
多异瓢虫对葡萄斑叶蝉的捕食作用
2019-07-29 08:31宋双姜彩鸽赵亚楠王国珍张怡
中外葡萄与葡萄酒 2019年4期
宋双,姜彩鸽,赵亚楠,王国珍,张怡*
(宁夏农林科学院植物保护研究所,宁夏银川 750002)
葡萄斑叶蝉[Erythroneura apicalis(Nawa)]也被称为葡萄二星叶蝉或葡萄二点叶蝉,属于半翅目,叶蝉科[1-2]。成、若虫多在叶背为害,一般喜在郁闭处活动取食,排出的虫粪污染了叶子和果实,造成黑褐色粪斑,影响当年以至第2年果实的质量和产量[3]。随着贺兰山东麓葡萄产区种植面积的不断扩大,葡萄斑叶蝉的发生量也在逐年增加。自2013年以来,玉泉营农场大面积发生了葡萄斑叶蝉,因消耗葡萄营养,影响叶片光合作用、枝条生长和花芽分化,造成葡萄早期落叶、树势衰退[4]。目前,葡萄斑叶蝉已经成为危害葡萄的主要害虫之一。
多异瓢虫[Hippodamia variegate(Goeze)]是一种多食性昆虫,具有很强的捕食能力,对斑叶蝉的有良好防治效果。目前,对瓢虫捕食功能的研究主要集中在多异瓢虫、异色瓢虫、七星瓢虫、小十三星瓢虫、小黑瓢虫和龟纹瓢虫等对蚜虫、木虱和粉虱的捕食功能[5-6],然而,没有关于多异瓢虫对葡萄斑叶蝉的捕食作用的研究报道。为了准确评价多异瓢虫对贺兰山东麓地区葡萄斑叶蝉的防治效果,本文针对多异瓢虫对葡萄斑叶蝉捕食功能进行了一系列研究,温度对其捕食功能和不同天敌干扰响应的影响[7],为有效利用和进行生物防治提供了科学依据。
1 试验材料和方法
1.1 供试虫源
从宁夏银川市永宁县玉泉营农场南大滩葡萄试验点采集了供试虫源葡萄斑叶蝉和多异瓢虫。试验前先将多异瓢虫成虫室内饥饿24 h再进行捕食试验;葡萄斑叶蝉成虫采用扫网法和人工捕捉将试虫放入布袋中带回室内进行试验。
1.2 多异瓢虫的捕食功能反应
葡萄斑叶蝉设置了10头、15头、20头、25头和30头5组密度处理(已通过预试验确定虫口密度),每一组处理共设置5个重复。将每1头多异瓢虫置于不同数量葡萄斑叶蝉的培养皿中,24 h后记录每个培养皿中葡萄斑叶蝉的存活量和自然死亡量。校正自然死亡率数据以确定天敌的每日捕食量,并确定捕食者效率与葡萄斑叶蝉虫口密度之间的相关性,建立捕食者功能反应模型。
1.3 多异瓢虫在不同温度下捕食功能的影响
在人工气候箱内设置5组温度处理,分别为15 ℃、20 ℃、25 ℃、30 ℃和35 ℃,每组处理共设置4个重复,每个培养皿中装入1头多异瓢虫和25头葡萄斑叶蝉,24 h后记录每个培养皿中葡萄斑叶蝉的存活量和自然死亡量。根据Holling模型[8-10],分析温度条件与捕食者行为之间的线性关系。
1.4 多异瓢虫虫口密度对葡萄斑叶蝉捕食的干扰响应
在室温(约25 ℃)下,多异瓢虫设置5组密度处理:即1头、2头、3头、4头和5头,每组处理共设置4个重复。每个培养皿放置50头葡萄斑叶蝉,24 h后检查每个培养皿中葡萄斑叶蝉的存活量和自然死亡量。根据Hassell和Verley提出的干扰响应模型[8],针对天敌密度对捕食率的影响进行分析。
1.5 多异瓢虫对叶蝉分布捕食作用的反应
将5组处理的葡萄叶片,即1片、2片、3片、4片和5片分别置于室温(约25 ℃)下的培养皿中,每1组处理共设置4个重复,每个培养皿都放置1头多异瓢虫和20头葡萄斑叶蝉,24 h后记录每个培养皿中葡萄斑叶蝉的存活量和自然死亡量,以确定天敌的每日捕食量。
2 结果与分析
2.1 多异瓢虫捕食功能反应
如表1,不同葡萄斑叶蝉虫口密度的试验结果表明:随着葡萄斑叶蝉虫口密度的增加,多异瓢虫的捕食量也逐渐增加;当葡萄斑叶蝉虫口密度增加到一定数量时,捕食量增加的速度减缓。对照Holling提出的3种不同的功能反应函数,符合其中的HollingⅡ型功能反应[8-10],可用HollingⅡ圆盘方程Na=aNtT/(1+aThNt)来拟合试验数据。
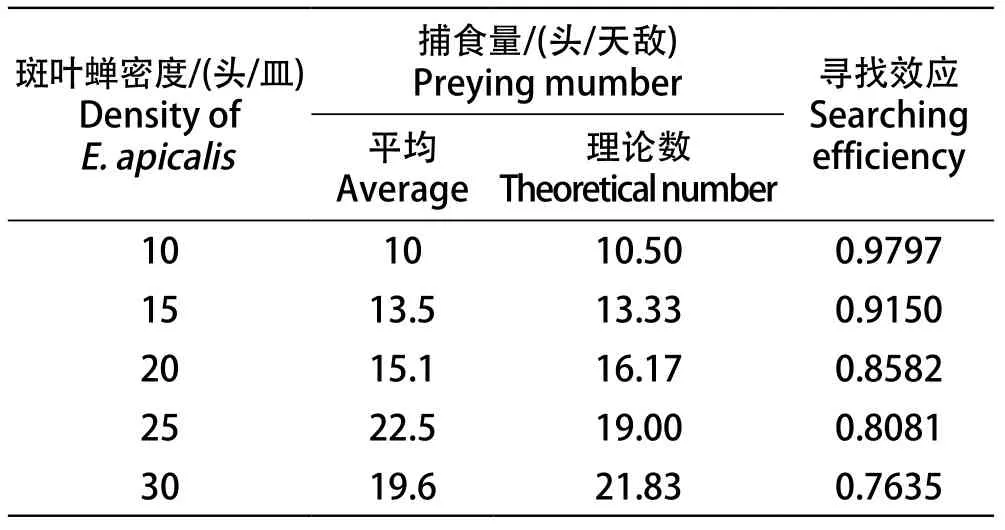
表1 多异瓢虫对不同密度葡萄斑叶蝉的寻找效应Table 1 Predation of H.variegate on different density of E. apicalis
在HollingⅡ圆盘方程的数学模型式中,Na是被捕食的葡萄斑叶蝉数量,Nt是葡萄斑叶蝉虫口密度,a是多异瓢虫对葡萄斑叶蝉的瞬时攻击率,T是多异瓢虫找到葡萄斑叶蝉的总时间,Th是捕食1头葡萄斑叶蝉所需的时间。计算出当df=3时,多异瓢虫1/N与1/Na之间的相关系数r为0.9781,r0.05=0.878,r0.01=0.959,r>r0.01。结果表明,两者密切相关,多异瓢虫对葡萄斑叶蝉的捕食功能反应拟合模型为:Na=1.1413Nt/(1+0.0165Nt),其中捕食1头葡萄斑叶蝉所需的时间Th为0.0145,瞬时攻击率a为1.1413,当葡萄斑叶蝉虫口密度为Nt→∞时,多异瓢虫的最大捕食量Namax为69头。为了测试模型的准确性,经卡方适合性检验,χ2值为0.5005,df=4时,χ20.05=9.49,χ2<χ20.05,拟合的结果很好。结果表明,上述模型能够较好的反映不同葡萄斑叶蝉密度下多异瓢虫的捕食变化规律。
寻找效应是捕食者捕食猎物的行为效应,取决于猎物昆虫种群的虫口密度,公式为S=a/(1+aThNt),表1显示了多异瓢虫对葡萄斑叶蝉的寻找效应,随着葡萄斑叶蝉虫口密度的增加,其寻找效应降低,当斑叶蝉为10头/皿时,其寻找效应为0.9797,而葡萄斑叶蝉为30头/皿时,其寻找效应下降到0.7635。
2.2 不同温度对多异瓢虫捕食功能的影响
由表2可以看出,在15~25 ℃的范围内,随着温度的升高,多异瓢虫捕食葡萄斑叶蝉的头数增加。多异瓢虫捕食率y与温度x之间的关系式为y=6x-62.33,r=0.9971,当df=1时,r0.05=0.997,r>r0.05,表明两者相关性显著。然而,在25~35 ℃范围内捕食率随着温度的升高出现降低,表明高温抑制了多异瓢虫的捕食活动。在25 ℃时,多异瓢虫捕食率最高,捕食率为89%,所以25 ℃是捕食的最佳温度。
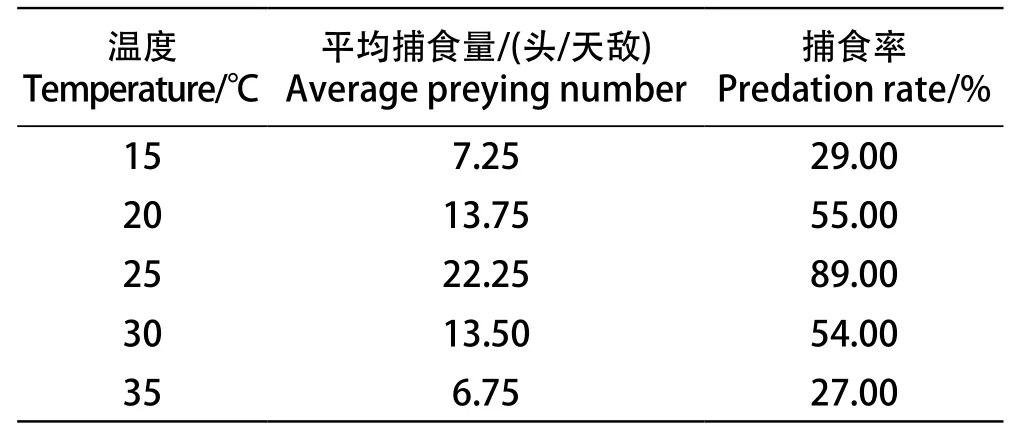
表2 不同温度对多异瓢虫捕食作用的影响Table 2 Predation of H. variegate under different temperature conditions
2.3 不同密度的多异瓢虫捕食的种内干扰
从表3可以看出,当葡萄斑叶蝉的密度为50头/皿时,多异瓢虫对成年葡萄斑叶蝉的平均捕食量随着自身密度的增加而逐渐降低,捕食作用率(E)也相应的降低。根据Hassell和Verley提出的干扰反应模型[8]:E=Q·P-m,或对数形式logE=logQ-mlogP,式中,E为竞争下的捕食作用,Q是搜索常数,P是捕食者密度,m是干扰系数。计算出搜索常数Q为0.4194,干扰系数m为0.7831。多异瓢虫捕食成年葡萄斑叶蝉过程中的自我干扰方程为:E=0.4194P-0.7831。经卡方检验,得χ2=0.046<χ2(0.05,3)=7.82。结果表明,上述模型可以用来描述多异瓢虫捕食过程中的自干扰。
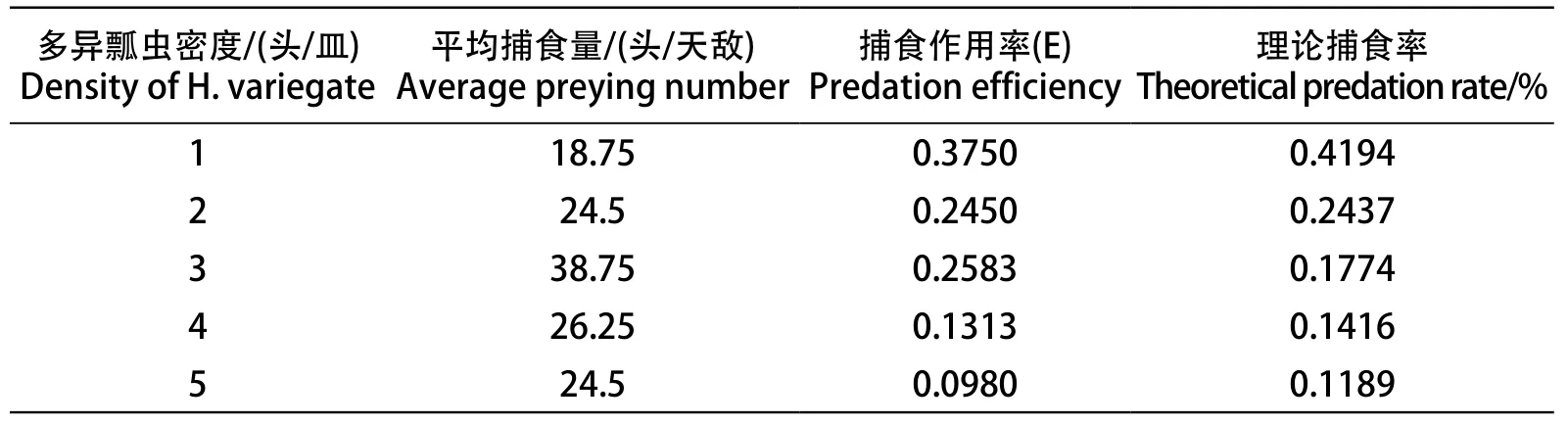
表3 不同密度下多异瓢虫对捕食量的干扰反应Table 3 Interference response of different densities of H. variegate to predation
2.4 多异瓢虫捕食作用对斑叶蝉分布的反应
从表4中的试验结果来看,葡萄叶片的数量与多异瓢虫捕食率成负相关,即随着叶片数量的增加,多异瓢虫的捕食率逐渐降低。这是由于天敌的活动范围和葡萄斑叶蝉的密度,取决于每单位面积的葡萄叶数。在一定范围内,叶片数量越多,天敌活动范围越宽,葡萄斑叶蝉的虫口密度越小;这将导致多异瓢虫与葡萄斑叶蝉之间的距离变得更大,天敌搜寻猎物的机会越少,捕获难度越大,捕食率越低。

表4 不同密度叶片数条件下的捕食率Table 4 Predation rate under different leaf numbers
3 讨论与结论
Holling提出根据反应曲线的形状将功能反应分为3种基本类型。在本试验条件下,多异瓢虫对葡萄斑叶蝉的捕食功能反应随着葡萄斑叶蝉密度的增加而增加,其捕食率随着葡萄斑叶蝉的密度呈负增加,这与HollingⅡ型功能反应模型是一致的[9-11]。这与于江南等[12]报道的研究结果相类似。
多异瓢虫对葡萄斑叶蝉的最大日捕食数量为69头,其葡萄斑叶蝉的a/Th为78.71,结果表明,多异瓢虫对葡萄斑叶蝉的防治能力很强。在贺兰山东麓地区,多异瓢虫发生在4月底至10月初,应注意保护和利用其对葡萄斑叶蝉的综合防治的应用
试验结果表明,多异瓢虫对葡萄斑叶蝉在15~25 ℃范围内的捕食作用随温度升高而增加,捕食率随着温度的升高在25~35 ℃范围内降低,捕食效果在25 ℃时最强。这与葡萄斑叶蝉发生的最适温度22~28 ℃基本一致,这可能与多异瓢虫和葡萄斑叶蝉的长期协同进化中形成的中间关系有关。
种内干扰实验结果表明,随着多异瓢虫数量增加,捕食作用率逐渐减少,说明多异瓢虫种内存在相互干扰。
猜你喜欢
今日农业(2021年3期)2021-12-05
今日农业(2021年9期)2021-07-28
今日农业(2021年4期)2021-06-09
天津农林科技(2021年2期)2021-04-16
红蜻蜓·低年级(2020年9期)2020-09-26
考试与评价·高二版(2020年4期)2020-09-10
知识就是力量(2020年2期)2020-05-19
发明与创新·大科技(2019年2期)2019-06-26
小学生作文(低年级适用)(2018年4期)2018-11-29
销售与市场(管理版)(2017年3期)2017-03-28
