
晚冬早春阶段增温对冬小麦光合性能及旗叶衰老的调控作用
2019-08-13 08:00:56闫鹏孙小诺杜雄高震边大红
中国农业科学 2019年15期
闫鹏,孙小诺,杜雄,高震,边大红
晚冬早春阶段增温对冬小麦光合性能及旗叶衰老的调控作用
闫鹏1,孙小诺2,杜雄1,高震1,边大红1
(1河北农业大学农学院/河北省作物生长调控重点实验室,河北保定 071000;2河北省保定市气象局,河北保定 071000)
【】针对华北平原北部冬小麦生长发育所需的适宜温度与环境实际温度间的矛盾,通过晚冬早春阶段性增温,研究拔节前增温及拔节后揭除塑膜相对降温对冬小麦旗叶光合性能和衰老的调控作用,以期为华北平原冬小麦通过调控温度延缓小麦衰老和挖掘产量潜力提供理论及方法依据。【】2015—2017年连续2个生长季在河北省农林科学院深州旱作节水农业试验站进行大田试验,以“衡观35”为试验材料,晚冬早春采用搭建温室的方法,设置覆盖2层塑膜(M2E,1月25日—3月25日)、覆盖2层塑膜(M2L,2月5日—3月25日)、覆盖1层塑膜(M1,2月20日—3月25日)、常规生产对照(CK)共4个处理,以覆盖有孔和无孔塑膜以及时间的早晚和长短来调控温度和小麦生长发育时间,由此获得了时间相同的小麦生长发育进程不同、发育进程相同而所处日期和温度不同的结果。试验过程中记录每个处理各生育期起始时间,并测定光合特性、衰老相关酶活性、产量、水分利用效率等指标。【】早覆盖的M2E处理的冬小麦比对照开花提前8 d,成熟提前3—4 d;灌浆期旗叶叶绿素相对含量SPAD提高17.3%、净光合速率提高30.8%、超氧化物歧化酶(SOD)活性提高23.7%、过氧化氢酶(CAT)活性提高27.2%、过氧化物酶(POD)活性提高19.4%、丙二醛(MDA)含量降低23.8%;开花时的旗叶面积增大27%,收获时籽粒产量提高22.8%,水分利用效率提高16.9%。随着增温时间的推迟和覆盖时间的缩短,上述指标与对照差异越来越小,以至于M1处理上述指标与CK均显著不差异。【】晚冬早春阶段增温既维持了旗叶较高的光合速率,又显著延缓了旗叶衰老,进而为灌浆提供了物质基础;既增加了小麦穗粒数,又延长了小麦灌浆时间,从而获得了高产,获得高产的同时虽然增加了耗水量但促进了水分的高效利用。晚冬早春阶段增温是华北平原北部冬小麦有效平衡热量资源供应与生长发育所需的方法,既可减轻晚冬早春冻害对小麦返青及拔节的影响,又可规避小麦生育后期干热风带来的危害。
冬小麦;阶段增温;光合特性;旗叶衰老;籽粒产量
0 引言
【研究意义】华北平原北部是我国重要的冬小麦种植区,但由于春季升温快使小麦穗发育时间缩短,导致穗粒数减少[1];花后高温且生育后期易遭受干热风危害,使灌浆时间缩短,导致粒重降低[2],河北单位面积小麦产量显著低于河南、山东两省。造成这一结果的根本原因在于温度变化同小麦的生长发育需求存在着显著的“不对称性”[3],即冬季低温致使小麦休眠且返青较晚,而春季升温快、温度高致使小花育性降低、灌浆时间短且快速衰老。因此,通过晚冬早春增温促使小麦提早生长发育,当外界温度升高后再相对降温,从而使环境温度与冬小麦生长发育相适应,这对挖掘小麦产量潜力及缓解区域气候原因对其生长发育消极影响具有十分重要的意义。【前人研究进展】冬小麦返青后的温度变化显著影响着小麦的生长发育和衰老进程,进而影响其干物质积累和光合产物向籽粒的转运[4]。有研究表明,拔节前低温会使小麦单株穗粒数显著降低[5],并且植株MDA含量显著增加,进而加快了小麦衰老过程[6],拔节前低温胁迫还会导致生育后期旗叶叶绿素含量、光合速率、蒸腾速率和叶片气孔导度等指标的显著下降[7]。而冬季全天或白天增温可显著增加小麦穗数和千粒重[8],春季夜间增温致使冬小麦物候期提前、生育期缩短、灌浆过程缩短而减产[9]。研究表明,冬小麦花后高温造成旗叶光合速率和干物质积累量降低[10-11],源库之间物质分配比例失衡[12],造成产量与品质协同降低的负面效应[13-14]。原因在于花后高温降低了旗叶可溶性蛋白含量及SOD、POD、CAT的活性,MDA含量升高,致使旗叶早衰[15]。因此探索如何避免小麦在拔节前低温和花后高温的胁迫的新方法成了进一步提高产量的关键所在。【本研究切入点】目前,关于温度对小麦产量形成及衰老效应的研究大部分集中在冬季、花后和全生育期增温等方面,而鲜有晚冬早春阶段性升温对旗叶光合特性及衰老的研究,其对小麦产量形成的生理原因也有待进一步探索。【拟解决的关键问题】本研究在大田环境下,在冬小麦完成春化后的晚冬早春,通过田间搭建密闭钢构棚室实现升温以提前小麦的生长发育进程,拔节后揭除塑膜相对降低早发小麦生长的环境温度,分析晚冬早春阶段性增温对冬小麦旗叶光合特性及其衰老特性的影响,以期为华北北部挖掘冬小麦产量潜力及通过调控温度延缓小麦衰老提供理论和方法依据。
1 材料与方法
1.1 试验地概况
田间试验于2015—2017年连续2个生长季在河北省农林科学院深州旱作节水农业试验站(37°54′12.5″N,115°42′10.94″E进行。深州试验站年平均气温 12.7℃,试验地土壤类型为壤质潮土,耕层土壤含有机质12.53 g∙kg-1、碱解氮65.8 mg∙kg-1、速效磷15.3 mg∙kg-1、速效钾121.9 mg∙kg-1。其生态与生产条件代表华北北部平原。
1.2 试验设计
1.2.1 田间管理 采用“衡观35”作为试验材料,在每年的10月8日播种。小麦播种前,前茬玉米秸秆全部粉碎翻压还田,按135 kg N ∙hm-2、135 kg P2O5∙hm-2、180 kg K2O∙hm-2施底肥,拔节期随灌水追氮(N)135 kg∙hm-2。采用精量小区播种机进行播种,播种量187.5 kg∙hm-2,15 cm等行距种植,其他管理措施同当地高产田。
1.2.2 试验处理 试验设4个处理,分别为M2E处理(1月25日覆盖2层塑膜,其中内层有孔、外层无孔,3月5日揭除外层无孔膜,3月25日揭除内层有孔膜);M2L处理(2月5日覆盖2层塑膜,其中内层有孔、外层无孔,3月5日揭除外层无孔膜,3月25日揭除内层有孔膜);M1处理(2月20日覆盖1层透明孔膜,3月25日揭除)和CK(常规对照)。每处理3次重复,共12个小区,随机区组排列,小区面积为16 m2(4 m×4 m),小区之间留0.8 m观察道。小麦春化后,在田间为每个升度处理搭建高30 cm的钢构棚室(4 m×4 m),棚室上面覆盖透明塑膜,以覆盖有孔、无孔塑膜以及覆盖时间的早晚和长短来调控棚室内温度和小麦返青起始时间,塑膜覆盖后压实四周,避免透风影响棚内温度,随着外界气温的回升,通过减少塑膜层数来调节棚内温度。由此获得了时间相同的小麦生长发育进程不同、生长进程相同而所处日期和温度不同的结果。
塑膜厚度均为0.03 mm且白色透明,孔膜由电钻钻取而成(孔口直径1 cm,孔口间距为2 cm左右)。棚室搭建后,露地和棚室内用MicroLite5008型U盘式温度记录仪测定温度,温度记录仪放置于小百叶箱内,每小时自动记录1次温度,以此计算各处理增温阶段日均温度(图1)和各处理开花前8 d至花后32 d日均温度(图2)。
M2E、M2L和M1等3个处理在覆盖升温阶段2016年分别增加积温274.3℃、204.4℃和15.3℃,2017年分别增加积温189.9℃、140.6℃和20.3℃。
开花前8 d至花后32 d积温M2E、M2L和M1比CK的积温减少量2016分别为62.07℃、46.64℃,和7.13℃,2017年分别为76.1℃、53.79℃和无降低。
1.3 测定项目及方法
1.3.1 生育时期调查 观察并记录各处理各生育时期出现的日期,扬花期标记各小区在开花时间相同的样株200株。当籽粒含水量降至18%时为成熟,此时小麦整株枯黄,籽粒黄色变硬。
1.3.2 旗叶叶面积的测定 于开花期每小区测量旗叶20片,3次重复,旗叶面积=叶长×最大叶宽×0.83[16]。
1.3.3 旗叶光合特性的测定 采用美国CID公司生产的CL−340手持式光合作用测量系统,在开花时和花后第8、16、24、32天的早上10:00−11:00测量旗叶净光合速率(n)、气孔导度(s)、蒸腾速度(r)、细胞间CO2浓度(i)。使用日本产SPAD−502叶绿素仪测定叶绿素相对含量。
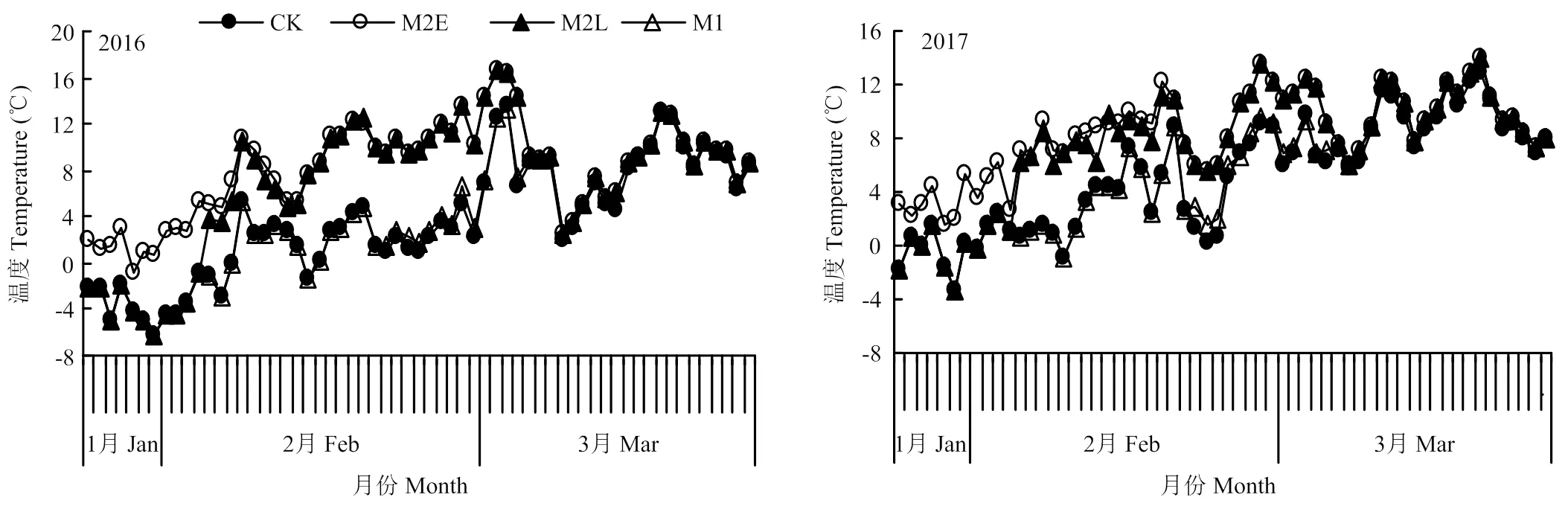
图1 2016和2017年处理增温阶段日均温度

图2 2016和2017年处理开花前8 d至花后32 d日均温度
1.3.4 旗叶生理生化指标的测定 开花至成熟 每8 d取标记植株的旗叶,用液氮速冻后置于-80℃超低温冰箱中保存。将0.5 g剪碎的叶片鲜样放入预冷的研钵中,加入1 ml磷酸缓冲液,在冰浴下研磨成浆,加缓冲液定容至终体积为5 ml,4℃条件下10 000 r/min离心20 min,取上清液用于酶活性测定。采用氮蓝四哇(NBT)光还原法[17]测定SOD活性,Sigma[18]法测定POD活性,赵士杰等[19]改进的方法测定CAT活性及MDA含量。
1.3.5 籽粒产量及其构成因素的测定 成熟期每小区连续收取小麦20株,测定穗粒数。选取中间未被取样破坏的两行,在1 m的区域内计数穗数并折合单位面积穗数,3次重复,而后进行收获脱粒并进行称重和测定籽粒含水量,按14%的含水量折算单位面积产量。
1.3.6 生育期耗水量测定及水分利用效率计算 在播种时和收获后用土钻每10 cm为一层钻取0—200 cm土层土样,采用烘干法测定土壤含水量。小麦全生育期内用自动气象站跟踪监测降水量,2015—2016和2016—2017生长季降水量分别为62.3 mm和99.8 mm。依据下列公式计算小麦田间耗水量和水分利用效率[2]:
土壤贮水量(mm)= 土层厚度(cm)×土壤容重(g∙cm-3)×土壤重量含水量×10;
田间耗水量(mm)= 播种时土壤贮水量(mm)+降水量(mm)+灌水量(mm)-收获时土壤贮水量(mm);
水分利用效率(kg∙hm-2∙mm-1)= 籽粒产量(kg∙hm-2)/田间耗水量(mm)。
1.3.7 花后每8 d积温和光合特性及酶活性数量关系的建立 以M2E、M2L、M1、CK花后每8 d积温为横坐标,这8 d中光合特性及酶活性的平均值为纵坐标建立散点图,然后添加花后8 d积温与旗叶光合特性及酶活性的二阶多项式趋势线,进而拟合日均温与旗叶光合特性及酶活性的数量关系。
1.4 数据处理
采用Excel 2010软件进行数据统计分析和作图,利用SPSS软件进行不同处理的差异显著性检验(LSD法)。由于2015—2017年度旗叶光合特性及酶活性变化趋势相似,所以图中数据均为2年数据平均值±标准差。
2 结果
2.1 冬小麦开花期旗叶面积和开花、成熟时间
由表1可以看出,晚冬早春积温的增加量越多,小麦开花日期提前就越明显,且开花时的旗叶面积就越大;棚室揭除后相对降低了早发小麦所处环境的温度,积温的降低量越多,小麦灌浆时间的延长效果就越显著。2个年度内M2E处理比CK开花提前了8—9 d,旗叶面积增大了24.6%—25.5%,灌浆时间延长了5—6 d;M2L处理开花时旗叶面积比CK提高了14.6%—20.3%,开花提前了6—7 d,灌浆时间延长了4 d;随着增温时间的推迟和覆盖时间的缩短,旗叶面积及开花和成熟日期与CK的差异越来越小,以至于M1处理与CK不具有显著差异。这表明晚冬早春合理的阶段增温能使小麦显著增大旗叶面积、提早开花日期、延长灌浆时间、获得更多光合产物,为小麦高产准备了条件。

表1 小麦开花期旗叶面积和开花及成熟时间
同列不同字母表示处理间差异显著(<0.05)。下同
Different letters in the same column mean significant differences among different treatments at 0.05 level. The same as below
2.2 冬小麦旗叶叶绿素相对含量及光合特性
2.2.1 旗叶叶绿素相对含量SPAD 由图3可以看出,各处理的旗叶叶绿素相对含量均随生育进程的推进而下降,同一时间不同处理差异显著。增温较早且时间较长的M2E和M2L处理在观测周期内旗叶叶绿素相对含量全部显著高于M1处理和CK,M2E和M2L处理的叶绿素相对含量比CK在花后16 d分别提高了19.0%和13.3%;花后24 d分别提高了15.6%和12.2%;花后32 d分别提高了56.4%和49.9%。而升温较晚和时间较短的M1处理与CK之间则差异不显著。这表明晚冬早春阶段增温后促进生育进程的效果创造了早发小麦灌浆期间相对低温的环境,有效降低了旗叶叶绿素的衰减速度。
2.2.2 旗叶光合特性 由图4可以看出,各处理旗叶净光合速率(n)和叶片气孔导度(s)均随生育进程而下降,蒸腾速率(r)短暂上升之后随即下降,细胞间二氧化碳浓度(i)一直上升,且处理间存在显著差异。花后24 d内,M2E处理净光合速率比CK提高了16.0%—31.5%,气孔导度增加了10.9%—20.8%,蒸腾速度增加了12.6%—22.7%,细胞间CO2浓度降低了11.4%—19.7%。随着处理时间的推迟及增温时间的缩短,n、s、r的增加幅度及i的下降幅度显著变小,以致花后n及其他3个指标在M1与CK间无显著差异。由图5可知,花后积温与旗叶n、s呈显著负相关,与i呈显著正相关,当花后日均温为25.9—26.0℃时,净光合速率迅速降低并达到最小值。由于CK和M1处理所处的环境温度较高,因此造成旗叶净光合速率显著降低,而处于相同生育期的M2E和M2L处理所处的环境温度相对较低,进而旗叶净光合速率得以显著提高。由此可见,晚冬早春阶段增温显著提高了旗叶光合性能,为提高灌浆速度和进而增产提供了物质基础。
2.3 旗叶抗氧化酶活性及与花后温度的关系
由图6可以看出,各处理旗叶SOD、POD、CAT活性均随着生育进程的推移先上升后下降,且花后第8天达到最高,活性指标及变幅在处理间存在显著差异。花后24 d 内,M2E和M2L处理的抗氧化酶活性均显著高于CK,因增温时间的推迟,M2L处理提高旗叶抗氧化酶活性的效果较M2E有所下降,但两者无显著差异。M2E处理与CK相比,SOD活性平均提高了15.0%、POD活性平均提高了14.2%、CAT活性平均提高了17%;M2L处理与CK相比的结果相似,只是变化幅度略有下降,SOD、POD的活性平均提高了13.1%和9.1%,CAT活性则提高了12%。随着增温时间的继续推迟及持续时间的缩短,M1和CK不具有显著差异。花后24—32 d,由于M1和CK所处的温度较高,导致抗氧化酶活性显著降低,旗叶迅速衰老。由图7可知,旗叶抗氧化酶活性与花后积温呈显著负相关,当花后日均温分别达到33.0℃、25.6℃、27.7℃时,会导致SOD、POD与CAT的活性迅速降低并达到最小值。晚冬早春阶段性增温提前了小麦的生长发育进程,移除棚室后对于早发的小麦又相对降低了生长环境的温度,因此显著提高了旗叶抗氧化酶的活性,从而延缓了小麦的衰老。

图3 开花后不同处理的旗叶叶绿素相对含量SPAD值
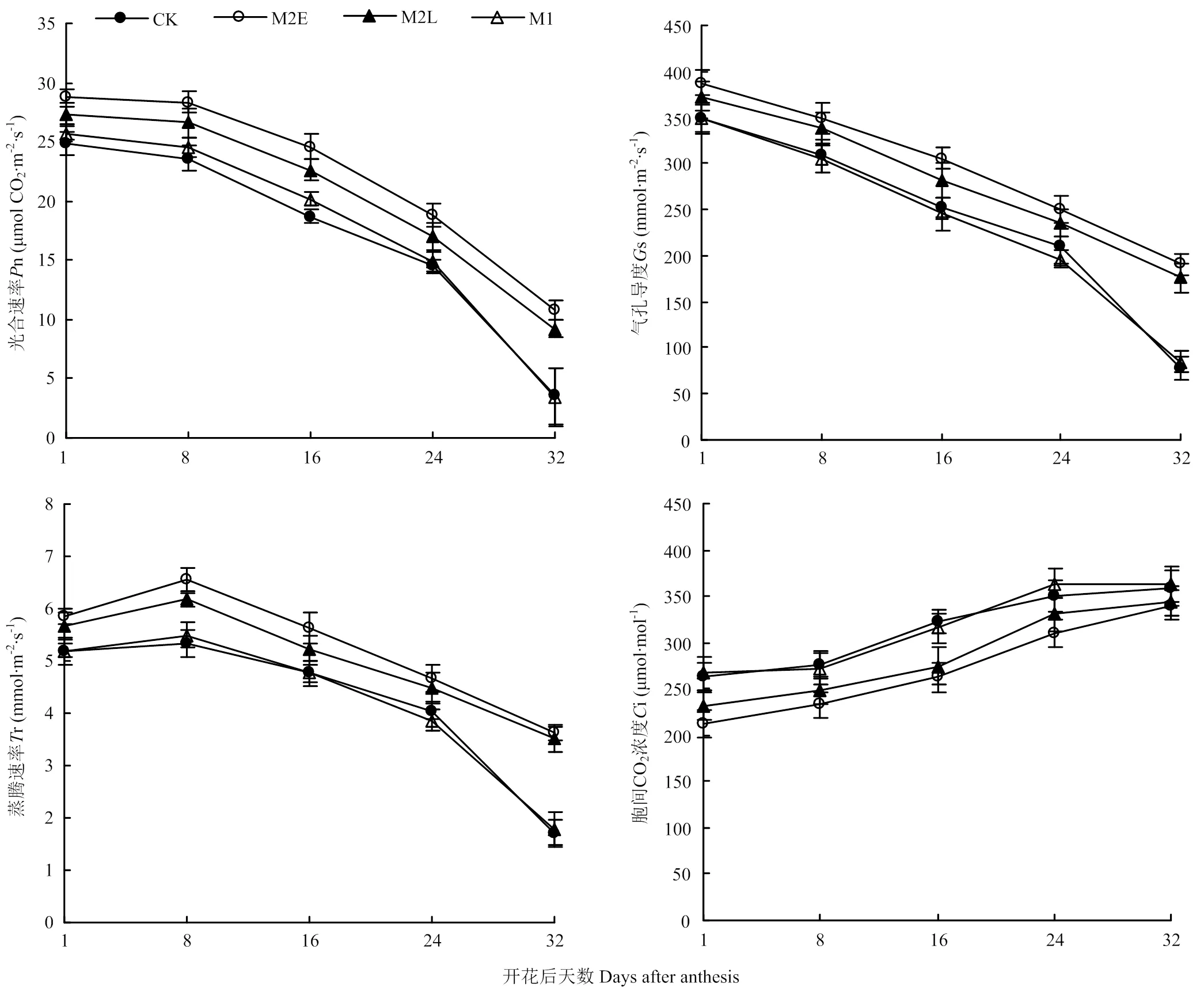
图4 开花后不同处理的旗叶光合特性变化
2.4 旗叶膜脂过氧化程度及与花后温度的关系
由图8可以看出,各处理的旗叶MDA含量随着生育进程均呈升高趋势,花后1—8 d增加缓慢,花后8—32 d增加较快,这可能与作物本身的衰老机制有关,衰老时作物机体各项修复机能衰退。花后8 d内,处理间的MDA含量差异不显著;花后8以后,处理间的MDA含量差异显著,其中M2E和M2L处理的MDA含量均显著低于CK和M1处理,M2E和M2L的MDA含量比CK分别降低了20.2%和14.6%。由图9可知,旗叶MDA含量与花后积温呈显著正相关关系,较高的花后温度造成MDA含量迅速升高,当花后日均温达到28.2℃时,MDA含量达到最大值,造成旗叶快速衰老。本研究采用的晚冬早春阶段性升温和揭膜后相对降低了早发小麦花后温度的方法,有效降低了旗叶MDA的含量,延长了旗叶功能期。

图5 开花后每8天积温和光合特性关系
Fig. 5 Relationship between accumulated temperature and photosynthetic characteristics every 8 days after flowering

图6 开花后不同处理的旗叶抗氧化酶活性变化

图7 花后每8天积温和抗氧化酶活性关系
Fig. 7 Relationship between accumulated temperature each 8-day after anthesis and antioxidant enzyme activity

图8 开花后不同处理的旗叶MDA含量
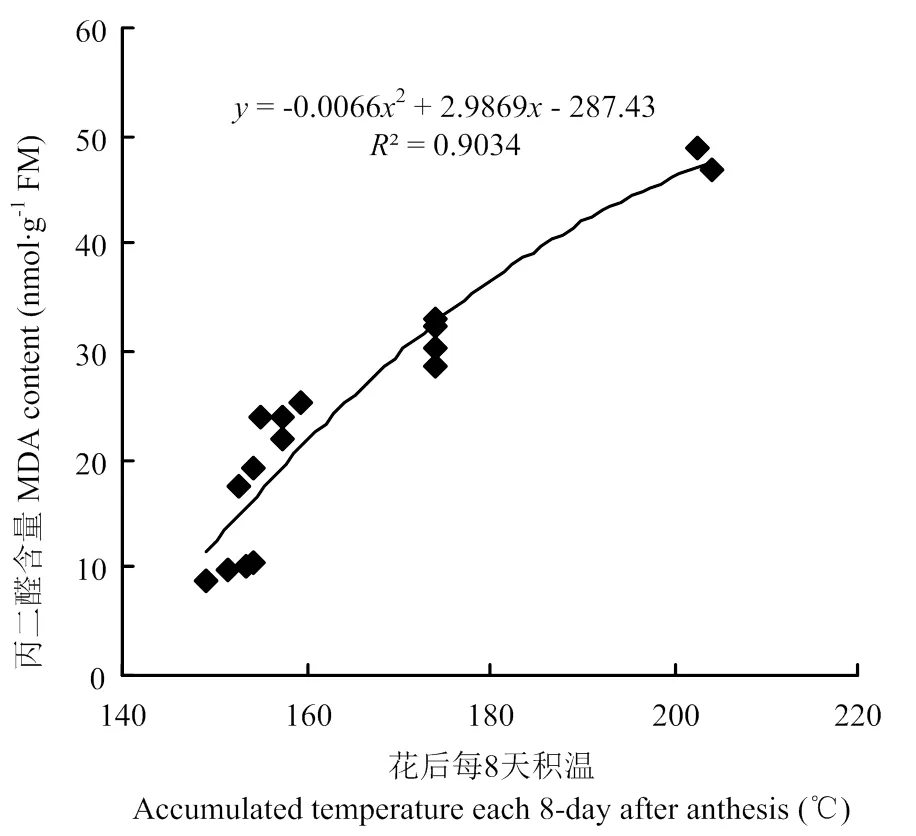
图9 花后每8天积温和MDA含量关系
2.5 籽粒产量及水分利用效率
由表2可以看出,2个试验年份M2E和M2L处理的产量及产量构成因素均高于M1处理和CK,其中穗粒数、千粒重和产量差异显著。对于产量最高的M2E处理,与CK相比2016年增产1 723.1 kg∙hm-2,增幅为20.5%;2017年增产1 857.6 kg∙hm-2,增幅为22.8%。2个试验年份产量构成因素对产量贡献率的顺序均为千粒重>穗粒数>单位面积穗数。这表明晚冬早春阶段增温显著提高产量的主要原因是增加了千粒重和穗粒数,而且升温时间越早且时间跨度越长,提高产量的效果就越显著。
晚冬早春阶段增温显著增产的同时,也相应增加了小麦的田间耗水量,但是小麦增产的速度要快于耗水的增加,因而显著提高了小麦水分利用效率。对于产量最高的M2E处理,与CK相比,2016年增产20.5%,耗水量增加4.7%,水分利用效率提高15.2%;2017年增产22.8%,耗水量增加5.4%,水分利用效率提高16.9%。这表明晚冬早春阶段增温在显著增产的同时,也显著促进了WUE的提高。

表2 不同处理的冬小麦产量和水分利用效率
3 讨论
小麦籽粒产量的形成主要来自旗叶的光合同化产物[20],但灌浆期高温会损伤旗叶光合器,并降低叶绿素含量,进而导致光合速率显著下降[21-22],同时高温也会造成光合产物输出加快,同化效率降低,进而导致产量和品质下降[23]。本研究中,在大田条件下小麦春化后的晚冬早春阶段性增温提早了小麦返青及随后的生长发育进程,进而使小麦开花提前且旗叶面积显著增大;揭除塑膜后相对降低了早发小麦的环境温度,又减慢了叶绿素分解或转移速度,显著提高了小麦花后旗叶的光合速率,为其灌浆提供了更多的光合产物。陈根云等[24]研究认为,光合速率与胞间CO2浓度呈负相关,说明光合速率随光强增加而增高主要是叶肉细胞的光合活性增大的结果,而不是i降低的结果。本研究中,花后温度与旗叶n、s呈显著负相关,与i呈显著正相关,即随着花后积温的降低,n和s上升,i则明显下降,这表明旗叶n的上升主要源于非气孔因素,推测可能与光和合系统损伤程度及叶绿素衰减程度有关。
正常条件下,植物细胞内的活性氧(ROS)清除系统会将过量的ROS清除以维持细胞内氧化还原状态的平衡。但高温胁迫会打破这种平衡,使细胞内ROS显著积累,并破坏ROS清除系统[25],进而导致膜脂过氧化程度加剧,质膜相对透性增加,旗叶衰老加快[26−27],光合能力下降,有机物积累受阻,最终导致粒重降低[28]。本研究中,晚冬早春阶段性增温提前小麦生长发育和揭膜后创造的相对低温,显著提高了花后各生育期旗叶保护酶SOD、POD、CAT的活性,降低了MDA含量,缓解了活性氧或其他过氧化物自由基对细胞膜系统的伤害,使细胞维持在可自身修复的调和状态,延缓了旗叶衰老,但随着增温时间的推迟和持续时间的缩短,旗叶衰老延缓的效果逐渐减弱。同时本研究也发现,花后温度与旗叶抗氧化酶活性呈显著负相关,与MDA含量呈显著正相关,即随着花后积温的减少,旗叶抗氧化酶活性提高,膜脂过氧化程度降低,从而衰老速度显著变缓。
前人研究表明,开花前6—8 d高温处理小麦,会使小花育性下降60%左右,进而导致穗粒数显著降低,而且高温对小花育性的危害呈现累加效应[29];Wardlaw等[30]研究表明开花期高温显著降低穗粒数,主要原因在于小花畸形、花粉败育和组织脱水。小麦籽粒灌浆期的长短也主要取决于温度,温度升高会引起叶片特别是旗叶的早衰[31-32],进而加快灌浆进程,缩短灌浆时间,造成粒重下降[33]。本研究中,2015—2017两个试验年度处理冬小麦开花至成熟平均温度分别为22.3℃和24.1℃,持续天数分别为35 d和34 d,日均温大于25.0℃的天数分别为1 d和11 d;而增产效果最好的M2E处理开花至成熟平均温度分别为21.0℃和22.3℃,持续天数都为40 d,日均温大于25.0℃的天数分别为1 d和8 d。由于增温处理的小麦发育进程提前,塑膜揭除后相对降低了早发小麦生长的温度,从而显著提高了有效穗粒数,延缓了旗叶衰老,进而延长了灌浆时间,增加了粒重,并且使成熟期提前,有效减轻了小麦在生育后期受干热风的影响程度。
研究表明,人工增加越冬前活动积温大于60℃时,幼穗发育进程明显加快,冬小麦物候期提前,使小麦整个发育期缩短,容易遭受春季低温危害,造成小花败育甚至死亡[3]。本研究中,2个试验年度常规对照冬小麦拔节至抽穗平均温度分别为17.0℃和17.3℃,持续天数分别为31 d和34 d,无冷害发生;而增产效果最好的M2E处理拔节至抽穗平均温度分别为14.3℃和14.0℃,持续天数分别为40 d和42 d,也无冷害发生。在晚冬早春温度较低时通过搭建温室增温促使小麦提早发育,当环境温度升高后揭膜降温,保持棚室内的温度不至于升高太快,在3月底温度稳定后才全部揭除塑膜,这可避免小麦因早发遭遇冻害的危险,由于揭膜后相对降低了拔节至抽穗的温度,从而延缓了穗分化,增加了穗分化时间,进而增加了穗粒数[34]。在全球气候变暖背景下,周林等[35]通过SUCROS模型研究发现,黄淮海平原秋、冬季或全生育期适度增温,冬小麦会出现全生育期缩短,但开花至成熟期延长的结果,进而有利于籽粒干物质积累;而春季增温使得冬小麦从播种至开花、开花至成熟的日数均缩短,致使小麦籽粒产量下降。本研究中,通过晚冬早春阶段性增温,利用了小麦原来因低温而不能利用的时间,大幅延长了返青到成熟期的周期跨度,并且延长了旗叶光合功能期,因而获得了高产。
4 结论
晚冬早春(1月底至3月底)阶段性增温使小麦开花提早了8 d,移除温室后的相对降温,有效延长了后续各生育阶段的持续时间,显著提高了小麦的穗粒数;而且旗叶面积比对照增加了27%,并显著提高了旗叶超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、过氧化物酶的活性(POD),有效缓解了旗叶的衰老。灌浆期旗叶叶绿素相对含量比对照增加13.3%,净光合速率提高了24.9%,从而获得了更高的粒重。晚冬早春阶段性增温合理调配了热量资源同小麦生长发育需求,获得了籽粒产量提高22.8%,水分利用效率提高16.9%的显著效果。
[1] 谭凯炎, 邬定荣, 赵花荣. 气候变暖背景下华北平原冬小麦生育期温度条件变化趋势分析. 中国农业气象, 2017, 38(6): 333-341.
Tan K Y, Wu D R, Zhao H R. Trend analysis of temperature conditions over different growth periods of winter wheat under climate warming in North China Plain., 2017, 38(6): 333-341. (in Chinese)
[2] 赵俊芳, 赵艳霞, 郭建平, 穆佳. 基于干热风危害指数的黄淮海地区冬小麦干热风灾损评估. 生态学报, 2015, 35(16): 5287-5293.
Zhao JF, Zhao YX, Guo JP, Mu J. Assessment of the yield loss of winter wheat caused by dry-hot wind in Huanghuaihai Plain based on the hazard index of dry-hot wind., 2015, 35(16): 5287-5293. (in Chinese)
[3] 杜雄, 张维宏, 张永升, 曹彩云, 李科江. 晚冬早春田间阶段性覆膜增温促进冬小麦产量提高. 作物学报, 2016, 42(10): 1530-1540.
Du X, Zhang WH, Zhang YS, Cao C Y, Li KJ. Artificial warming from late winter to early spring by phased plastic mulching increases grain yield of winter wheat., 2016, 42(10): 1530-1540. (in Chinese)
[4] 李向东, 张德奇, 王汉芳, 邵运辉, 方保停, 吕风荣, 岳俊芹, 马富举. 越冬前增温对小麦生长发育和产量的影响. 应用生态学报, 2015, 26(3): 839-846.
Li X D, Zhang D Q, Wang H F, Shao Y H, Fang B T, LÜ F R, Yue J Q, Ma F J. Impact of temperature increment before the over-wintering period on growth and development and grain yield of winter wheat., 2015, 26(3): 839-846. (in Chinese)
[5] 张军, 鲁敏, 孙树贵, 杜万里, 刘洋, 武军, 陈新宏. 拔节期低温胁迫对小麦生理生化特性和产量的影响. 西北农业学报, 2014, 23(2): 73-79.
Zhang J, Lu M, Sun S G, Du W L, LiuY, Wu J, Chen X H.Changes of physiological and biochemical parameters and grain yield at jointing stage of wheat under low temperature stress., 2014, 23(2): 73-79. (in Chinese)
[6] Xu J, Li Y, Sun J, Du L, Zhang Y, Yu Q, Liu X. Comparative physiological and proteomic response to abrupt low temperature stress between two winter wheat cultivars differing in low temperature tolerance., 2013, 15: 292-303.
[7] Kratsh H A, Wise R R. The ultrastructure of chilling stress.2000, 23: 337-350.
[8] 石姣姣, 江晓东, 史宏斌, 陈元珺, 袁久坤, 姜鸣. 冬季增温对田间小麦光合作用及产量的影响. 麦类作物学报, 2015, 35(3): 352-356.
Shi J J, Jiang X D, Shi H B, Chen Y J, Yuan J K, Jiang M. Effect of winter warming treatments on photosynthesis and yield of wheat., 2015, 35(3): 352-356. (in Chinese)
[9] 房世波, 谭凯炎, 任三学. 夜间增温对冬小麦生长和产量影响的实验研究. 中国农业科学, 2010, 43(15): 3251-3258.
Fang S B, Tan K Y, Ren S X. Winter wheat yields decline with spring higher night temperature by controlled experiments., 2010, 43(15): 3251-3258. (in Chinese)
[10] 陶龙兴, 谈惠娟, 王熹, 曹立勇, 宋建, 程式华. 开花和灌浆初期高温胁迫对国稻6号结实的生理影响. 作物学报, 2009, 35(1): 110-117.
Tao L X, Tan H J, Wang X, Cao L Y, Song J, Cheng S H. Physiological effects of high temperature stress on grain-setting for guodao 6 during flowering and filling stage., 2009, 35(1): 110-117. (in Chinese)
[11] Farooq M, Bramley H, Palta J A, Siddique K H M. Heat stress in wheat during reproductive and grain-filling phases., 2011, 30(6): 491-507.
[12] 傅晓艺, 何明琦, 史占良, 赵彦坤, 王秀堂, 郭进考. 灌浆期高温胁迫对小麦灌浆特性和品质的影响. 麦类作物学报, 2015, 35(6): 867-872.
Fu X Y, He M Q, Shi Z L, Zhao Y K, Wang X T, Guo J K. Effect of high temperature stress during grain-filling period on wheat grain-filling characteristics and quality., 2015, 35(6): 867-872. (in Chinese)
[13] 谭凯炎, 杨晓光, 任三学, 房世波. 高温胁迫对华北地区冬小麦灌浆及产量的影响. 生态学报, 2015, 35(19): 6355-6361.
Tan K Y, Yang X G, Ren S X, Fang S B. Impact of high temperature stress at the grain-filling stage on winter wheat yield., 2015, 35(19): 6355-6361. (in Chinese)
[14] NARAYANAN S, PRASAD P V V, FRITZ A K, BOYLE D L, GILL B S. Impact of high night-time and high daytime temperature stress on winter wheat., 2015, 201(3): 206-218.
[15] 姜春明, 尹燕枰, 刘霞, 王振林. 不同耐热性小麦品种旗叶膜脂过氧化和保护酶活性对花后高温胁迫的响应. 作物学报, 2007, 33(1): 143-148.
Jiang C M, Yi Y P, Liu X, Wang ZL. Response of flag leaf lipid peroxidation and protective enzyme activity of wheat cultivars with different heat tolerance to high temperature stress after anthesis., 2007, 33(1): 143-148. (in Chinese)
[16] 吕学莲, 白海波, 董建力. 春小麦旗叶大小相关性状的QTL定位分析. 麦类作物学报, 2016, 36(12): 1587-1593.
LÜ X L, Bai H B, Dong J L. QTL mapping for size traits of flag leaf in spring wheat., 2016, 36(12): 1587-1593. (in Chinese)
[17] 刘家尧, 刘新. 植物生理学实验教程. 北京: 高等教育出版社, 2010: 61-62.
Liu JY, Liu X.Beijing: Higher Education Press, 2010: 61-62. (in Chinese)
[18] Fakhari F, Sadeghi H. The effect of pod elimination on water stress in relation to antioxidant enzymes activity and proline in three annual medics species., 2016, 19(1): 109-115.
[19] 赵世杰, 苍晶. 植物生理学实验指导. 北京: 中国农业出版社, 2016: 234-235.
Zhao SJ, Cang J.. Beijing: China Agriculture Press, 2016: 234-235. (in Chinese)
[20] 徐澜, 高志强, 安伟. 冬麦春播条件下旗叶光合特性、叶绿素荧光参数变化及其与产量的关系. 应用生态学报, 2016, 27(1): 133-142.
Xu L, Gao Z Q, An W. Flag leaf photosynthetic characteristics, change in chlorophyll fluorescence parameters, and their relationships with yield of winter wheat sowed in spring., 2016, 27(1): 133-142. (in Chinese)
[21] Prasad P V V, Pisipati S R, Ristic Z, Bukovnik U, Fritz A K. Impact of nighttime temperature on physiology and growth of spring wheat., 2008, 48(6): 2372-2380.
[22] Ristic Z, Bukovnik U, Prasad P V V. Correlation between heat stability of thylakoid membranes and loss of chlorophyll in winter wheat under heat stress., 2007, 47(5): 2067-2073.
[23] Feng B, Liu P, Li G, Dong S T, Wang F H, Kong L A, Zhang J W. Effect of heat stress on the photosynthetic characteristics in flag leaves at the grain-filling stage of different heat-resistant winter wheat varieties., 2014, 200: 143-155.
[24] 陈根云, 陈娟, 许大全. 关于净光合速率和胞间CO2浓度关系的思考. 植物生理学通讯, 2010, 46(1): 64-66.
Chen G Y, Chen J, Xu D Q. Thinking about the relationship between net photosynthetic rate and intercellular CO2concentration., 2010, 46(1): 64-66. (in Chinese)
[25] Narayanan S, Prasad P V V, Fritz A K, Boyle D L, Gill B S. Impact of high night-time and high daytime temperature stress on winter wheat., 2015, 201(3): 206-218.
[26] 刘萍, 郭文善, 浦汉春, 封超年, 朱新开, 彭永欣. 灌浆期高温对小麦剑叶抗氧化酶及膜脂过氧化的影响. 中国农业科学, 2005, 38(12): 2403-2407.
Liu P, Guo W S, Pu H C, Feng C N, Zhu X K, Peng Y X. Effects of high temperature during grain filling period on antioxidant enzymes and lipid peroxidation in flag leaves of wheat., 2005, 38(12): 2403-2407. (in Chinese)
[27] 张英华, 杨佑明, 曹莲, 郝杨凡, 黄菁, 李金鹏, 姚得秀, 王志敏. 灌浆期高温对小麦旗叶与非叶器官光合和抗氧化酶活性的影响. 作物学报, 2015, 41(1): 136-144.
Zhang Y H, Yang Y M, Cao L, Hao Y F, Huang J, Li J P, Yao D X, Wang ZM. Effect of high temperature on photosynthetic capability and antioxidant enzyme activity of flag leaf and non-leaf organs in wheat., 2015, 41(1): 136-144. (in Chinese)
[28] Prasad P V, Pisipati S R, Momčilović I, Ristic Z. Independent and combined effects of high temperature and drought stress during grain filling on plant yield and chloroplast EF-Tu expression in spring wheat., 2011, 197: 430-441.
[29] Prasad P V V, Djanaguiraman M. Response of floret fertility and individual grain weight of wheat to high temperature stress: sensitive stages and thresholds for temperature and duration., 2014, 41(12): 1261-1269.
[30] Wardlaw I F, Wrigley C W. Heat tolerance in temperate cereals: an overview., 1994, 21(6): 695-703.
[31] Lobell D B, Ortizmonasterio J I, Asner G P, Matson P A, Naylor R L, Falcon W P. Analysis of wheat yield and climatic trends in Mexico., 2005, 94(2): 250-256.
[32] Wang X, Cai J, Jiang D, Liu F, Dai T, Cao W. Pre-anthesis high-temperature acclimation alleviates damage to the flag leaf caused by post-anthesis heat stress in wheat., 2011, 168(6): 585-593.
[33] 宋维富, 周超, 杨雪峰, 张延滨, 宋庆杰, 张春利, 辛文利, 肖志敏, 张延明, 李集临. 灌浆期不同阶段高温胁迫对春小麦籽粒生长的影响. 麦类作物学报, 2017, 37(9): 1195-1200.
Song W F, Zhou C, Yang X F, Zhang Y B, Song Q J, Zhang C L, Xin W L, Xiao Z M, Zhang Y M, Li J L. Effect of heat stress during grain filling on grain growth of spring wheat varieties., 2017, 37(9): 1195-1200. (in Chinese)
[34] 龚绍先. 粮食作物与气象. 北京农业大学出版社, 1988: 39-58.
Gong S X.. Beijing: Beijing Agricultural University Press, 1988: 39-58. (in Chinese)
[35] 周林, 王汉杰, 朱红伟. 气候变暖对黄淮海平原冬小麦生长及产量影响的数值模拟. 解放军理工大学学报(自然科学版), 2003, 4(2): 76-82.
Zhou L, Wang H J, Zhu H W. Simulation study on the impact of climate warming on production of winter wheat in huang-Huai-Hai plain of china., 2003, 4(2): 76-82. (in Chinese)
Effects of Artificial Warming from Late-Winter to Early-Spring on Photosynthesis and Flag Leaf Senescence of Winter Wheat
YAN Peng1, SUN XiaoNuo2, DU Xiong1, GAO Zhen1, BIAN DaHong1
(1College of Agronomy,Hebei Agricultural University/Key Laboratory of Crop Growth Regulation of Hebei Province, Baoding 071000, Hebei;2Baoding Meteorological Bureau, Baoding 071000, Hebei)
【】 In view of the contradiction between the optimum temperature required for the growth and development of winter wheat and the actual temperature provided in the environment in the north of North China Plain, we attempted to artificially increase the temperature from the end of January to the end of March through plastic-film greenhouse and study the effect of temperature on photosynthesis and flag leaf senescence of winter wheat, in order to provide theoretical and methodological basis for delaying wheat senescence by regulating temperature and tapping the yield potential of winter wheat in North China Plain.【】 Field experiments were conducted at Shenzhou Dry-farming and Water-saving Agricultural Test Station of Hebei Academy of Agricultural and Forestry Sciences in two consecutive growing seasons from 2015 to 2017. Using ‘Hengguan 35’ as experimental material, greenhouses were constructed in late winter and early spring. Four treatments were set up, i.e., cover 2 layers of plastic film (M2E, January 25-March 25), 2 layers of plastic film (M2L, February 5-March 25), 1 layer of plastic film (M1, February 20-March 25), and conventional production control (CK). Temperature and wheat production and development time were regulated by covering porous and non-porous plastic film and its covering time, which resulted in different growth and development processes in the same date, and the same growth process in different dates and temperatures.During the experiment, the beginning time of each growth period was recorded, and the indexes of photosynthetic characteristics, senescence-related enzymes activity, yield, and water use efficiency were measured. 【】 The treated wheat was in a relatively higher temperature in pre-jointing stage and lower temperature after jointing, it promoted the wheat to grow earlier and prolong its developing period. The winter wheat in M2E treatment was 7-8 days earlier than the control in flowering and 3-4 days earlier in ripening; At grain filling stage, the net photosynthetic rate of flag leaf increased by 24.9%, the relative content of chlorophyll, superoxide dismutase (SOD), catalase (CAT) and peroxidase (POD) increased by 13.3%, 17%, 17% and 14.2%, respectively, while the content of malondialdehyde (MDA) decreased by 22.7%.flag leaf area increased by 27% at flowering, grain yield increased by 22.8% at harvest and water use efficiency increased by 15.2%. With the delay of warming time and the shortening of covering time, the difference of the above indexes was smaller and smaller, so that there was no significant difference between M1 treatment and CK treatment.【】 Warming in late-winter and early-spring not only significantly prolonged the filling time of wheat, but also maintained higher photosynthesis of flag leaves and delayed senescence of flag leaves, which provided more material basis for grain filling, and promoted the efficient use of water at the same time of obtaining high yield. Increasing temperature in late winter and early spring could be suitable for rational allocation of heat resources and growth and development needs of winter wheat in the north of North China Plain. At the same time, it could not only alleviate the harm of late spring cold on wheat, but also avoid the influence of dry-hot wind on wheat in later growth stage.
winter wheat; artificial warming; photosynthetic characteristics; flag leaf senescence; grain yield
2019-03-11;
2019-04-25
国家重点研发计划项目(2017YFD0300908)、河北省青年优秀拔尖人才专项基金、河北农业大学作物学科梯队建设基金(TD2016C204)
闫鹏,E-mail:402372695@qq.com。
杜雄,E-mail:duxiong2002@163.com
(责任编辑 杨鑫浩)
猜你喜欢
作物学报(2022年1期)2022-11-05 08:06:34
河北科技师范学院学报(2022年2期)2022-08-26 08:55:32
作物学报(2022年9期)2022-07-18 00:58:42
农业工程学报(2022年7期)2022-07-09 06:16:32
中国典型病例大全(2022年9期)2022-04-19 21:49:39
中国乳业(2022年1期)2022-02-22 06:48:58
干旱地区农业研究(2021年6期)2021-12-08 07:52:22
今日农业(2021年17期)2021-11-26 23:38:44
浙江农业科学(2019年11期)2019-11-09 01:37:48
今日农业(2019年11期)2019-08-15 00:56:32
