
银鲫的生殖方式
2019-08-13 07:48葛彦龙李池陶贾志英石连玉
水产学杂志 2019年4期
葛彦龙,李池陶,贾志英,石连玉
(中国水产科学研究院黑龙江水产研究所,黑龙江 哈尔滨 150070)
银鲫Carassius auratus gibelio原指区别于黑鲫和红鲫的银色鲫[1]。银鲫具有2N和3N两种类型,2N个体具有100条染色体,而3N个体约有156条染色体[2-5]。普通鲫和2N银鲫都进行正常的减数分裂和两性生殖,产生各半的雌雄后代,而3N银鲫生殖方式十分特殊。通常,3N物种不能减数分裂而不育[6],但银鲫可以产生不减数卵子[7](或减数后加倍而不减数),且受精后雌核排除雄核而维持3N倍性[8]。雌核发育的银鲫本应产生全雌后代,但后代中具有雄性。雌银鲫远缘杂交时,进行雌核发育产生5%的雄性,外源雄性基因不起作用,不能解释雄性的来源;而银鲫同系繁殖时,产生15%的雄性后代[9],其偏向雌核发育并含有两性生殖过程[10],产生更多的雄性,需要确定银鲫雌核发育过程如何加入了部分两种生殖过程。2N鲫和3N银鲫生活在同一水域,基因组十分相似[11,12],似乎发生着频繁的基因交流,需要确定其中的机理。关于银鲫的生殖细胞发生过程,一般认为银鲫卵子不减数而进行天然雌核发育,精子减数而可育,但这不符合相同核型细胞减数分裂一致的遗传原理。本文不再确定银鲫配子是否发生减数,只通过银鲫繁殖的后代组成分析其亲本和配子倍性的可能性。为叙述方便,本文将2N银鲫归为普通鲫,而银鲫特指3N银鲫。
1 银鲫中具有较多含有雄性基因的嵌合体
雌银鲫用2N鲤Cyprinus carpio精子受精后进行雌核发育,后代基本为雌性,但有一尾雌银鲫雌核发育后代主要为雄性,即3%的雌性和97%雄性。用雄性特异标记鉴定发现,银鲫亲本和雄性后代都具有雄性标记,而鲤和雌性后代都没有雄性标记[13](图1右)。这表明银鲫性别主要由基因型决定,少量雌性银鲫后代不是由性逆转产生。基因型分析表明,如果这尾银鲫亲本为由同种体细胞构成的纯合体银鲫,若配子减数,银鲫亲本为XY基因型时[14],将产生各半的X、Y雌雄配子,雌核发育需要配子加倍产生各半的XX雌性和YY超雄后代;银鲫亲本为YY基因型时,只产生Y雄配子,配子加倍只产生YY超雄后代。若配子不减数,银鲫亲本只能为XXY基因型,雌核发育产生全雄后代。二者产生半数或全部雄性后代,不能形成具有少数雌性的后代组成,因此,这尾雌银鲫亲本只能为嵌合体。雌雄配子不融合且共同发育会产生嵌合体,而极体与雌核共同发育产生的嵌合体不具有雄性基因,这样,这尾嵌合体银鲫由雌雄配子不融合产生。若配子减数,X、Y雌雄配子不融合且分别加倍产生XX/YY嵌合体,根据不融合配子分裂启动的先后能产生0%~100%的雄性后代。若配子不减数,XXX、XXY雌雄配子不融合且不加倍产生XXX/XXY嵌合体,也能产生0%~100%的雄性后代。银鲫性别主要由基因型决定,含有雄性细胞的嵌合体银鲫为雄性,极少为雌性。因此,含有雄性细胞的嵌合体雌银鲫很少见。而嵌合体雄性即便很多,也不容易被发现,因为嵌合体和非嵌合体雄性和雌性银鲫交配时都主要进行雌核发育,且两种雄性都可以产生雄配子。雄配子参与生殖时都产生余外的雄性,后代都是产生多数雌性和相对远缘杂交更多的雄性,不能区分。为了证明雄性嵌合体的存在,可以直接将雄性个体的体细胞分离后用雄性标记鉴定,比如对银鲫体细胞进行涂片原位杂交或单细胞PCR。也可将雄性逆转为雌鱼,通过其雌核发育的后代组成来分析亲本的体细胞组成。将两条雄性银鲫逆转为伪雌鱼,同鲤进行雌核发育。用雄性标记进行鉴定,银鲫亲本同雄性后代都具有雄性标记,而鲤和雌性银鲫后代都没有雄性标记。后代性别完全由基因型决定,一尾逆转鱼产生了18%的雌性后代和82%的雄性后代[15](图1中)。和上文的雌性嵌合体相同,这尾逆转鱼只能为嵌合体银鲫,因为纯合体银鲫只能产生半数或全部雄性后代。它由配子不融合产生,减数时为XX/YY嵌合体,不减数时为XXX/XXY嵌合体。另一尾逆转鱼产生了48%的雌性后代和52%的雄性后代[15](图1左)。虽然可以为同种细胞构成的非嵌合体XY个体,但也同样适用于嵌合体,只是两种细胞的比例为1∶1即可。而均等比例嵌合体比不均等比例嵌合体更多。下文介绍的远缘杂交中使用的雄银鲫也可以证明为嵌合体。随机使用的雄性都含有嵌合体,说明雄性中嵌合体的比例很大,不排除纯合体雄性的存在。

图1 银鲫伪雌鱼的雌核发育Fig.1 Gynogenesis of pseudo-female silver crucian carp
2 雄银鲫的远缘杂交
远缘杂交时,雌银鲫进行雌核发育,N雄核被排除,3N雌核单独发育而产生大量的后代[16,17]。而雄银鲫远缘杂交则十分不同,3N雄核一般不会排除N雌核,杂交也不会产生大量存活的后代。用2N雌鲤同雄银鲫杂交时,一般情况下,雌核同雄核都解凝并可以融合,但不是简单的倍性相加,而是倍性相加后排除部分染色体而产生非整倍体。原核融合后纺锤体会发生错误,部分染色体遗留在赤道板处而形成非整倍体。二极纺锤体也可能继而转变为三极或四极纺锤体而停止分裂。虽然有基本正常的高成胚率,但是胚胎发育到尾部游离前后即死亡[18]。杂交是否可以产生少量存活的个体,需要进一步验证。用2N雌红鲫和3N雄银鲫杂交,也具有正常的成胚率,同样胚胎发育到尾部游离前后大量死亡。胚胎染色体数从50~142,倍性为1~2.9N[19]。胚胎的最少染色体数为50,为母本的染色体数,说明母本很可能保留了所有的染色体。若雄核为减数的1.5N,那么原核融合的初始倍性为2.5N,染色体丢失后胚胎倍性不可能高于2.5N。因此,笔者认为两性融合的雄核为不减数的3N,原核融合后胚胎初始倍性为4N,丢失了1.1~3N的雄核染色体,可以产生了1~2.9N的不整倍体胚胎。
这些非整倍体中,如果细胞核恰巧丢失了银鲫的1个或2个完整倍性的基因组,那就是产生了3N或2N的整倍体杂交鲫,可以存活。其具有鲫的1个倍性和银鲫2个或1个倍性的基因组。杂交共获得了2尾青灰色仔鱼,他们只能为整倍体且各个倍性基因组完整才能存活,而单倍体或非整倍体的鱼均不能成活。完整基因组的整倍体有几种可能性:雌核发育的2N鲫个体、雄核发育3N银鲫个体、倍性相加后未丢失染色体的4N杂交个体、倍性相加后丢失1套银鲫整倍基因组的3N杂交个体和倍性相加后丢失2套银鲫整倍基因组的2N杂交个体。雌核发育个体表型为母本的红色,这两尾青灰色鱼只能为其他类型的整倍体鱼,但后代数量较少不能统计组成,不能简单地排除2N雌核发育个体。普通鲫同3N银鲫杂交实验与以上结果大致相同,其成胚率较高,大部分胚胎尾部游离前后死亡。但不同的是,其孵化率较高,达到5%[20],其体型介于鲫和银鲫之间[21],基本为杂交鱼,但没有具体数据,需要进一步考证。成活个体可能为倍性相加的4N杂交鲫和去除1套或2套银鲫整倍基因组的3N或2N杂交鲫。除此之外,成活个体中有5%的个体为父本体型体色的3N银鲫,为雄核发育产生,雌雄比例为9∶1[20](图2)。雄核排除了雌核独立发育,从雌雄比例推断父本只能为嵌合体,因为纯合体父本雄核发育只能产生半数或全部雄性后代。总之,雄银鲫同2N鲫杂交时,主要产生非整倍体胚胎,不能存活。少量成活个体为整倍的4N或3N和2N两性生殖后代和少量的3N雄核发育后代,可能产生2N雌核发育后代。如果存在3N和2N杂交后代,就说明2N鲫和3N银鲫可以相互转化,使二者基因组保持高度相似性。
3 银鲫同系或近缘杂交
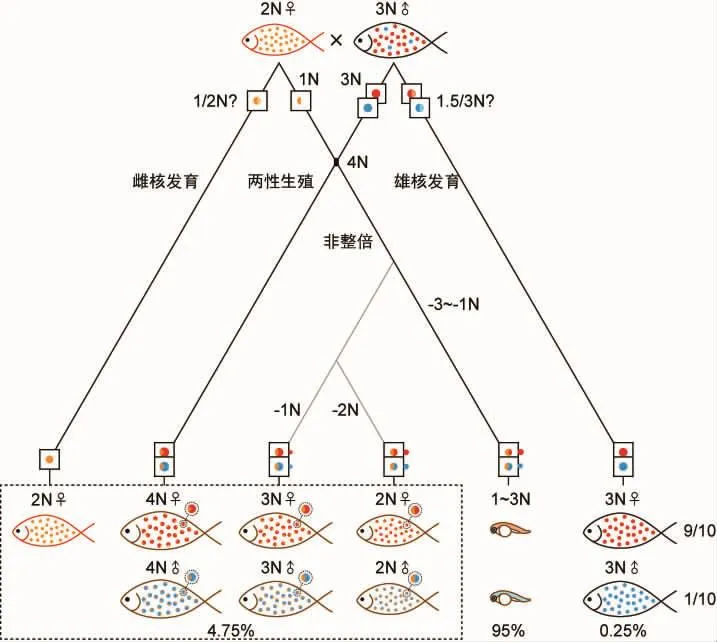
图2 雄性银鲫的远缘杂交Fig.2 Distant hybridization of male silver crucian carp
两种近缘3N银鲫杂交时,可根据体型等特征区分亲本。杂交后有9%的受精卵可发育为成鱼。这些存活个体具有三种特征:80%的个体为母本特征,15%的个体为介于父本和母本的中间型特征,5%个体为父本特征,其中40%为雄性[22](图3右)。80%的母本特征后代为雌核发育产生,基因完全来自雌亲本,雄核凝缩而被排除。5%的父本特征的后代为雄核发育产生,基因完全来自雄亲本,雌核被排除。15%的中间特征后代基因分别来自父本和母本,基因组贡献都比较大,才能形成明显区别父本和母本的后代。这些中间型后代可能为原核融合产生,需要父母本的配子都减数为1.5N,融合后恢复3N倍性。也可以由细胞嵌合产生,父母本的配子都不减数(或减数后加倍而不减数)且不融合可以产生3N嵌合体后代。笔者证明了雄性中具有较多嵌合体,雌性中也应该具有相应比例的嵌合体,只是雌性嵌合体不能用后代性别比例进行鉴别。这些嵌合体只能在两性生殖中产生,由配子不减数(或减数后加倍而不减数)且不融合产生。这样就存在配子不减数(或减数后加倍而不减数)且不融合的嵌合途径,也可能存在配子减数且融合的纯合途径,但不是必要的。雄核发育时雌雄比例为3∶2,不是1∶1,说明雄亲本为嵌合体的可能性较大。总之,近缘杂交时,亲本产生不减数雌雄配子(或减数后加倍而不减数)排除对方配子,分别进行雌核发育和雄核发育。或者雌雄不减数配子(或减数后加倍而不减数)均存活且不融合形成嵌合体个体。可能产生减数配子,在两性生殖时减数配子融合形成纯合后代。3N银鲫同系繁殖与近缘杂交相同,也应该有相同比例父母本特征的后代,只是这些父母本特征不能通过表型进行分辨,仅可以比较亲本和后代之间的基因型来区分。银鲫同系繁殖也产生80%以上的雌性个体,这说明近缘或同系繁殖产生各种类型后代的比例基本一致,至少雌核发育部分基本一致。此同系繁殖时使用的父本是嵌合体伪雌鱼的雌核发育后代[13],只能为减数时配子加倍形成的YY雄性或不减数时配子直接发育形成的XXY雄性。任何情况下,其两性生殖后代和雄核发育后代都具有雄性标记,用近缘杂交过程的比例计算,雄性比例为15%加上5%等于20%,实际统计为22%,比例基本一致(图3左)。总之,在银鲫同系和近缘杂交时,有配子不发生减数的可能性,雌核更容易单独发育产生多数的雌核发育个体。雄核可以小比例掺入(同雌核不融合)雌核发育中,形成嵌合体。雌核也可以小比例失活,使雄核进行雄核发育,配子减数也可能发生。雌核发育或雄核发育时,减数配子加倍后独立发育,两性生殖时减数配子融合发育或加倍后嵌合发育。
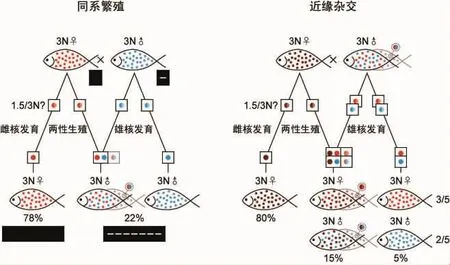
图3 银鲫的两性生殖Fig.3 Sexual reproduction of silver crucian carp
4 银鲫性别的不稳定性
雌银鲫远缘杂交时进行雌核发育而保持物种稳定,远缘雄核无法解凝最终被排除。这种远缘雌核发育产生的银鲫后代叫做异育银鲫[23]。2N鲫和3N银鲫生活在一起,外形也很难区分。银鲫主要为雌性,2N雄鲫有更多机会受精雌银鲫,产生天然的异育银鲫。这样,自然界中会存在很多天然的异育银鲫,自然采集的银鲫为异育银鲫的可能性也比较大。异育银鲫不为杂交鱼的形态,均为银鲫体型,即使生长速度有所差异,但并不影响成年鱼的最终体型,均为雌核发育的3N银鲫。异育银鲫和银鲫的基因组相同,没有本质的区别。雌核发育子代应为全雌后代,而银鲫远缘雌核发育时,具有产生全雌后代的情况[24]。但是,更多的情况却是银鲫远缘雌核发育总会产生较小比例的雄性后代。俞豪祥用鲤作父本,对银鲫进行多代雌核发育繁殖,后代中总有5%的雄性个体[23]。雄银鲫后代不被使用而被排除,但在后续繁殖时可以不断产生。用2N鲫作父本,银鲫也进行雌核发育,后代也会持续产生5%以下的雄性[25]。如果使用麦穗鱼Pseudorasbora parva、荷包红鲤Cyprinus carpio var.wuyuanensis等其他父本,后代也会产生5%以下的雄鱼。这说明远缘杂交时,父本的改变并不会改变银鲫雌核发育产生少数雄性后代的特点。而雄性不育型银鲫雄鱼精子无效,雌鱼只能进行远缘杂交繁殖,依然可以持续产生小于5%的不育雄性[1]。远缘雄性基因对银鲫无效且没有加入银鲫基因组的途径,这说明雌银鲫可以自发产生5%左右的雄性,与外缘雄性基因无关。一般认为,银鲫的性别由基因型决定,如果是完全的基因决定,雌鱼和鲤父本不具有雄性基因,雄性后代的雄性基因没有来源。因此,银鲫的性别为不完全的基因型决定,或者说银鲫的性别具有5%的不稳定性。如果雌性具有不稳定性,每一代都会有小比例的雌鱼逆转为伪雄鱼,伪雄鱼不具有雄性基因。笔者将它定义为雌性不稳定性,每一代都十分均等(图4左)。如果雄性具有不稳定性,就有一定比例的后代逆转为雌性,以在群体雌核发育中维持雄性基因,笔者将其称为雄性不稳定(图4右)。具有雄性基因的伪雌鱼,第一代根据是否减数为XXY或XY伪雌鱼,将产生全部XXY雄性后代或半数YY超雄后代。从第二代开始,XXY或YY伪雌鱼只产生雄性后代。因为银鲫的性别主要由基因型决定,这些雄性只有少数可以逆转为伪雌鱼。伪雌鱼和雌鱼共同繁殖时,伪雌鱼产生少量的伪雌鱼,而雌鱼全部产生雌鱼。这样下一代雌鱼中伪雌鱼的比例很小而很难被使用到,因此,群体繁殖时,雄性不稳定性途径只能在偶然使用到伪雌鱼时产生,并很快消失。而带有雄性基因的嵌合体银鲫更可能具有性别不稳定性[26]。但嵌合体银鲫经过一次雌核发育后会形成纯合体银鲫而终止嵌合体不稳定性途径。这样,雌银鲫远缘杂交时多代产生小比例雄性现象应该由雌性不稳定性产生,而不是由雄性不稳定性或嵌合体不稳定产生。
5 讨论
银鲫的生殖现象错综复杂,充满矛盾。早期建立的银鲫繁殖理论只是部分符合事实和遗传规律,尚需进一步完善。这些指导了银鲫生殖的研究方向,但也限制了思维,如异精生长效应。蒋一圭等[24,27]用兴国红鲤精子受精方正银鲫卵子,培育出了生长比银鲫原种快30%以上的异育银鲫。但是,雄核被排除,裂解的DNA片段被代谢掉,不能参与遗传,线粒体遗传也比较保守,所以,异精效应很难找到遗传学依据[28]。针对异精效应,沈俊宝作过多年的实验,用野鲤、镜鲤雄鱼与银鲫杂交和多次回交,各组合后代完全表现出母本性状。在相邻池塘中同银鲫进行生长对比,各组合后代的生长优势几乎相等,没有发现异精效应[9]。再如,异精雄性效应,蒋一圭认为银鲫繁殖时,雄鱼同银鲫亲缘关系越近,后代雄性比例越高;同鲤杂交,雄性后代为0,同鲫杂交,雄性后代为2.5%,与银鲫自交为15%[24,29]。这引导人们去寻找其中的遗传依据。2N鲫可能具有银鲫雄性基因,受精银鲫可产生雄性。鲤没有鲫雄性基因,受精银鲫不能产生雄性。而雄性后代都具有雄性条带,这似乎表明2N鲫的雄性基因确实进入到银鲫后代中并影响了后代性别。但上面的数据并不准确,雌银鲫同鲤等远缘杂交时,在一代或多代繁殖中都会产生0%~5%的雄鱼[30]。同2N鲫杂交时,也能产生0%~5%雄鱼。只有银鲫自交时雄性比例才会明显提高。这只能说明3N银鲫同系繁殖和雌银鲫远缘杂交具有不同的生殖途径,而和亲缘关系远近无关。雄鱼具有鲫雄性基因与否,雌鱼远缘杂交都会产生相同比例的雄性,说明银鲫雌核发育可以自发产生雄性,而不需外缘雄性基因的参与。
再如,银鲫精子减数分裂和卵子不减数的说法。卵子不减数可以天然雌核发育,而雄性可育是因为精子发生了减数。雌核和后代倍性相同,减数1.5N雄核不能置换或嵌合到不减数3N雌核中,但雄性配子确实加入到后代中而增加雄性比例。桂建芳认为,减数的1.5N雄核和不减数的3N雌核融合成为4.5N合子后排除1.5N染色体以保持3N倍性[31]。这里有两点不符合遗传规律。首先,相同的核型应该有相同的减数分裂过程,雌核和雄核都发生减数或者都不发生减数,或者都具有减数和不减数的部分。其次,不均等配子融合后会随机的丢失染色体,不会排除确定数量的染色体。比如用2N雌鲫同3N雄银鲫杂交,4N合子会不同程度丢失了染色体,形成不能存活的非整倍体和偶然存活的整倍体后代。因此,产生高比例的两性生殖后代需要银鲫雌雄配子为均等配子。卵子和精子同时减数时融合或加倍后嵌合可以产生相同倍性两性生殖后代,同时不减数时嵌合也可以产生相同倍性的两性生殖后代。一般认为,卵子没有发生减数,雌银鲫远缘杂交时,会观察到临时的3极染色体[32,33],说明雌核可能为3倍体。雄银鲫远缘杂交时,不育的非整倍胚胎中也可以观察到3极和4极纺锤体[18],这说明雄核也可能为3N而形成3极以上的纺锤体。这些胚胎中,雄核最多嵌入大于1.5N的92条染色体,也支持雄核不减数。雌核无法大量提取而不能直接证明其是否发生减数,相反,精子减数有了初步的证明。其通过2N、3N鲫精子间或体细胞间染色比对得来[34]。但是,体细胞染色程度比较彻底,而精子致密,内部可能无法染色,这导致结果不够准确。2N和3N鲫精子因压缩程度不同,比较体积确定DNA含量也可能不准确[35]。为了解释3N精子发生减数的结论,出现了3N倍性实际是6n倍性的观点[36]。6n也可归为2倍体,但普通的二倍体会产生正常性比的后代,不会出现雌核发育为主的现象,所以银鲫归为6n的说法是错误的。银鲫生殖方式的特殊性原因可能是其存在原核不融合过程,导致多数雄核失活的雌核发育和少数雄核存活的嵌合或雄核发育现象。配子是否发生减数没有确定,2N、3N银鲫发生遗传交流的方式也没有确定。2N鲫和3N银鲫生活在同一水域,从外观和基因型都很难区分,说明它们发生着频繁的遗传交流,遗传交流不是通过雌核发育或雄核发育过程,那样两个物种基因组没有融合,还相互独立。若发生频繁的基因交流,需要两种鲫杂交时配子融合且倍性可以单向或双向转换。2N鲫自交如果产生不减数配子,受精后会产生3N鲫后代,但染色体数目是150条而不是156条,且2N鲫同系繁殖中没有发现自然产生3N鲫。3N银鲫的配子无论是减数的1.5N或不减数的3N,都不能和2N鲫的N配子相加直接形成2N或3N后代。但是,3N雄银鲫同2N鲫杂交时,3N和N配子融合形成4N合子通过丢失整倍N或2N可间接得到杂合的3N或2N鲫。这样2N鲫和银鲫基因组可以整倍融合,类似于两性生殖,而且倍性可以相互转换。其他组成的非整倍体胚胎都会死亡,这样便增加了3N和2N杂合后代的比例,加大了两种倍性鲫遗传交流的几率。这就解释了两种鲫为何倍性不同又如此类似。丢失整套染色体说明鲫的基本倍性为N并具有一定的整体性,单倍性为N而不是1.5N。
猜你喜欢
数学小灵通·3-4年级(2022年10期)2022-12-31
科学与社会(2021年3期)2021-12-02
教学考试(高考生物)(2021年1期)2021-04-20
小学生学习指导(中年级)(2020年9期)2020-10-24
大自然探索(2020年3期)2020-05-03
生物学教学(2018年7期)2018-08-08
发明与创新·中学生(2017年6期)2017-06-12
方圆(2016年15期)2016-09-14
分析化学(2014年7期)2014-12-13
小学生·多元智能大王(2014年9期)2014-08-28
