膜上,转运铁、锌离子。OsIRT1在根表皮、伸长区外皮层和成熟区皮层的内层、伴胞中表达[8,28]。组成性Fe、Mn、Cd运载体OsNRAMP5可能参与铁吸收[29-30]。Fe、Cd运载体OsNRAMP1参与根到茎部铁的运输[31]。
3.2铁的吸收机制——螯合机制
3.2.1铁载体的合成
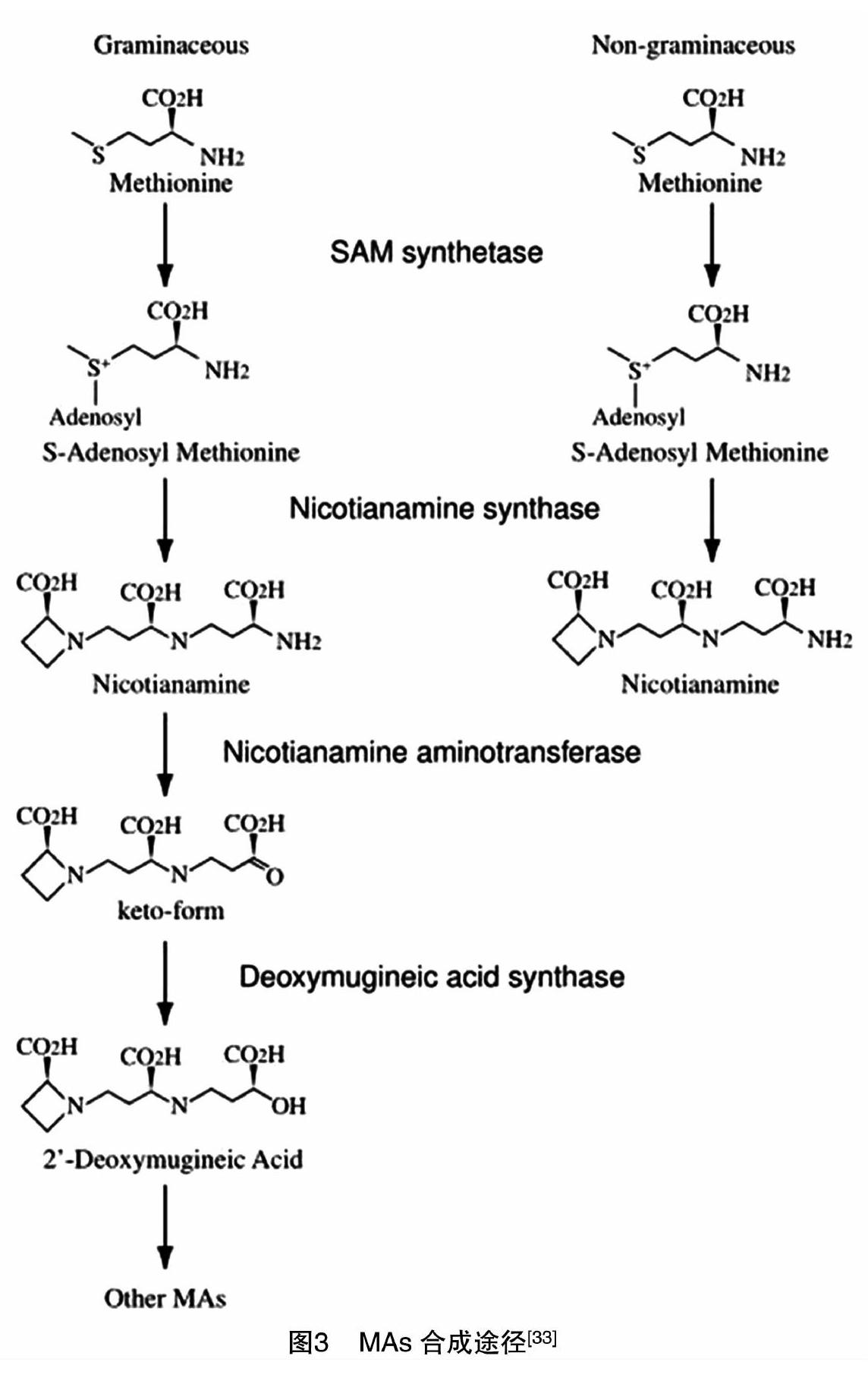
水稻根系所分泌的麦根酸(mugineic acid,MA)能够强烈鳌合Fe3+。后来陆续发现黑麦、小麦、玉米等的根系分泌物阿凡酸(avenic acid)、啤麦根酸(disticonic acid)等也具有相似功能[32]。这些物质在结构功能上与微生物分泌的铁载体相似,所以将它们统称为植物铁载体(PS)。MAs合成途径见图3[33]。烟酰胺(nicotianamine,NA)在尼克酞胺氨基转移酶(nicotianamine aminotransferase,NAAT)[34]的催化下,形成一种酮类的中间产物,然后经脱氧麦根酸合成酶(2′-deoxymugeneic acid synthase,DMAS)[35]的还原产生 2-脱氧麦根酸。水稻、小麦和玉米只能分泌2-脱氧麦根酸(2′-deoxymugeneic acid,DMA),而大麦能够分泌多种形式的麦根酸,所以大麦具有更强的缺铁耐受性[35]。
研究表明,尼克酞胺合成酶(nicotianamine synthase,NAS)[33]、尼克酞胺氨基转移酶、脫氧麦根酸合成酶(2′-deoxymugeneic acid synthase,DMAS)是PS合成的限速酶[25]。目前已经克隆到3个NAS基因(OsNAS1-3)[25,33]。在铁充足条件下,OsNAS1和OsNAS2仅在根中表达,OsNAS3主要在的叶中表达;但在缺铁条件下,OsNAS1和OsNAS2在根和叶中都强烈表达,OsNAS3基因在根中的表达略微增强,而在叶中的表达受到抑制[33]。
3.2.2Fe2+-PS螯合物的运输
水稻OsTOM1和大麦HvTOM1负责转运铁载体到根部[36]。PS鳌合溶解的Fe3+形成Fe3+-PS鳌合物[25]。玉米YS(yellow stripe)转运体负责将Fe3+-PS螯合物运输到根部[7]。水稻有18个YSL(yellow stripe-like)基因,只有OsYSL15具有运输Fe3+-PS螯合物到根部的功能[25,37](图4)。
IDE1、IDE2是首先被克隆的缺铁感应元件[38]。IDE结合[CM(25]因子IDEF1和IDEF2,在水稻根、叶中组成性表达[18,39]。
缺铁应答基因OsIRO2是bHLH转录因子,特异性结合CACGTGG序列[39-40]。OsIRO2调节Fe(Ⅲ)-MA运载体和MAs合成途径相关基因(OsNAS1、OsNAS2、OsNAAT1、OsDMAS1等)的表达[18]。OsIRO3负向调节缺铁应答基因[41]。OsbHLH133调节根茎之间铁的分布[3]。
4铁由根到茎的运输
铁在水稻体内主要通过木质部和韧皮部进行运输,其中韧皮部的铁运输到新叶中,木质部的铁大多运输到老叶中[42]。水稻体内大部分铁以铁蛋白质或螯合物的形式存在[25]。NA、MAs、柠檬酸(citrate)、酚类物质(phenolics)是重要的植物螯合剂,其中MAs是禾本科植物特有的螯合剂[25]。
4.1铁在木质部的运输
在木质部的长距离运输中,铁离子主要与柠檬酸螯合[25]。OsFRDL1是一个柠檬酸转运子,可以高效运输水稻根系的铁[43]。OsFRDL1属于多药及毒性化合物外排家族的基因。OsFRDL1敲除突变体osfrdl1根部石柱细胞铁含量高,但叶片铁含量很低,导致叶片失绿,osfrdl1木质部中柠檬酸和Fe3+浓度远低于对照。说明OsFRDL1主要负责在木质部运输柠檬酸。OsFRDL1主要在长距离运输细胞中表达,也在生殖器官中表达,且不受缺铁信号调控[44]。
OsYSL2是Fe(Ⅱ)-NA和Mn(Ⅱ)-NA转运体基因,该基因沉默降低了铁由根向地上部分的转运及在结实过程中向种子中的转运,因而推测该基因可能参与了Fe(Ⅱ)-NA在木质部中的长距离运输[45-46]。
原儿茶酸(protocatechuic acid,PCA)、咖啡酸(caffeic acid,CA)等酚类物质能够溶解、鳌合质外体沉积的Fe3+,并将其还原成Fe2+[47]。PCA和CA的浓度在水稻OsPEZ1(phenolics efflux zero 1,PEZ1)基因突变体的木质部液中显著降低;爪蟾卵母细胞中表达OsPEZ1后能够转运PCA[48]。这些研究结果说明,OsPEZ1负责转运PCA进入木质部,调节质外体铁平衡。OsPEZ1和OsPEZ2(efflux transporters,phenolics efflux zero1和2)也属于MATE转运体家族[10,30,49]。
4.2铁在韧皮部的运输
水稻韧皮部铁主要是Fe(Ⅲ)-DMA[50],Fe(Ⅱ)-NA的含量相对较少。OsYSL2是Mn-NA和Fe-NA运载体编码基因[45],該基因不但参与木质部铁运输,还在韧皮部Fe、Mn运输中发挥重要功能,说明Fe(Ⅱ)-NA对水稻生长发育也很重要[45-46]。OsYSL15是根部负责吸收Fe(Ⅲ)-DMA的转运体基因,但该基因也在维管组织中强烈表达,说明该基因也参与水稻体内铁运输[37,51]。OsYSL18是Fe(Ⅲ)-DMA的转运载体基因,在花粉、花粉管、叶片和叶鞘接头部位的韧皮部中表达,说明该基因很可能在水稻受精和韧皮部的铁运输中起作用[52]。OsYSL16编码产物参与体内尤其是生殖器官中Fe(Ⅲ)-DMA的分布及转导,还负责运输Cu-NA,调节体内Cu分布[53]。
5展望
铁是生物体必需的微量元素,铁含量不足破坏代谢平衡、危害严重,但铁含量过高容易引起铁中毒,所以生物体进化出精密的机制严格调控体内铁含量。表1对比了各种强化途径的优缺点。生物强化的花费主要集中在前期品种选育上,不需要持续的资金支持,不增加消费者的负担[16]。统计显示,一个75亿的项目一年能为375亿人提供铁生物强化水稻(http://www.biokemi.org/biozoom/issues/525/articles/2392)。2001年,在亚洲国际发展银行和国际食物政策研究所(ADB-IFPRI)的支持下,通过9个月的人群干预研究发现,人体内铁的贮存量在富铁水稻品种IR68144-3B-2-2-3干预下增加20%[54]。水稻是人类不可或缺的主食,即使稻米的铁含量只有微量增加,日积月累也会对人体健康起到深远的影响[55]。综上所述,水稻铁生物强化是世界人口,尤其是贫困人口获得足够膳食营养的最经济、最有效途径,意义深远[4]。
不同水稻品种资源间的铁含量存在很大的遗传差异[56-57],所以科研人员能够通过育种手段培育富铁新品种。日本水稻品种GCN4和系026铁含量比普通水稻铁含量高出3~6倍[58]。吴敬德等培育的IR41994-SO-2-1-3(S1)、IR68144-213-2-2-3(S5)铁含量分别为42.86、 36.28 mg/kg,均高于普通品种[59]。李琬等培育的富铁品系中龙粳201的3年精米平均铁含量为12.65 mg/kg,是普通品种的3~4倍(未发表)。
水稻铁浓度受环境因素影响大[56,60],且测定成本高。因此,育种者在常规育种的基础上,须要结合分子标记辅助育种技术、转基因技术挖掘和创制富铁新种质,且应加强提高主栽品种精米铁含量和培育广适性富铁品种。培育富铁水稻实际上就是提高水稻对外部环境中铁的吸收、运输和积累能力的过程。因此,研究水稻对铁的吸收、运输、积累机制及其相关控制基因,对铁生物强化育种具有重要意义。
参考文献:
[1]Meng F H,Wei Y Z,Yang X E. Iron content and bioavailability in rice[J]. Journal of Trace Elements in Medicine & Biology,2005,18(4):333-338.
[2]Guerinot M L,Yi Y. Iron:nutritious,noxious,and not readily available[J]. Plant Physiology,1994,104(3):815-820.
[3]王璐. 缺铁响应转录因子OsbHLH133的功能和缺铁诱导乙烯合成分子机理的研究[D]. 杭州:浙江大学,2013.
[4]Sperotto R A,Ricachenevsky F K,de Abreu W V,et al. Iron biofortification in rice:its a long way to the top[J]. Plant Science,2012,190:24-39.
[5]R-mheld V,Marschner H. Evidence for a specific uptake system for iron phytosiderophores in roots of grasses[J]. Plant Physiology,1986,80(1):175-180.
[6]Bell W D,Bogorad L,McIlrath W J. Yellow-stripe phenotype in maize Ⅰ. Effects of ys1 locus on uptake and utilization of iron[J]. Botanical Gazette,1962,124(1):1-8.
[7]Curie C,Panaviene Z,Loulergue C,et al. Maize yellow stripe1 encodes a membrane protein directly involved in Fe(Ⅲ) uptake[J]. Nature,2001,409(6818):346-349.
[8]Ishimaru Y,Suzuki M,Tsukamoto T,et al. Rice plants take up iron as an Fe3+ -phytosiderophore and as Fe2+[J]. Plant Journal,2006,45(3):335-346.
[9]Cheng L J,Wang F,Shou H X,et al. Mutation in nicotianamine aminotransferase stimulated the Fe(Ⅱ) acquisition system and led to iron accumulation in rice[J]. Plant Physiol,2007,145(4):1647-1657.
[10]Bashir K,Ishimaru Y,Shimo H,et al. Rice phenolics efflux transporter 2 (PEZ2) plays an important role in solubilizing apoplasmic iron[J]. Soil Science and Plant Nutrition,2011,57(6):803-812.
[11]Xiong H,Kakei Y,Kobayashi T,et al. Molecular evidence for phytosiderophore‐induced improvement of iron nutrition of peanut intercropped with maize in calcareous soil[J]. Plant,Cell & Environment,2013,36(10):1888-1902.
[12]Marschner H. Mineral nutrition of higher plants[M]. 2nd ed. London:Academic Press,1995.
[13]Kobayashi T,Itai R N,Aung M S,et al. The rice transcription factor IDEF1 directly binds to iron and other divalent metals for sensing cellular iron status[J]. The Plant Journal,2012,69(1):81-91.
[14]Kobayashi T,Nagasaka S,Senoura T,et al. Iron-binding haemerythrin RING ubiquitin ligases regulate plant iron responses and accumulation[J]. Nature Communications,2013,4:2792.
[15]Long T A. The bHLH transcription factor POPEYE regulates response to iron deficiency in Arabidopsis roots[J]. Plant Cell,2010,22(7):2219-2236.
[16]Kobayashi T,Nishizawa N K . Iron uptake,translocation,and regulation in higher plants[J]. Annual Review of Plant Biology,2012,63(1):131-152.
[17]Hindt M N,Guerinot M L. Getting a sense for signals:regulation of the plant iron deficiency response[J]. BBA-Molecular Cell Research,2012,1823(9):1521-1530.
[18]Kobayashi T,Ogo Y,Itai R N,et al. The transcription factor IDEF1 regulates the response to and tolerance of iron deficiency in plants[J]. Proceedings of the National Academy of Sciences,2007,104(48):19150-19155.
[19]Kakei Y,Ogo Y,Itai R N,et al. Development of a novel prediction method of cis-elements to hypothesize collaborative functions of cis-element pairs in iron-deficient rice[J]. Rice,2013,6(1):22.
[20]Kobayashi T,Ogo Y,Aung M S,et al. The spatial expression and regulation of transcription factors IDEF1 and IDEF2[J]. Annals of Bbotany,2010,105(7):1109-1117.
[21]Zhang L X,Itai R N,Yamakawa T,et al. The bowman-birk trypsin inhibitor IBP1 interacts with and prevents degradation of idef1 in rice[J]. Plant Molecular Biology Reporter,2014,32(4):841-851.
[22]Long T A,Tsukagoshi H,Busch W,et al. The bHLH transcription factor POPEYE regulates response to iron deficiency in Arabidopsis roots[J]. The Plant Cell,2010,22(7):2219-2236.
[23]Kobayashi T,Nishizawa N K. Iron sensors and signals in response to iron deficiency[J]. Plant Science,2014;224(13):36-43.
[24]Wu J J,Wang C,Zheng L Q,et al. Ethylene is involved in the regulation of iron homeostasis by regulating the expression of iron-acquisition-related genes in Oryza sativa[J]. Journal of Experimental Botany,2010,62(2):667-674.
[25]Bashir K,Nozoye T,Ishimaru Y,et al. Exploiting new tools for iron bio-fortification of rice[J]. Biotechnology Advances,2013,31(8):1624-1633.
[26]Qi Y H,Wang S K,Shen C J,et al. OsARF12,a transcription activator on auxin response gene,regulates root elongation and affects iron accumulation in rice (Oryza sativa)[J]. New Phytologist,2012,193(1):109-120.[HJ1.5mm]
[27]Robinson N J,Procter C M,Connolly E L,et al. A ferric-chelate reductase for iron uptake from soils[J]. Nature,1999,397(6721):694-697.
[28]Bughio N,Yamaguchi H,Nishizawa NK et al. Cloning an iron-regulated metal transporter from rice[J]. J Exp Bot,2002,53:1677-1682.
[29]Ishimaru Y,Bashir K,Nishizawa N K. Zn uptake and translocation in rice plants[J]. Rice,2011,4(1):21-27.
[30]Ishimaru Y,Kakei Y,Shimo H,et al. A rice phenolic efflux transporter is essential for solubilizing precipitated apoplasmic iron in the plant stele[J]. Journal of Biological Chemistry,2011,286(28):24649-24655.
[31]Takahashi R,Ishimaru Y,Senoura T,et al. The OsNRAMP1 iron transporter is involved in Cd accumulation in rice[J]. Journal of Experimental Botany,2011,62(14):4843-4850.
[32]常正堯. 水稻缺铁胁迫下渗透酶基因的克隆、亚细胞定位及膜泡运输相关基因的分析[D]. 北京:首都师范大学,2006.
[33]Inoue H,Higuchi K,Takahashi M et al. Three rice nicotianamine synthase genes,OsNAS1,OsNAS2,and OsNAS3 are expressed in cells involved in long-distance transport of iron and differentially regulated by iron[J]. Plant Journal,2003,36(3):366-381.
[34]Inoue H,Takahashi M,Kobayashi T,et al. Identification and localisation of the rice nicotianamine aminotransferase gene OsNAAT1 expression suggests the site of phytosiderophore synthesis in rice[J]. Plant Molecular Biology,2008,66(1/2):193-203.
[35]Bashir K,Nishizawa N. Deoxymugineic acid synthase:a gene important for Fe-acquisition and homeostasis[J]. Plant Signaling & Behavior,2006,1(6):290-292.
[36]Nozoye T,Nagasaka S,Kobayashi T,et al. Phytosiderophore efflux transporters are crucial for iron acquisition in graminaceous plants[J]. Journal of Biological Chemistry,2011,286(7):5446-5454.
[37]Lee S,Chiecko J C,Kim S A,et al. Disruption of OsYSL15 leads to iron inefficiency in rice plants[J]. Plant Physiology,2009,150(2):786-800.
[38]Kobayashi T,Nakayama Y,Itai R N,et al. Identification of novel cis-acting elements,IDE1 and IDE2,of the barley IDS2 gene promoter conferring iron‐deficiency‐inducible,root-specific expression in heterogeneous tobacco plants[J]. The Plant Journal,2003,36(6):780-793.
[39]Ogo Y,Nakanishi Itai R,Nakanishi H,et al. The rice bHLH protein OsIRO2 is an essential regulator of the genes involved in Fe uptake under Fe‐deficient conditions[J]. The Plant Journal,2007,51(3):366-377.
[40]Ogo Y,Itai R N,Kobayashi T,et al. OsIRO2 is responsible for iron utilization in rice and improves growth and yield in calcareous soil[J]. Plant Molecular Biology,2011,75(6):593-605.
[41]Zheng L,Cheng Z,Ai C,et al. Nicotianamine,a novel enhancer of rice iron bioavailability to humans[J]. PLoS One,2010,5(4):e10190.
[42]Tsukamoto T,Nakanishi H,Uchida H,et al. 52Fe translocation in barley as monitored by a positron-emitting tracer imaging system (PETIS):evidence for the direct translocation of Fe from roots to young leaves via phloem[J]. Plant and Cell Physiology,2008,50(1):48-57.
[43]沈宏,杨旭健,傅友强. 一個水稻铁转运基因(OsFRDL1)参与缺氧诱导根系铁膜形成的调节过程[J]. 科学通报,2014,59(9):787-795.
[44]Inoue H,Mizuno D,Takahashi M,et al. A rice FRD3-like (OsFRDL1) gene is expressed in the cells involved in long-distance transport[J]. Soil Science and Plant Nutrition,2004,50(7):1133-1140.
[45]Koike S,Inoue H,Mizuno D,et al. OsYSL2 is a rice metal-nicotianamine transporter that is regulated by iron and expressed in the phloem[J]. The Plant Journal,2004,39(3):415-424.
[46]Ishimaru Y,Masuda H,Bashir K,et al. Rice metal-nicotianamine transporter,OsYSL2,is required for the long‐distance transport of iron and manganese[J]. The Plant Journal,2010,62(3):379-390.
[47]Yoshino M,Murakami K. Interaction of iron with polyphenolic compounds:application to antioxidant characterization[J]. Analytical Biochemistry,1998,257(1):40-44.
[48]张会敏. 水稻铁稳态正调控因子OsPRI1促进铁平衡[D]. 合肥:中国科学技术大学,2018.
[49]Ishimaru Y,Bashir K,Nakanishi H,et al. The role of rice phenolics efflux transporter in solubilizing apoplasmic iron[J]. Plant Signaling & Behavior,2011,6(10):1624-1626.
[50]Nishiyama R,Kato M,Nagata S,et al. Identification of Zn-nicotianamine and Fe-2′-deoxymugineic acid in the phloem sap from rice plants (Oryza sativa L.)[J]. Plant and Cell Physiology,2012,53(2):381-390.
[51]Inoue H,Kobayashi T,Nozoye T,et al. Rice OsYSL15 is an iron-regulated iron (Ⅲ)-deoxymugineic acid transporter expressed in the roots and is essential for iron uptake in early growth of the seedlings[J]. Journal of Biological Chemistry,2009,284(6):3470-3479.
[52]Aoyama T,Kobayashi T,Takahashi M,et al. OsYSL18 is a rice iron (Ⅲ)-deoxymugineic acid transporter specifically expressed in reproductive organs and phloem of lamina joints[J]. Plant Molecular Biology,2009,70(6):681-692.
[53]Zheng L,Yamaji N,Yokosho K,et al. YSL16 is a phloem-localized transporter of the copper-nicotianamine complex that is responsible for copper distribution in rice[J]. The Plant Cell,2012,24(9):3767-3782.
[54]Haas J D,Beard J L,Murray-Kolb L E,et al. Iron-biofortified rice improves the iron stores of nonanemic Filipino women[J]. The Journal of Nutrition,2005,135(12):2823-2830.
[55]Gregorio G B,Senadhira D,Htut T. Improving iron and zinc value of rice for human nutrition[J]. Agriculture et Developpement,1999(23):77-81.
[56]Gregorio G B,Senadhira D,Htut H,et al. Breeding for trace mineral density in rice[J]. Food and Nutrition Bulletin,2000,21(4):382-386.
[57]曾亞文,刘家富,汪禄祥,等. 云南稻核心种质矿质元素含量及其变种类型[J]. 中国水稻科学,2003,17(1):26-31.
[58]董彦君. 日本新性状稻米品质研究进展[J]. 中国稻米,1998(1):36-38.
[59]吴敬德,郑乐娅,张瑛,等. 富含铁锌水稻的筛选[J]. 安徽农业科学,2006,34(4):635.
[60]Cabuslay G S,Sison C B,Laureles E,et al. Grain mineral density:nitrogen response and seasonal variation[J]. Workshop on Rice Breeding for Better Nutrition,2003(4):7-11.
猜你喜欢
化学工程师(2022年11期)2022-12-17
中国化妆品(2020年2期)2020-04-13
中国化妆品(2019年1期)2019-04-18
分析仪器(2018年3期)2018-06-07
中国洗涤用品工业(2017年6期)2017-08-22
湖北师范大学学报(自然科学版)(2017年1期)2017-06-27
西华师范大学学报(自然科学版)(2015年3期)2015-02-27
天津药学(2015年2期)2015-02-23
药学与临床研究(2014年3期)2014-03-06
无机化学学报(2014年6期)2014-02-28
