
玉米脱镁叶绿酸氧化酶基因ZmPAO表达与叶绿素含量动态变化的关联分析
2019-08-14 07:47韦宁宁聂佳伟王亚辉赵志鑫张兴华薛吉全徐淑兔
植物营养与肥料学报 2019年7期
韦宁宁,聂佳伟†,王亚辉,李 婷,赵志鑫,张兴华,薛吉全,徐淑兔*
(1 西北农林科技大学农学院/农业部西北旱区玉米生物学与遗传育种重点试验室,杨凌,712100;2 陕西省玉米工程技术研究中心,杨凌,712100)
叶绿素是自然界最重要的光合色素之一,植物中的叶绿素主要为叶绿素a和叶绿素b[1],以激发形式将光子能量转变成化学能[2]。在籽粒成熟后期,有效的叶绿素分解可以促进营养物质向贮存器官转移,是制约玉米产量进一步提高的重要因素[3-4]。挖掘成熟后期的玉米脱镁叶绿酸氧化酶基因 (ZmPAO) 的有效变异位点有助于理解叶绿素的代谢机制,为玉米分子遗传育种提供一定的理论基础。
自从Hendry等35年前提出叶绿素降解是生物学之谜后[5],叶绿素降解机制得到了不断发展。在模式植物拟南芥中,叶绿素代谢通路通过分子生物学,生理学和代谢组学等手段已经逐渐明晰[6-8]。植物体内的叶绿素水平主要由叶绿素循环[9-10]和叶绿素降解[11-12]所调节。研究认为“PAO”途径完全决定了叶绿素酶促降解过程,叶绿素b通过向叶绿素a的转化[13],进而实现叶绿素b的唯一降解途径[14]。叶绿素a的分解始于被叶绿素酶 (CLH) 催化形成脱植基叶绿素,再由一个金属螯合物脱去镁离子形成脱镁叶绿酸a (pheide a),然后通过脱镁叶绿酸氧化酶(PAO),叶绿素酶 (RCC) 开启了卟啉环的裂解,再通过红色叶绿素代谢产物还原酶 (RCCR) 进一步分解,生成初生荧光叶绿素代谢产物 (pFCC)[15-17],并最终经过几次修饰之后运输至液泡中转化为非荧光的叶绿素分解代谢物 (NCC)[18](图1)。这些酶蛋白的结构在20多种植物都得到了分离和鉴定[19-21]。
在拟南芥中,PAO突变体曾被定义为加速细胞死亡1 (ACD1)[22],PAO的表达有助于拟南芥抵抗针叶假单胞菌的感染[23]。在过去的20年里,Kräutler等人第一次阐述了叶绿素代谢在藻胆素中的相关结构[24]和后续调解机制[25]。通过后续鉴定得到了一种打开脱镁叶绿酸a卟啉环的加氧酶PAO[26],其C端跨膜结构域被定位在类囊体膜上[27],是“PAO”代谢途径的重要酶[28-29]。作为叶绿素降解关键基因,挖掘脱镁叶绿酸氧化酶 (PAO) 基因中与玉米成熟后期叶绿素组分含量的有利变异位点对解析玉米叶片衰老具有重要意义。因此,本研究以源自陕A群、陕B群选育的自交系和国内骨干自交系共141份玉米自交系为材料,开展ZmPAO基因序列分析和该基因与成熟后期叶绿素组分含量的关联分析,拟找到ZmPAO的有效功能位点,为解释该基因结构提供一定的研究基础,可以提高我们对控制叶绿素含量变异这一数量性状的遗传机制的理解,以服务功能保绿型的玉米分子育种。

图1 叶绿素代谢通路图Fig. 1 Chlorophyll metabolic pathway
1 材料与方法
1.1 试验材料与田间种植
本试验研究材料来自于西北农林科技大学提供的141份玉米自交系(来自A、B两个自交系群体)。试验于2017年在西北农林科技大学榆林综合试验站和杨凌综合试验站进行。试验采取随机区组实验设计,每个自交系种植2行,行长5 m,行距60 cm,种植密度75000 株/hm2,3次重复,正常田间浇水管理。
1.2 表型测定与分析
在玉米开花后20天(T1)起开始取样,依次为T2 (开花后 26天)、T3 (开花后 32天)、T4 (开花后38 天)、T5 (开花后 44 天)、T6 (开花后 50 天) 和 T7(开花后56天)。取样过程为用直径0.6 cm的打孔器从两地随机取每小区生长状况一致的玉米穗位叶中部10枚圆片放入2 mL离心管,各做3个重复,立即与1 mL 95%乙醇混合。在4 ℃黑暗条件下处理48 h,用分光光度计 (Bio-RAD,上海,中国) 测叶绿素提取上清液在645 nm和663 nm的波长处的吸光度,取3个重复的平均值为最终叶绿素含量。
Chl. a和Chl. b的计算采用Arnon[30]提出的下列公式:

所有数据分析使用JMP 10软件[31]和Microsoft Excel。
1.3 ZmPAO基因克隆
模式植物拟南芥叶绿素代谢通路及相关基因的功能已有较好的研究基础,因此本研究通过拟南芥基因数据库 (https://www.arabidopsis.org/) 搜索关键词PAO,根据基因注释信息确定AT3G44880是叶绿素代谢中的关键氧化还原酶。利用拟南芥中该基因蛋白序列信息在EnsemblPlants网站 (http://plants.ensembl.org/index.html) 比对玉米基因组序列信息利用tBLASTn,通过控制E值 (E value ≤ 3.8E-6) 得到玉米中脱镁叶绿酸氧化酶同源基因Zm00001d027656,命名为ZmPAO。
1.4 序列分析
通过使用改良的CTAB方法[32]从141个玉米自交系的4叶期幼苗的新鲜叶中提取基因组DNA。使用分光光度计Nanodrop 2000 (Bio-Rad公司,上海,中国) 和1%的凝胶电泳确定浓度和质量。以参考基因组 B73 (http://www.maizesequence.org) 提供的ZmPAO基因序列为基础通过primmer 3.0 (http://primer3.ut.ee/) 在线应用设计和筛选引物,用于关联分析中的该基因的重测序。基因测序结果使用Muscle (http://www.ebi.ac.uk/Tools/msa/musc) 和BioEdit进行多序列比对。
1.5 关联分析
对实验室已有的141份材料的简化全基因组序列使用STRUCTRE估算玉米自交系的群体结构。使用SPAGeDi计算相对亲缘关系矩阵[33]。使用TASSEL[34]提取141个玉米品系中具有最小等位基因频率 (MAF) ≥ 0.05的ZmPAO基因中的SNPs和InDels位点。根据TASSEL计算两个位点之间的连锁不平衡情况 (LD)[34],排除显著连锁 (R2〉 0.2) 的位点,以各时期各个材料的叶绿素组分含量为表型数据,筛选最显著的位点以挖掘该基因有效功能位点并进行性状的单倍型分析[35]。
2 结果与分析
2.1 叶绿素组分含量基本统计分析
在榆林和杨凌试验站正常管理下,对141份玉米自交系在7个取样时间点的叶绿素组分含量通过使用JMP 10软件进行了基本统计分析。相关性分析结果 (图2) 表明,在同一地点同一时期叶绿素a和叶绿素b的相关性为0.956,只有榆林的花后44天和杨凌的花后38天的相关系数相对较低,分别为0.303和0.527。不同地点或不同时期间的叶绿素a和叶绿素b的正相关性较低,说明叶绿素组分含量间的密切程度较低,有较大的独立变化性。这也说明了叶绿素组分含量受环境和基因型的影响较为敏感,处于时刻变化中。除榆林试验站的花后26天和杨凌试验站的花后26天和38天的叶绿素组分含量的峰度和偏度较大外,其他叶绿素组分含量基本符合正态分布 (表1),表明叶绿素含量是数量性状,其测定值可用于后续关联分析。

图2 榆林和杨凌地区7个取样时期玉米穗位叶叶绿素a和叶绿素b含量的相关性分析Fig. 2 Correlation of chlorophyll a and b contents at the seven sampling periods in Yulin and Yangling regions
2.2 不同取样时期叶绿素组分含量变化分析
杨凌和榆林试验站正常田间管理下,7个取样时期141份材料的叶绿素a和叶绿素b含量平均值的系列波动说明了玉米在成熟后期叶绿素的动态变化和衰老过程。榆林在开花后38天总叶绿素含量最高为4.58 mg/L,开花后56天最低为3.09 mg/L,差异为48.22%;杨凌处理在开花后44天总叶绿素最高含量为4.52 mg/L,开花后26天最低为3.05 mg/L,差异为48.19%,开花后56天的总叶绿素含量仍较高,为3.82 mg/L。这一变化说明榆林和杨凌地区在玉米成熟后期的总叶绿素变化幅度基本一致,主要由材料本身决定;不同的是杨凌比榆林的玉米生育期长,叶绿素合成和降解速度较慢。图3的叶绿素组分变化结果表明,玉米成熟后期叶绿素a含量普遍低于叶绿素b含量,并不符合传统3∶1的比值;后期叶绿素b均呈下降趋势,说明叶绿素b可能正通过向叶绿素a转化实现自身降解;除此之外,取样后期的总叶绿素含量也在降低。玉米成熟后期,总叶绿素的降解在普遍发生,进一步表明叶绿素代谢关键基因ZmPAO可能起着重要的作用,符合关联分析对于叶绿素代谢过程的要求。
2.3 ZmPAO的核苷酸多样性
ZmPAO有两个转录本Zm00001d027656_T001和Zm00001d027656_T002。前者氨基酸序列较长,由七个外显子组成,外显子跨越染色体1上的4349 bp∶10,261,506-10,265,855 (图 4),它含有2131 bp的CDS长度,编码520个氨基酸序列,故以其进行序列分析。通过PCR (聚合酶链式反应)扩增、产物电泳检测与测序分析最终筛选了6对引物用于141份自交系群体目的基因的测序 (引物序列详见表2,引物所在基因位置详见图4)。通过141份玉米自交系ZmPAO的重测序和多重序列比对,共检测到108个多态位点。它包括72个单核苷酸多态性(single nucleotide polymorphism,SNP) 位点和36个插入缺失 (insertion/deletion,InDel) 位点,平均每40 bp区间内检测到一个多态位点。ZmPAO的核苷酸多样性分布不均,第3内含子的核苷酸多样性水平最高 (表 3)。

表1 榆林和杨凌玉米各取样时期叶绿素a和b含量的群体特征 (mg/L)Table 1 Population characteristics of chlorophyll a and b contents in Yulin and Yangling regions
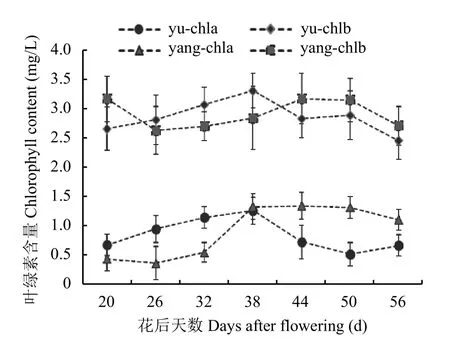
图3 榆林和杨凌玉米花后7个取样期穗位叶叶绿素含量Fig. 3 Contents of chlorophyll a and b in the seven sampling periods in Yulin and Yangling
2.4 ZmPAO与叶绿素组分含量的有效关联位点分析
利用群体结构和亲缘关系为基础的混合线性模型对已鉴定的多态位点和各时期叶绿素含量进行关联分析。经LD分析,该基因108个多态性位点中有19个(13个SNPs和6个Indels)与叶绿素化合物显著相关 (P〈 0.05),有12个多态性位点与两个或更多的性状相关。在19个位点中,1个SNP位点位于3′UTR,14个变异位点位于内含子区域;只有4个SNP位点位于外显子区域,其中有2个引起了氨基酸的变化 (丙氨酸A-苏氨酸T,脯氨酸P-丝氨酸S),且都为疏水氨基酸到亲水氨基酸的变化 (表3)。这说明ZmPAO基因对于叶绿素的影响既存在蛋白水平的调控也存在转录水平的调控。
榆林点关联分析共找到12个显著变异位点(6个SNPs和6个Indels),有趣的是所有的Indels位点都对其叶绿素组分含量起一定的作用,其中SNP位点S528、S545、S1390和Indel位点S1144对于同时期的叶绿素a和叶绿素b都存在贡献,但花后32天和38天没有找到相关有效变异位点。变异位点对于叶绿素组分含量的表型解释率在3.89%~8.70%,分别作用于花后56天的叶绿素b和26天的叶绿素a。以作用于花后20天和56天的叶绿素组分表型变异的关联位点数较多,为5个 (2个SNPs和3个Indels) 和4个 (2个SNPs和2个Indels);花后20天的叶绿素a的表型总解释率高达25.77%,同时期叶绿素b的表型总贡献率为19.16%;花后56天的叶绿素a和叶绿素b的表型总解释率分别为9.98%和18.11%。其他时期以1个变异位点为主,花后26天的SNP位点S528 (G/A) 对叶绿素a和叶绿素b分别有8.7%和6.77%的表型贡献;花后44天的Indel位点S996解释了8.09%的叶绿素b表型变异;花后50天的SNP位点S3232 (A/C) 和Indel位点S3235分别解释叶绿素a的4.92%和6.41%变异。

图4 ZmPAO的基因结构Fig. 4 Structure of gene ZmPAO

表2 ZmPAO基因的引物序列Table 2 Primer sequences for re-sequencing of ZmPAO gene using PCR
杨凌点关联分析结果共找到11个有效变异位点(9个SNPs和2个Indels)。各时期叶绿素a和叶绿素b表型变异相关的各有8个变异位点,其中SNP位点S3182和S3227分别只对开花后44天和56天的叶绿素a起到5.24%和9.40%的效应,SNP位点S3569以及Indel位点S3235只与花后50天的叶绿素b相关,表型总效应高达23.23%。花后26天和50天的叶绿素组分含量各有5个相关变异位点,有趣的是对叶绿素b表型变异都有较高总解释率,分别为18.48%和35.36%,与叶绿素a的效应相对较低 (前者为6.13%,后者为6.42%)。花后20天和38天都各有一个相关变异位点,分别为SNP位点S3675和S3817,其中S3675的效应值相对较大,均在12%以上。花后32天的SNP位点S1001、S1352、S1602对叶绿素b表型变异的效应值均为7.73%。
选取榆林开花后20天的叶绿素b等表型性状的显著关联位点进行单倍型分析 (表4),结果表明同一性状的不同单倍型在陕西A、B群中的差异性并不相同,榆林花后20天叶绿素b的单倍型1在B群中表现突出,而单倍型2恰好相反。但杨凌花后26天叶绿素b的两种单倍型间存在显著差异,在A群中的表型性状普遍较高,其中单倍型2的有效位点与表3中的变异位点偏好相同。榆林花后56天叶绿素a的单倍型1和2间的差异显示具体位点碱基缺失对于表型性状可能有利,榆林花后20天叶绿素b的单倍型2和3中也有同样发现。
3 讨论与结论

表3 榆树和杨凌区各取样时期玉米叶绿素a和b的关联分析Table 3 Correlation of chlorophyll a and b in different stages of maize in Yulin and Yangling regions
已有研究表明依靠作物全生育期光能利用率的提高可以进一步提高作物产量[36]。玉米生长后期是玉米产量形成的关键时期,人们以生育后期保持较高的叶绿素含量为标准选育长效光合持续期的品种取得一定进展[37]。植物叶绿素含量直接影响作物后期的光合作用和物质向果实的运输积累,干旱胁迫下对于作物的生存和产量形成更为重要[38-39]。随着技术的研究,关联分析的应用加快了复杂数量性状遗传机理的研究,但是关于玉米中叶绿素代谢相关功能等位基因的报道很少,大量的研究基于QTL初定位,明确了影响叶绿素代谢的区段,但未能找到真正的有效功能位点[40-43]。为此,通过研究叶绿素代谢通路的关键酶PAO的具体变异位点具有一定的创新性,以关联分析确定显著效应位点为后期该基因功能位点编辑提供分子基础。
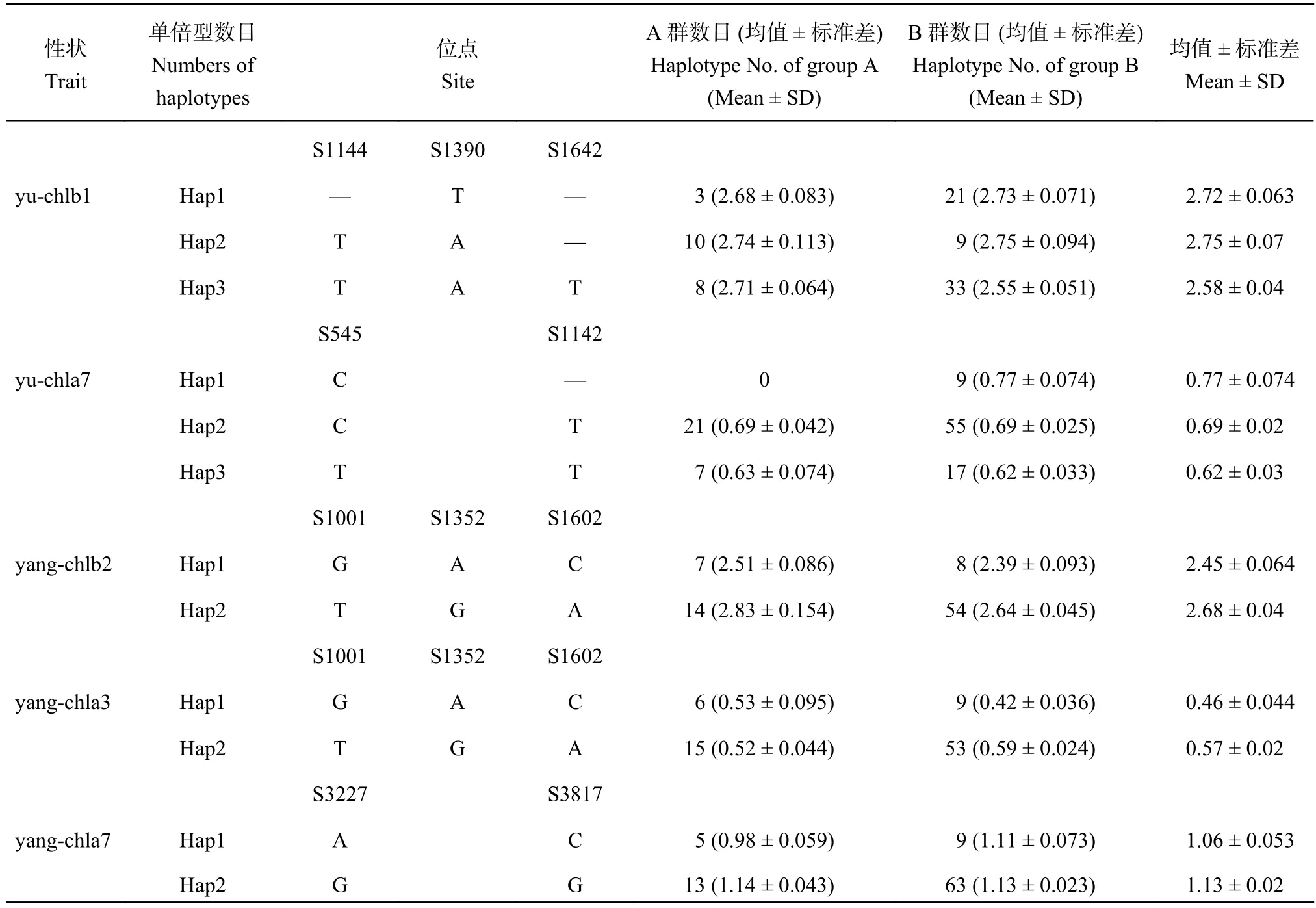
表4 性状显著变异位点的单倍型分析Table 4 Haplotype analysis of significantly varied loci in traits
本研究以西北农林科技大学141个玉米自交系的两个环境7个时期叶绿素组分含量为表型数据挖掘ZmPAO基因的功能位点。结果显示,叶绿素b的含量都明显高于同时期的叶绿素a的含量,不符合植物叶片叶绿素a∶b为3∶1[44],这可能与玉米成熟后期叶绿素a的降解快于叶绿素b向叶绿素a的转化速度有关。另外,榆林地区的总叶绿素变化幅度和杨凌地区相似,但杨凌的材料相对持绿性更久,可能与两地的播期相关 (榆林为玉米春播区、杨凌为夏播区),后者的前期生长时间较短,故授粉后44天杨凌的叶绿素各组分含量普遍高于榆林,总体呈下降趋势。
关联结果表明,有8个有效多态位点 (4个SNPs和4个Indels) 只与榆林地区的叶绿素组分含量变异相关,其中SNP位点S528、S545和S1390以及Indel位点S1144对同时期叶绿素a和叶绿素b都存在效应;有7个SNPs只对杨凌的叶绿素组分含量变异具有贡献效应。剩下的4个变异位点 (2个SNPs和2个Indels) 对两地叶绿素组分含量变异都存在作用,说明这些位点的效应可能存在普遍性,后期需要进一步进行验证。对于杨凌各时期的叶绿素组分变异的总效应值分布普遍高于榆林地区,前者总效应值最高的是花后50天叶绿素b的41.78%总表型解释率,后者最高的是花后20天的叶绿素a的25.77%的总贡献率。性状单倍型分析的有利位点与关联分析显著位点的偏好一致,说明MLM模型的关联分析结果具有可靠性。本研究发现ZmPAO基因中与玉米成熟后期穗位叶叶绿素组分含量相关的有效关联位点多以非氨基酸变异位点为主,氨基酸变异位点S528和S545都只与榆林地区花后26天和56天的叶绿素组分变异相关。这些现象的出现可能基于以下原因:1) 试验使用的是B73参考基因组中的ZmPAO序列,与141份材料的实际基因序列相比可能存在结构划分的不同,特别是可变剪切[45]现象的存在,导致一些DNA序列被误认为并不起编码作用;2) 基因调控除了蛋白水平的调控还存在转录水平的调控,本研究测序基因范围可能未将真正的功能位点涵盖,需要更深入的研究;3) 内含子变异不仅能参与结构基因的再剪接,也能参与启动子启动活性和启动子内含的暗诱导元件诱导活性的调节[46],特别是3′UTR处S3817的SNP位点可能影响相应的miRNA的结合强度[47]而调控基因表达。
本研究结果对进一步解析玉米叶绿素代谢机理有重要的指导意义,为玉米成熟后期适度衰老品种的创造提供遗传机制,但不易排除叶绿素含量其他相关基因的作用;后期可以通过组学手段研究该基因的表达调控网络,从而在全基因组表达水平揭示其基因功能,辅助玉米分子育种。
猜你喜欢
作物学报(2022年1期)2022-11-05
河北科技师范学院学报(2022年2期)2022-08-26
农业工程学报(2022年7期)2022-07-09
河北果树(2021年4期)2021-12-02
今日农业(2021年14期)2021-10-14
天津医科大学学报(2021年1期)2021-01-26
当代陕西(2020年14期)2021-01-08
医药前沿(2020年20期)2020-11-10
今日农业(2019年11期)2019-08-15
当代陕西(2019年12期)2019-07-12
