
鱼类脂肪酸结合蛋白研究进展
2019-07-23 00:33隋修磊张玉茹
水产科学 2019年4期
隋修磊,张玉茹
( 河南师范大学 水产学院,河南 新乡 453007 )
脂肪酸结合蛋白(FABPs)是一类来自共同祖先基因的小分子蛋白质,相对分子质量为14~16 ku,广泛存在于动物肠道、心脏、脑、脂肪和肌肉等多种组织的细胞内,占细胞内可溶性蛋白总量的1%~8%[1]。研究表明,机体内脂肪酸跨膜运输是被动扩散或由膜蛋白介导的过程,脂肪酸结合蛋白主要结合细胞内游离的脂肪酸(FA)和其他疏水性配体,并将其输送到过氧化物酶体、线粒体和细胞核等细胞器,进而调控脂肪酸氧化和甘油三酯及磷脂合成等脂代谢过程[2](图1)。脂肪酸结合蛋白家族成员的命名是根据其首次分离或鉴定出的组织来命名的,例如肠型脂肪酸结合蛋白(I-FABP)、肝型脂肪酸结合蛋白(L-FABP)等。然而,很多组织中往往有多种脂肪酸结合蛋白基因(fabps基因)同时表达,研究者又对不同的脂肪酸结合蛋白基因按阿拉伯数字命名[3],如fabp1基因(肝型)、fabp2基因(肠型)、fabp3基因(心脏型)等。自1972年Ockner等[4]从大鼠的小肠中分离出肠型脂肪酸结合蛋白以来,研究者已经在脊椎动物基因组中鉴定出了12种不同的脂肪酸结合蛋白[5]。不同类型的脂肪酸结合蛋白均由125~135个氨基酸组成,其氨基酸序列相似性由20%至70%不等[6]。虽然不同脂肪酸结合蛋白序列多变,但脂肪酸结合蛋白基因和蛋白结构相对保守,大多基因由4个外显子和3个内含子组成[7-8],其上游区域存在大量的调控元件,同时保守存在一个TATA盒[9-10](图2)。结构生物信息学研究表明,多数脂肪酸结合蛋白由10个反向平行的β片层(βA-βJ)和2个α-螺旋结构组成,其中β片层折叠成“桶”状结构,通过范德华力和疏水相互作用等结合不同的疏水性配体[11];氮端的α螺旋(α-Ⅰ和α-Ⅱ)形成“帽子”样结构控制着疏水性配体的出入[12]。研究还发现,脂肪酸结合蛋白和生物体的许多生命过程相关,它们在生物体的能量贮存、信号转导、生物膜的合成、过氧化物酶体的氧化、酶活力的调节、机体免疫反应以及细胞的增殖和分化等过程中发挥着重要的生物学功能[13-15]。
与哺乳动物不同,淡水硬骨鱼类具有Δ12及Δ15等脂肪酸去饱和酶,可以把亚油酸和亚麻酸转化为二十碳五烯酸和二十二碳六烯酸等多聚不饱和脂肪酸储存在肌肉等组织中[16-17]。因此,鱼类是人体多不饱和脂肪酸的重要来源,鱼体中脂肪酸结合蛋白能转运脂肪酸参与脂代谢反应,是重要的脂肪性状相关基因,对鱼类脂肪的沉积具有重要调控作用,鱼体中脂肪含量的高低是影响鱼肉品质的重要因素。因此对脂肪酸结合蛋白及其基因进行深入、全面、系统研究,对于水产养殖具有重要意义。目前,研究者在哺乳动物脂肪酸结合蛋白的种类、分布、结构、功能和基因表达等方面取得了突破性的研究进展;家畜和家禽类中脂肪酸结合蛋白的研究主要集中在生产性状的改良、畜禽产品品质的提升和品种及品系的选育等方面;但是,鱼类中脂肪酸结合蛋白的研究目前主要集中在基因的克隆、组织表达和表达模式等方面,其基因结构、种类和基因及蛋白的功能等方面有待进一步深入研究。因此,笔者对鱼类脂肪酸结合蛋白的种类、基因表达以及染色体定位等研究进行了综述,以期为鱼类中脂肪酸结合蛋白超家族基因及蛋白的演化和功能分析以及鱼类营养调控提供理论依据。
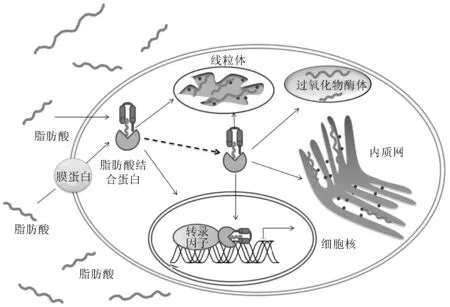
图1 脂肪酸结合蛋白结合和转运脂肪酸示意[18-19]

图2 fabp2基因结构示意[8,20]22aa、58aa、36aa和16aa表示外显子所编码的氨基酸的个数;横线上的数字表示内含子的长度,单位bp.
1 鱼类中脂肪酸结合蛋白的类型

上述鱼类中主要包含fabp1、2、3、6、7、10、11七种类型基因,其中大西洋鲑中没有fabp1和fabp11基因,可能是由于其基因组信息不完善导致暂未发现,或在进化过程中丢失。在斑马鱼、虹鳟和大西洋鲑等中存在多种旁系同源基因,而雀鳝中的脂肪酸结合蛋白基因都以独立体基因存在,没有同源基因,可能是由于不同鱼类进化模式不同导致的[38]。鱼类进化分析结果表明,作为一类原始的淡水鱼类,雀鳝没有经历硬骨鱼类特有的全基因组倍增事件[45-46],被称为鱼类进化研究的“活化石”。
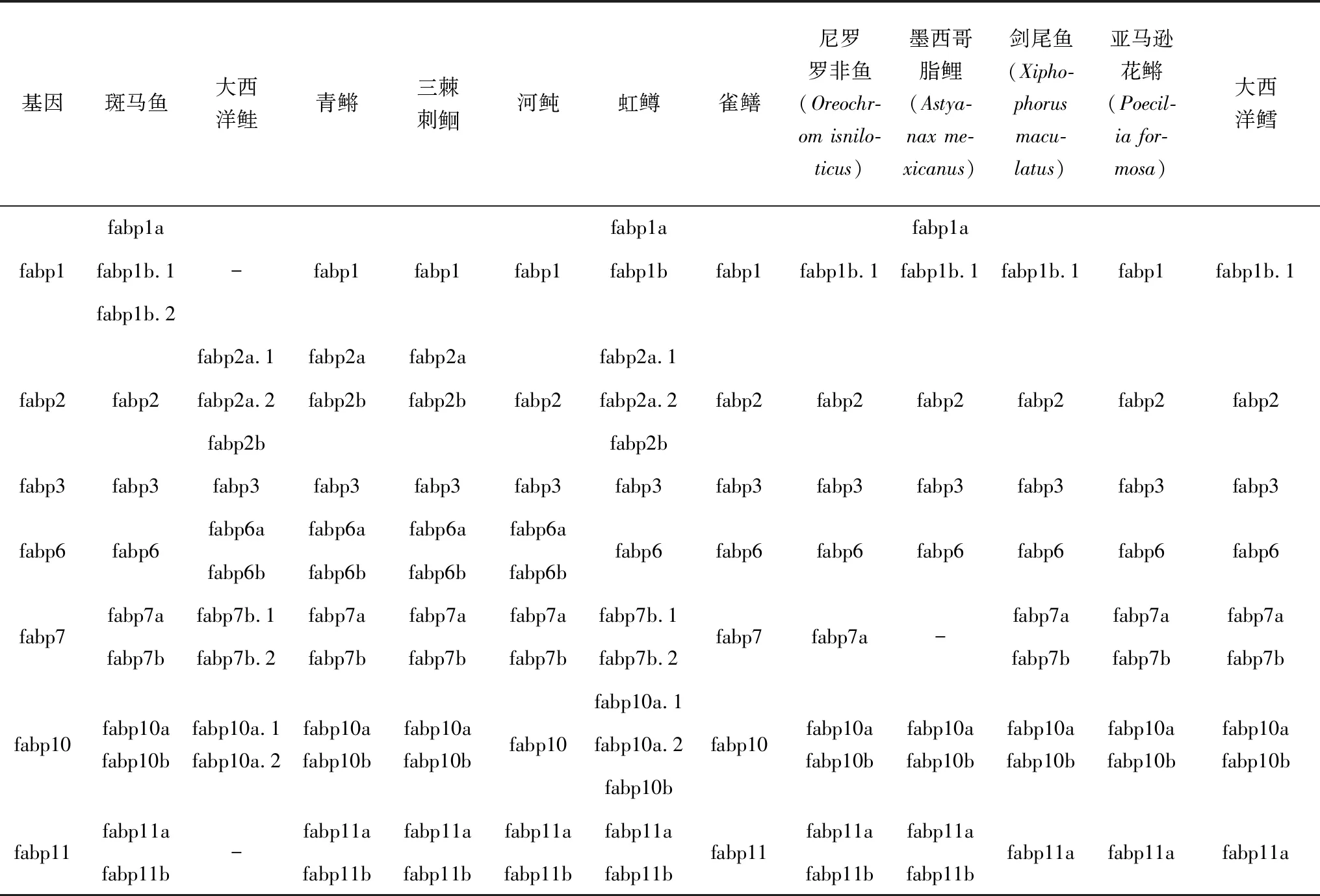
表1 代表鱼类中脂肪酸结合蛋白基因
Venkatachalam等[38]在研究雀鳝脂肪酸结合蛋白基因结构、染色体定位和基因演化时发现,其脂肪酸结合蛋白基因没有经历硬骨鱼类特有的全基因组倍增事件,因此可以作为脊椎动物中脂肪酸结合蛋白基因演化分析时的外群。
国内外研究报道论述了几种常用试验鱼类基因中的脂肪酸结合蛋白基因类型,明晰了鱼类中都存在哪些脂肪酸结合蛋白基因。作为一类重要的脂肪酸结合和转运蛋白,脂肪酸结合蛋白基因的种类和鱼类脂肪酸的转运、储存和代谢等活动息息相关[35,47-49]。研究表明,脂肪酸结合蛋白由于亚型结构上的微小区别,其结合长链脂肪酸时在配体的选择、亲和力的大小和结合机制上都不同。一般来说,除不饱和脂肪酸,配体的疏水性越强,亲和力越大[50]。靶细胞本身的特性可能决定了不同组织主要脂肪酸结合蛋白亚型的亲和力甚至选择性。例如,脑型脂肪酸结合蛋白对长链脂肪酸如二十二碳六烯酸有很高的选择性[51]。因此,该部分结果可为鱼类不同食物饲喂下脂肪酸的摄取、动员、代谢和存储等问题提供新的研究视角,从而为渔业生产中植物油替代鱼油等新型饲料的研发提供理论基础。
2 鱼类脂肪酸结合蛋白基因的组织表达
由表2可知,鱼类中fabp1和fabp10a基因主要在肝脏中表达,这表明现存硬骨鱼类中fabp10基因可能保留了祖先fabp1基因的功能,而fabp10b基因在肝脏中的表达量非常低或完全检测不到,这可能是硬骨鱼类fabp10a和fabp10b基因的空间转录调节分裂造成的[37];哺乳动物和鱼类中fabp2基因的表达情况相似,都主要在肠道中表达;fabp3基因主要在鱼类的肝胰脏和肌肉中表达;鱼类的fabp6基因主要在肠道中表达,而有些鱼类中的fabp6b基因在其他组织中表达量较高,这可能是进化过程中发生了亚功能化造成的[32,41];不同鱼类的fabp7基因及其同源基因在不同组织中的表达优势各异;fabp11基因主要在鱼类的眼中表达,个别在肾脏中表达量较高。
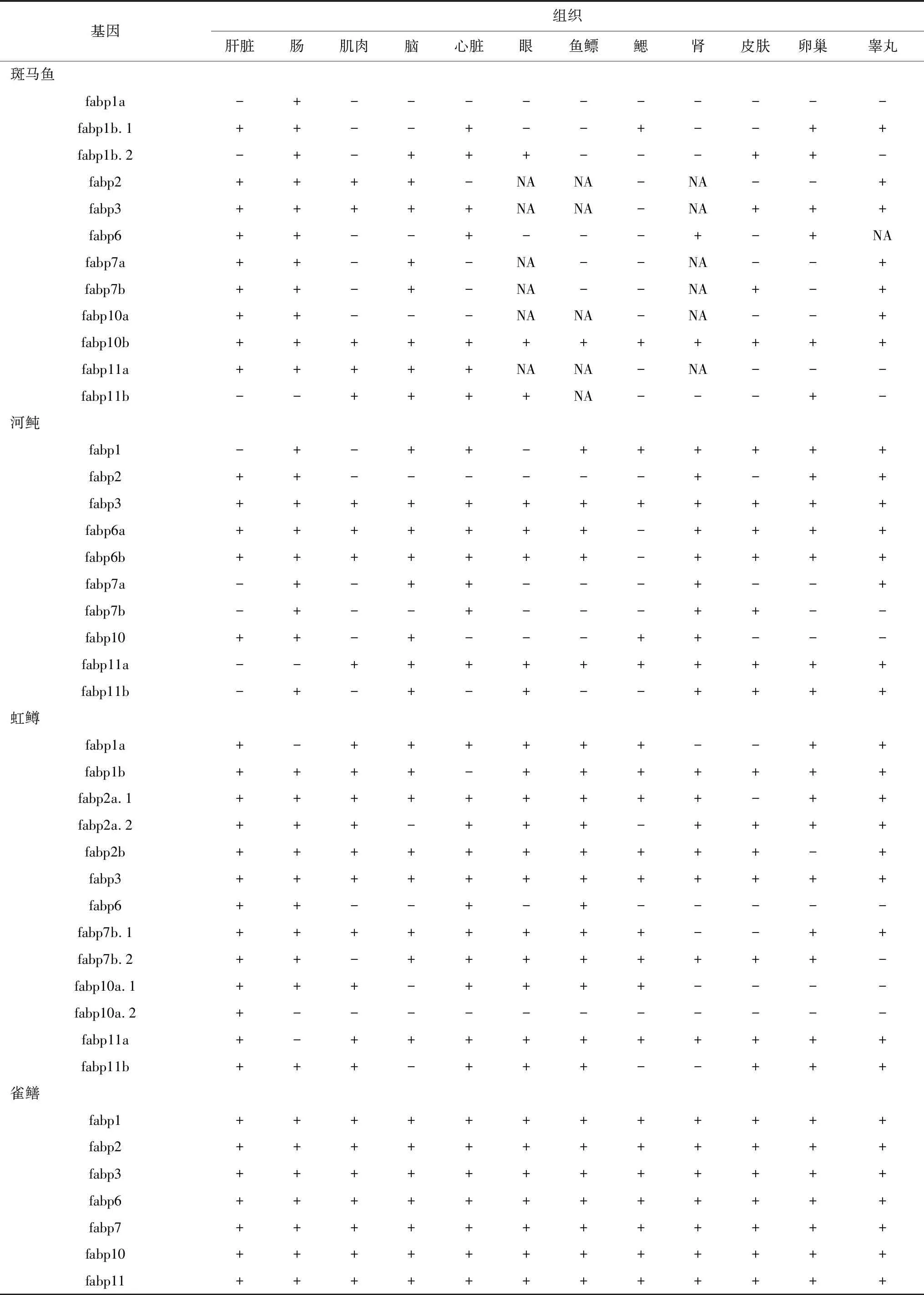
表2 几种代表鱼类脂肪酸结合蛋白基因的组织表达情况
注:“+”表示在该组织中表达;“-”表示在该组织中不表达;“NA”表示没有在该组织中检测表达情况.
2.1 肝脏型脂肪酸结合蛋白
肝脏型脂肪酸结合蛋白(L-FABP或FABP1)主要存在于哺乳动物的肝脏和肠道中[52-54]。不同鱼类中fabp1基因的组织表达各异。斑马鱼和河鲀中的fabp1基因主要在肠中富集[27,36,55],并广泛存在于心脏、鳃、卵巢和睾丸等组织,除斑马鱼肝脏中有fabp1b.1基因少量表达外,其他类型的fabp1基因均未在斑马鱼和河鲀中检测到[27,36,55]。与之相对,草鱼(Ctenopharyngodonidellus)[56]和虹鳟[34]中的fabp1基因主要在肝脏中表达,在脾脏、肠、肾、心脏等组织中也有少量表达。此外,雀鳝中fabp1基因在心脏中表达量最高,在其他组织中也有表达[38]。
除了上述的肝脏型脂肪酸结合蛋白外,研究者在斑马鱼[22]、鲨鱼(Halaetunusbivius)[57]、肺鱼(Lepidosirenparadoxus)[58]、萨波雷氏鲇(Rhamdiasapo)[59]和鲤鱼(Cyprinuscarpio)[60]等肝脏中发现了肝脏基本型脂肪酸结合蛋白(Lb-FABP)。肝脏型脂肪酸结合蛋白和肝脏基本型脂肪酸结合蛋白是由同一祖先基因复制而来的旁系同源蛋白[61]。鲤鱼Lb-fabp基因在肝脏、胰脏和肠道中表达量较高,在脑、肌肉、心脏、卵巢和脾脏等组织中也有少量表达,但肠道和前肠中却未检测到该基因的表达[60]。在斑马鱼中,通过原位杂交和放射自显影的方法只在成体的肝脏中检测到Lb-fabp基因的表达[22]。而在萨波雷氏鲇中,免疫组化分析结果显示只在肝脏中检测到肝脏基本型脂肪酸结合蛋白[59]。
2.2 肠型脂肪酸结合蛋白
2.3 肌肉和心脏型脂肪酸结合蛋白
肌肉和心脏型脂肪酸结合蛋白(H-FABP或FABP3)在畜禽类中研究比较广泛,多集中在基因的多态性和肌内脂肪含量等方面的研究[71-73],而有关鱼类fabp3基因及其表达的研究报道尚少。目前已在斑马鱼[24]、齐口裂腹鱼(Schizothoraxprenanti)[74]、4种南极硬骨鱼类[75]、鲤鱼[74]、雀鳝[38]、鲨鱼[76]、兴凯湖翘嘴鲌(Culteralburnus)[77]、虹鳟[34]、建鲤[78]、瓦氏黄颡鱼(Pelteobagrusvachelli)[79]和河鲀[36]等鱼类中克隆得到了fabp3基因。利用原位杂交方法只在斑马鱼卵巢和肝脏组织中检测到fabp3基因表达,利用RT-PCR可检测到其他组织,如脑、心脏、肠、肌肉、皮肤和精巢等中fabp3基因的表达[24,28]。齐口裂腹鱼、鲤鱼、雀鳝、兴凯湖翘嘴鲌和河鲀的肝(胰)脏中fabp3基因表达量最高[36,38,74,77]。鳗鲡科、鲨鱼、拉氏南美南极鱼(Patagonotothenramsayi)和底鳉(Fundulusheteroclitus)的fabp3基因也主要在肝脏中表达[75,76,80-81]。而虹鳟的fabp3基因在肝脏、肠、幽门、胃、脑、肌肉和心脏等所有检测的组织中表达,其在肌肉中的表达量最高,在肝脏中的表达量相对较低[34]。
2.4 回肠型脂肪酸结合蛋白
2.5 脑型脂肪酸结合蛋白
2.6 肝脏基础型脂肪酸结合蛋白
2.7 肾脏型脂肪酸结合蛋白
肾脏型脂肪酸结合蛋白(K-FABP或FABP11)是鱼类特有的脂肪酸结合蛋白。大多鱼类中存在fabp11a和fabp11b两个旁系同源基因。与斑马鱼和青鳉中fabp11a和fabp11b基因的表达情况相似,河鲀的fabp11基因在眼中的表达最高[36],并且斑马鱼fabp11b基因转录产物仅在眼中检测到[87];虹鳟中fabp11a基因在眼中表达量最高,而fabp11b基因在幽门盲肠中表达量较其他组织丰富[34];雀鳝的fabp11基因在肝脏、肠、脑和肾脏等组织中存在,其中在肾脏中表达量最高[38]。
综上所述,不同类型的脂肪酸结合蛋白基因成员具有组织表达特异性。大部分脂肪酸结合蛋白基因主要在某些特定的组织或细胞类型中表达,如鱼类中的fabp1和fabp10a基因主要在肝脏中表达;但并不是每一种类型只特异地存在于一种组织或细胞,如心脏型脂肪酸结合蛋白和表皮型脂肪酸结合蛋白广泛存在身体多个部位[88]。不同组织脂肪酸结合蛋白基因表达水平的比较分析可为脂肪酸结合蛋白基因的功能演变和转录调节提供理论基础。进化分析结果表明,脂肪酸结合蛋白基因家族是由一个共同祖先基因复制演化而来。基因复制可以有效地增加生物体的复杂性,但在进化过程中不允许复制后的多基因发挥同样的功能从而出现功能的冗余性,因此,复制后的多基因功能在进化过程中会出现不同程度的演变。复制基因的共同命运主要有以下3种:(1)原功能退化丢失;(2)演化出不同于祖先基因新功能;(3)保留祖先基因的一部分亚功能[35]。复制基因功能的演变是因不同复制基因蛋白质编码区的突变和启动子中顺式作用调节元件的丢失或获得等突变事件所致[82]。如Laprairie等[35]研究斑马鱼中fabp1三个复制基因(fabp1a、fabp1b.1和fabp1b.2)的功能演化时发现,斑马鱼fabp1a基因的启动子含有PPARα识别位点,fabp1b.1基因的启动子含有PPARγ作用位点,而fabp1b.2基因的启动子无PPAR识别位点。该研究中,PPARs通过特异性地识别脂肪酸结合蛋白基因上游的启动子区域调控斑马鱼中fabp1基因的3个复制基因的组织表达和功能。此外,很多由鱼类特有的全基因组倍增复制而来的脂肪酸结合蛋白基因(如虹鳟中fabp7b.1/fabp7b.2和fabp10a.1/fabp10a.2基因)具有相似的组织表达特性,预示这些基因的表达由保守的转录调节因子控制[34],但调控脂肪酸结合蛋白基因表达的具体的转录原件和调控因子仍需深入发掘和阐明。
3 鱼类中脂肪酸结合蛋白基因染色体定位
鱼类中各脂肪酸结合蛋白基因种类和位置信息可为fabps基因结构和功能的演化过程奠定理论基础。系统发育分析结果表明,脂肪酸结合蛋白基因家族是由一个共同祖先基因复制演变而来。据估计,第1次基因复制发生在大约9.3亿年前,随后的基因复制发生在距今5亿年前[90],鱼类中的脂肪酸结合蛋白基因还存在第3次鱼类特有的基因组复制事件。根据基因组中脂肪酸结合蛋白连锁基因存在情况,可以分析该基因是由祖先基因直接垂直遗传而来还是由祖先基因复制而来,不同的物种中是否出现了特定脂肪酸结合蛋白基因丢失等进化过程,进而深入分析脂肪酸结合蛋白基因结构和功能的演化历程。
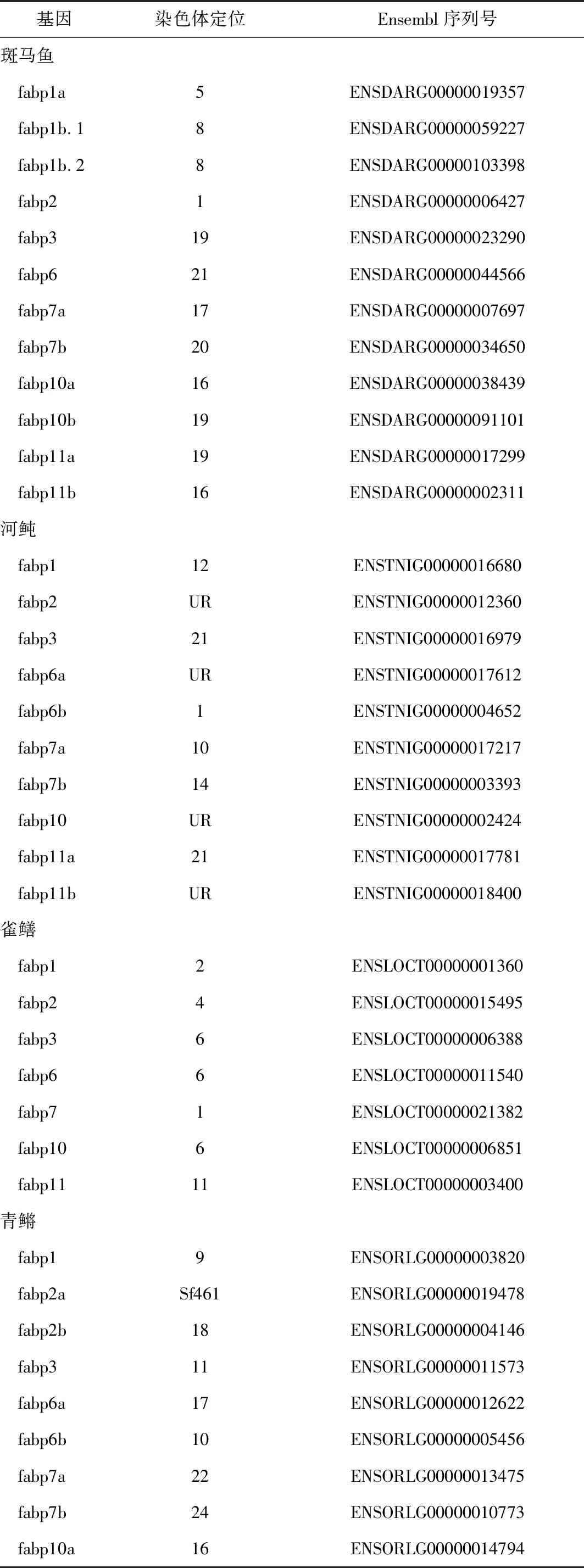
表3 几种代表鱼类的脂肪酸结合蛋白基因定位[36-38,89]
续表3
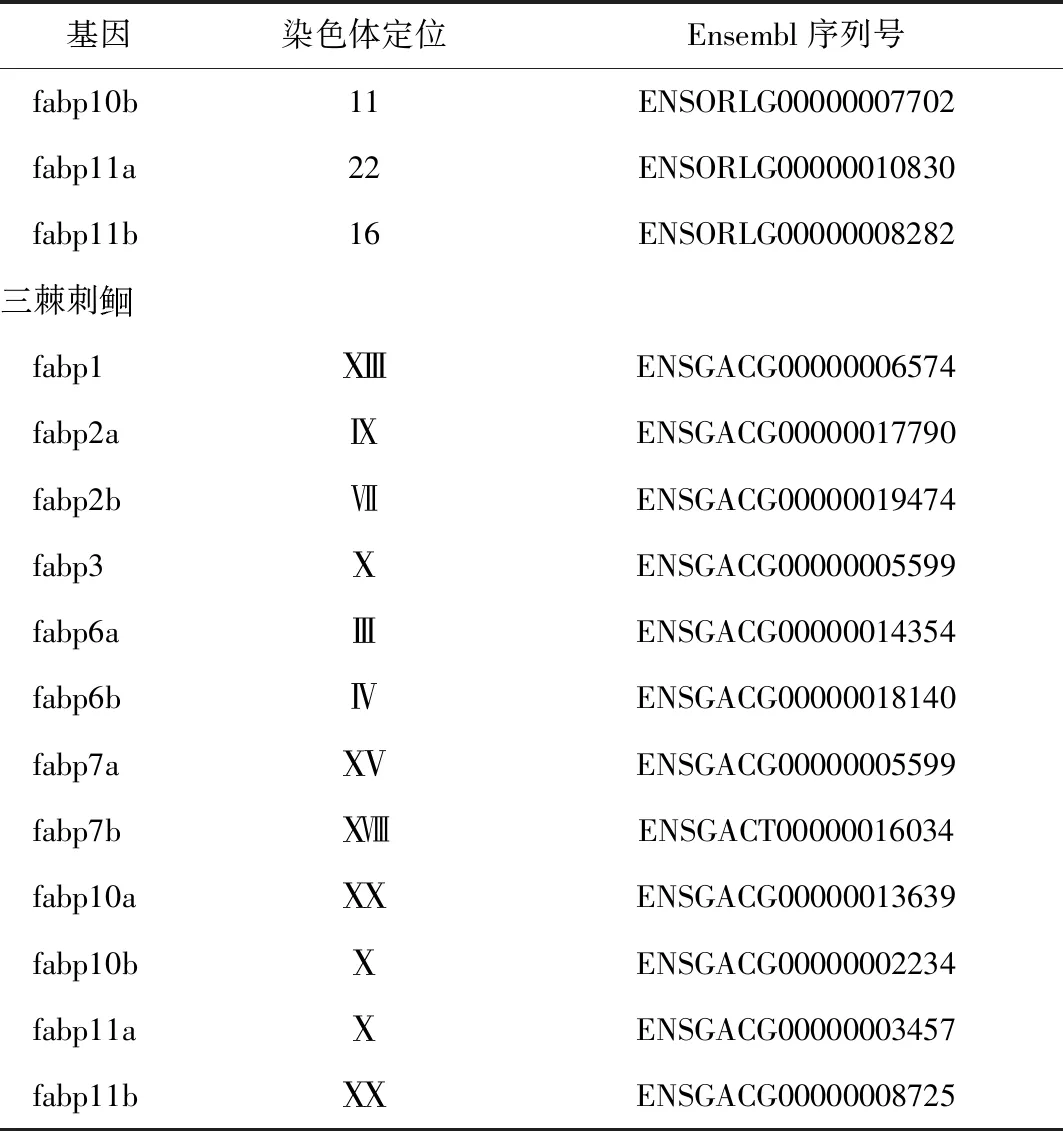
基因染色体定位Ensembl序列号fabp10b11ENSORLG00000007702fabp11a22ENSORLG00000010830fabp11b16ENSORLG00000008282三棘刺fabp1ⅩⅢENSGACG00000006574fabp2aⅨENSGACG00000017790fabp2bⅦENSGACG00000019474fabp3ⅩENSGACG00000005599fabp6aⅢENSGACG00000014354fabp6bⅣENSGACG00000018140fabp7aⅩⅤENSGACG00000005599fabp7bⅩⅧENSGACT00000016034fabp10aⅩⅩENSGACG00000013639fabp10bⅩENSGACG00000002234fabp11aⅩENSGACG00000003457fabp11bⅩⅩENSGACG00000008725
注:Sf,基因组构架;UR,非随意染色体.
4 总结与展望
自20世纪70年代发现脂肪酸结合蛋白以来,科研人员对其进行了大量的研究。然而,现有的研究多关注哺乳动物中脂肪酸结合蛋白的结构、功能和基因表达,鱼类上的相关研究数据报道仍然不足。如,和哺乳动物相比鱼类中有哪些脂肪酸结合蛋白?他们在不同的组织中如何表达?不同鱼类脂肪酸结合蛋白种类和表达是否相同?为了探究上述问题,笔者系统的总结了鱼类中脂肪酸结合蛋白的种类、组织表达和染色体定位情况。通过归纳总结和比较分析发现,鱼类中共发现了7种类型fabps基因,分别是fabp1、fabp2、fabp3、fabp6、fabp7、fabp10和fabp11;与哺乳动物相比,代表鱼类中脂肪酸结合蛋白基因的旁系同源基因很多,如fabp1a、fabp1b.1和fabp1b.2基因,但fabp3基因在鱼类和哺乳动物基因组中都以独立基因形式存在;脂肪酸结合蛋白基因在多种组织中广泛表达,fabp1基因主要在肝脏和肠中表达,fabp3基因在上述鱼类的所有组织中表达,而斑马鱼的fabp1a基因仅在肠中表达;此外,通过Ensembl数据库搜索和文献阅读发现,脂肪酸结合蛋白基因在鱼类和人等物种中存在广泛的共线性,并总结了几种代表鱼类中脂肪酸结合蛋白基因在染色体上的位置。作为影响鱼体脂肪沉积部位和鱼肉脂肪酸成分及含量的候选基因,脂肪酸结合蛋白的种类、组织分布、多态性和遗传变异调节了鱼肉品质和鱼体脂代谢的平衡。因此,该家族基因及蛋白的研究可应用到水产动物育种和代谢性疾病的防治等实践中。比如,可为水产养殖中的水产品品质的提升和特色水产品培育(如富含二十碳五烯酸和二十二碳六烯酸的鱼类)提供参考;还可为目前集约化养殖环境下养殖鱼类脂肪过度蓄积引发的脂肪肝等营养代谢性疾病的预防和治疗提供新的研究视角。但是,鱼类中脂肪酸结合蛋白如何调控脂肪酸的转运,不同类型的脂肪酸结合蛋白对不同的底物是否具有不同的结合能力,脂肪酸结合蛋白又是如何与脂代谢通路的上游转录因子和下游脂肪合成与分解的酶协同作用调控脂肪的合成与分解,脂肪酸结合蛋白调控鱼类脂代谢的具体功能机制是怎样,这些问题都有待深入研究。
猜你喜欢
化工管理(2022年13期)2022-12-02
安徽医科大学学报(2022年6期)2022-07-13
小天使·二年级语数英综合(2021年8期)2021-08-16
中国生殖健康(2020年5期)2021-01-18
健康博览(2020年11期)2020-11-23
儿童时代·幸福宝宝(2020年9期)2020-09-08
探索科学(2017年5期)2017-06-21
作文评点报·作文素材小学版(2016年8期)2016-03-16
第二课堂(小学版)(2015年2期)2015-05-22
食品工业科技(2014年13期)2014-03-11
