
嫁接对橡胶树砧木甲基化水平的影响
2019-07-19 02:23毛常丽张凤良李小琴倪书邦
热带农业科技 2019年3期
毛常丽,张凤良,李小琴,杨 湉,倪书邦,吴 裕
(云南省热带作物科学研究所,景洪666100)
表观遗传学(epigenetics)是研究表观遗传变异的遗传学分支学科,表观遗传变异(epigenetic variation)是指在基因的DNA序列没有发生改变的情况下,基因功能发生了可遗传的变化,并最终导致表型的变化,其主要涉及DNA与染色体上的一些可逆修饰以及一些转录调控机制。DNA甲基化、组蛋白修饰和非编码RNA调控是表观遗传学研究的主要方向,并在植物生长发育、逆境胁迫或环境适应过程中发挥着重要的作用[1-3]。其中,DNA甲基化是基因组DNA的一种最重要的表观遗传修饰方式,MSAP技术是在AFLP技术上建立起来的[4],已被广泛用于检测植物在高温[5-9]、低温[10-12]、干旱[13-15]等非生物胁迫条件下 DNA甲基化水平变化的研究。
橡胶作为重要的物资,是国民、国防不可或缺的资源,天然橡胶是橡胶工业发展的基础,其来源主要是巴西橡胶树(Hevea brasiliensis)。天然橡胶种植材料为橡胶树芽接苗,橡胶树的生长、产量与砧木和接穗以及栽种的立地环境息息相关。关于砧木与接穗间的相互影响,已经在黄瓜[16]、葡萄[17]、阿月浑子[18]以及番茄[19]等作物上有过研究。在橡胶树上,砧木与接穗间的研究包括蛋白质组学[20]、径向生长差异[21]、生化[22]、内源激素[23]等。在表观遗传学方面,黄关青等构建了橡胶树的DNA甲基化过滤文库[24];李海林等对橡胶树全基因组水平DNA甲基化位点进行检测,确定巴西橡胶树基因组DNA甲基化模式和水平[25];吴春太等建立了橡胶树的MSAP反应体系,并分析了无性系DNA的甲基化水平[26]。对于橡胶树砧木与接穗间的甲基化水平如何,至今未见报道。
本研究利用MSAP技术对巴西橡胶树砧木与接穗间的全基因组DNA甲基化水平进行了分析,为巴西橡胶树高产育种提供分子辅助手段及理论基础。
1 材料和方法
1.1 实验设计说明
从橡胶树无性系IAN873、GT1和RRIM600的开放授粉子代中各选1株作为采穗原株(分别记为 SI、SG、SR);同时以这 3 个无性系的半同胞家系为砧木(分别记为RI、RG、RR)。砧木长到地径约1.5 cm时,在离地30~40 cm高度锯干,从抽生的新芽中选择方向相对的两枝留下,其余抹除。待留下的两枝长到可嫁接粗度后开始嫁接工作。

图1 橡胶树自接和对接
实验设计为3个采穗原株与3个家系建立9个砧穗组合(3×3),即得到9株砧木的双叉植株(图1)。为了减少嫁接过程本身造成的差异,从砧木本株采一芽片嫁接,称为“自接”(图1中,A);按实验设计建立的砧穗组合称为“对接”(图1中,B)。
1.2 材料
砧木采样:在嫁接前,采集砧木 RI、RG、RR的变色期叶片,硅胶干燥保存。自接和对接采样:嫁接完成后第二年全部锯干,待自接和对接的芽片都抽生枝条且正常生长到2蓬叶以上,采集其变色期叶片,硅胶干燥保存。即包括9株砧木嫁接前的叶样、嫁接后自接叶样和对接叶样,共27个样品。
1.3 方法
MSAP实验委托生工生物工程(上海)股份有限公司完成。采用EcoRⅠ/MspⅠ和EcoRⅠ/HpaⅡ进行酶切,用3730XL测序分析仪(美国ABI公司)对MSAP选择性扩增PCR产物进行电泳分离扩增片段,得到的峰图用Genemapper 4.0软件进行分析。实验用到的接头及引物相关信息见表1。

表1 M S A P分析所用的接头及引物信息
2 结果和分析
2.1 橡胶树砧木嫁接后的甲基化模式
对EcoRⅠ/MspⅠ和EcoRⅠ/HpaⅡ酶切产物经过连接、2轮扩增后,用3730XL测序分析仪进行检测,得到MSAP峰图(图2、图3)。将得到的峰图进行转换,得到0/1数据(“0”表示扩增后无带,“1”表示扩增后有带)。经统计,橡胶树嫁接后出现以下3种甲基化模式:当E+M有带,E+H无带时,表明CCGG位点发生双链DNA内甲基化,即全甲基化位点;当E+M无带,E+H有带时,表明CCGG位点发生单链DNA外甲基化,即半甲基化位点;当E+M和E+H都有带时,表明CCGG位点未发生甲基化,即非甲基化位点。

图2 I A N 8 7 3砧木用引物组合E+M扩增所得峰图
图3 I A N 8 7 3砧木用引物组合E+H扩增所得峰图
2.2 橡胶树砧木嫁接后的甲基化水平
针对橡胶树砧木与接穗基因组DNA的甲基化,共选出8对引物进行扩增。根据转换得到的0/1数据,8对引物共扩增得到13 922条带,其中甲基化位点6 862个(表2),甲基化率为49.29%,甲基化位点中全甲基化位点2 859个(41.66%),半甲基化位点4 003个(58.34%)。

表2 橡胶树砧木甲基化位点
对3种砧木嫁接前后甲基化模式之间的转换进行统计,结果见表3。
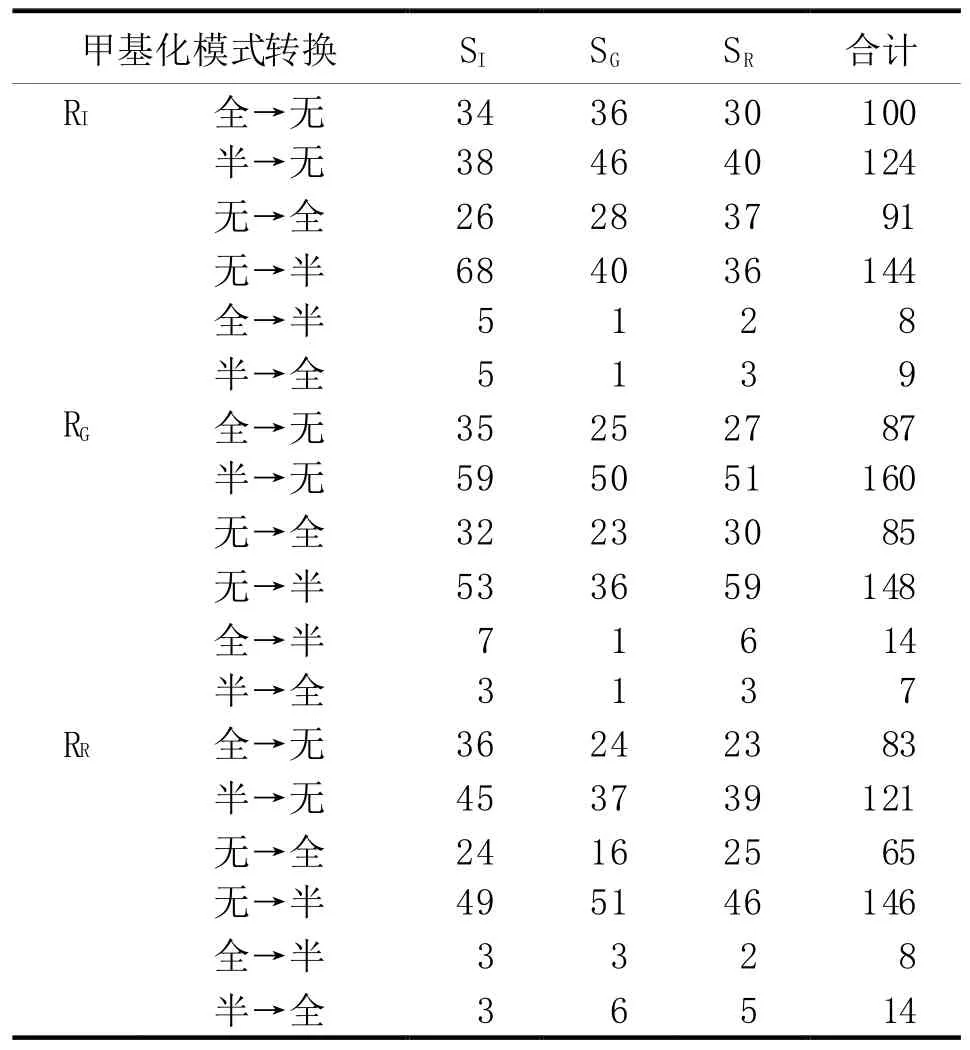
表3 橡胶树嫁接前后的甲基化模式转换
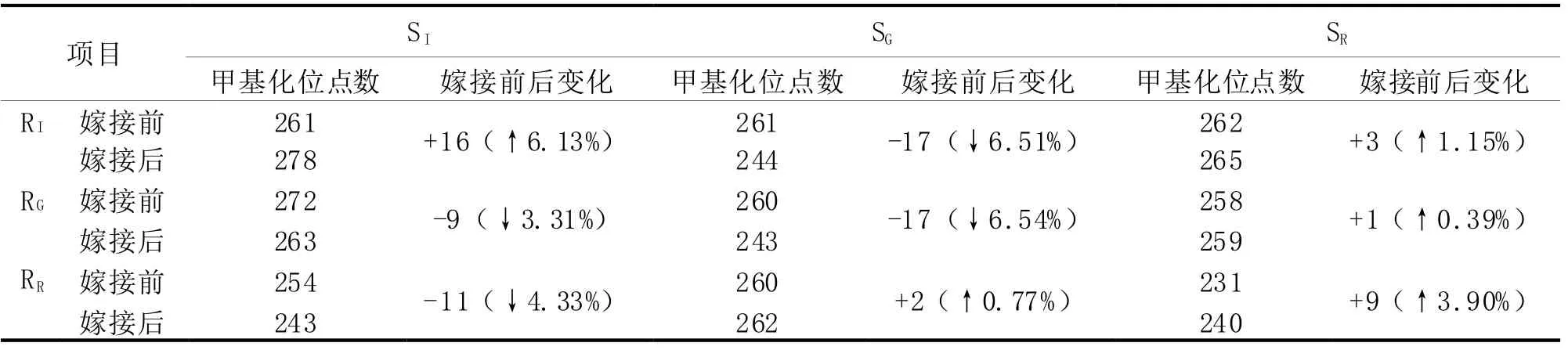
表4 橡胶树砧木嫁接前后甲基化位点变化
统计以上数据,3种接穗嫁接于3种砧木后,以半甲基化与无甲基化间的相互转换为主,而由半甲基化转化为全甲基化或者由全甲基化转化为半甲基化的很少,全甲基化与无甲基化间的转换居中。
根据以上数据,3种接穗嫁接于3种砧木后,对IAN873砧木(RI)的甲基化位点数的影响为SI>SG和SR;对GT1砧木的甲基化位点数的影响为SI/SG>SR;对IAN873砧木的甲基化位点数的影响为 SI>SG∕SR。
3种砧木嫁接前后甲基化变化情况见表4。从表4中的数据可以看出,IAN873砧木嫁接IAN873接穗后,甲基化位点数增加了16个,嫁接GT1接穗后,甲基化位点减少了17个,而RRIM600嫁接到IAN873后引起的甲基化位点数变化不大;当以GT1为砧木时,引起甲基化位点变化较大的接穗是IAN873及GT1,而RRIM600变化不大;当以RRIM600为砧木,接穗IAN873使嫁接后的甲基化位点数下降,而嫁接GT及RRIM600后甲基化位点增加,且RRIM600接穗引起位点数增加的幅度大于GT1接穗。
3 讨论
在植物中,DNA甲基化主要参与植物基因表达的调控,进而调节植物的生长发育[27],其主要通过基因的甲基化和去甲基化来调控基因的表达。本研究探讨了橡胶树砧木嫁接后的甲基化水平变化,结果表明:嫁接后砧木的甲基化水平发生了变化,且同一砧木嫁接不同的接穗引起的甲基化变化有差异;嫁接使砧木的甲基化模式发生改变,主要以半甲基化与无甲基化间的相互转换为主。
橡胶树种植材料为嫁接的优良无性系,橡胶树的生长、产量等性状取决于接穗品种、栽培的立地环境以及砧木所能提供给树体的养分情况等,砧木品种的选择对橡胶树嫁接及性状表现至关重要。本研究发现,嫁接可以改变砧木的甲基化水平及模式。Avramidou等[28]也发现,嫁接可以引起接穗甲基化水平的改变,使接穗的甲基化水平上升。李海林等[25]对巴西橡胶树DNA甲基化的位点进行分离测序,得到橡胶树基因组中包括转录因子、蛋白激酶等在内的多种类型的DNA序列中均存在甲基化现象。因此,橡胶树嫁接后,可能在砧木与接穗间有相关的基因被甲基化或者去甲基化表达,且在砧木与接穗间相互转移,从而导致嫁接对砧木及接穗的影响。而嫁接后砧木与接穗间甲基化及相关物质间转移的调控机制还需作进一步的实验进行验证。
猜你喜欢
热带生物学报(2022年1期)2022-03-09
农业科技与信息(2021年9期)2021-12-07
农业科技与信息(2021年8期)2021-12-06
福建基础教育研究(2019年10期)2019-05-28
小学生导刊(低年级)(2016年11期)2016-11-14
世界热带农业信息(2016年6期)2016-07-02
数学大王·中高年级(2016年8期)2016-05-14
数学大王·中高年级(2014年7期)2014-08-06
读写算·小学低年级(2009年10期)2009-10-27
