
铝对蚕豆叶绿素荧光和叶绿素含量影响探讨
2019-07-15 11:13张慧敏刘东华
绿色科技 2019年12期
张慧敏,刘东华
(1.山西省大同云冈区园林管理处,山西 大同 037006;2.天津师范大学,天津 300074)
1 引言
自然界中铝分布极广,地壳中的铝含量为7.35%,仅次于氧和硅。铝在自然界的化学性质十分活泼,铝离子存在状态与溶液pH值密切相关。在pH<5.0的酸性水体中,水溶性铝是植物重要的生长限制因子。随着酸雨沉降和酸性肥料的施用,土壤酸化加剧,铝溶出量增加,环境中活性铝成为酸性土壤中影响植物生长发育的重要因素。酸性土壤危害粮食作物减产,因此,改良酸性土壤,提高作物产量,是当前亟待解决的一项重要任务。
叶绿素荧光动力学技术可快速、无损伤的反映叶片光合功能内在特征。植物长期处于不良环境中,会出现叶绿素含量降低,光合作用速率下降等症状,进而表现为荧光参数发生变化。铝处理可抑制玉米和大豆叶片叶绿素的合成,降低了大豆叶片叶绿素含量,叶片叶绿素a/b含量比也有一定下降趋势,光合速率和蒸腾速率下降。Al3+毒害引起的叶绿素总量下降,可能是金属离子抑制叶绿素酸酯还原酶的活性,从而影响叶绿素的合成。同时,Al3+还可与Mg2+竞争根表质膜上载体的结合点与吸收点,从而抑制植物对Mg2+的吸收,降低植物体内Mg2+的含量来影响植物叶片叶绿素总含量。铝胁迫下,油菜叶绿素荧光参数随着铝浓度的增加而降低。常绿杨的铝胁迫实验发现,植株的光合效率,生长量和生物量都受到影响,并且观察到叶肉细胞超微结构损伤程度与叶绿素荧光参数和植株生长量趋势一致。烟草在铝胁迫作用下,叶片的叶绿素含量和净光合速率等荧光参数显著降低。可见铝胁迫对植物叶片的光合作用产生了一定的抑制,光合参数则在一定程度上反映出铝胁迫对植物体的损伤情况。本实验选取蚕豆作为实验材料,观察不同浓度Al3+胁迫下对蚕豆幼苗生长发育的影响,研究叶绿素荧光参数及叶绿素含量变化,探讨Al3+对蚕豆光合作用影响的各个可能的因素及其能量耗散途径,为进一步了解Al3+胁迫对蚕豆生理机制提供参考。
2 实验方法
2.1 蚕豆幼苗的培养及Al3+胁迫
选取大小均一饱满的蚕豆300粒,自来水浸泡2 d,待种子露白后,置于黑暗条件下生根。根长约2~3 cm 时用蒸馏水培养,并移至阳光充足的地方生长。待蚕豆幼苗的第一对叶片完全展开,第二对叶片刚生长时,移入配制的不同浓度Al3+(10 μM、50 μM、100 μM)的Hoagland营养液中生长,Al3+以硫酸铝(Al2(SO4)3·化学纯)的形式提供,对照组幼苗生长在Hoagland营养液。用NaOH和HCl调节溶液pH值,使其控制在5.5。每盆20株,营养液用气泵持续通气。每天观察处理期间蚕豆的生长情况,每隔5 d更换处理液。
2.2 光合色素含量的测定(Lichtenthaler比色法 单位:mg/g)
蚕豆经Al3+处理3 d、6 d、9 d后,选取5株蚕豆植株相同位置上的叶片(第二对),称量记录鲜重,研磨,丙酮浸提。次日早上进行离心(4000×g 20 min),测定其吸光度A663、A646 ,A470,根据以下公式计算叶绿素a,叶绿素b含量。叶绿素a、b含量的测定方法参照张志良主编《植物生理学实验指导方法》方法进行测定。①叶绿素a含量(mg/g)=2×10×10-3×(12.21×A663-2.81×A646)/FW;②叶绿素b含量(mg/g)=2×10×10-3×(20.13×A646-5.03×A663)/FW。
2.3 叶绿素荧光的测定
蚕豆经Al3+处理3 d、 6 d、 9 d后,用LI-COR 公司生产的LI-6400 配套的荧光叶室,测定蚕豆相同叶位(第二对叶片)的叶绿素荧光参数,初始荧光(Fo),最大荧光(Fm),光系统Ⅱ(PS Ⅱ)潜在光化学效率(Fv/Fm),开放的PS II反应中心的激发能捕获效率(Fv′/ Fm′),PS Ⅱ实际的电子传递的量子效率(ETR),非光化学猝灭参数(qN)。每组测5株幼苗。
2.4 数据统计方法
采用Sigmaplot 8.02软件处理所有数据,统计并作图,选用平均值和标准方差进行统计分析,使用 SPSS 13.0 对数据进行差异性分析。
3 结果与分析
3.1 Al3+胁迫对蚕豆株高和根长的影响
实验过程中,对照组幼苗株高生长良好,茎杆粗壮,叶片呈绿色。如图1所示,Al3+处理3 d后,Al3+处理组幼苗株高生长与对照组幼苗株高没有明显差异。但随着Al3+处理时间的延长,幼苗株高生长逐渐受到抑制。10 μM和50 μM Al3+处理6 d 后,两组幼苗株高低于对照组,但差异不显著;而100 μM Al3+处理组幼苗株高显著低于对照组(P<0.05)。Al3+处理9 d后,10 μM、50 μM和100 μM Al3+处理组的幼苗株高均显著低于对照组(P<0.05),各Al3+处理组之间的幼苗株高没有显著差异。因此,Al3+对蚕豆幼苗生长的抑制随时间延长和浓度的增加,其抑制作用越明显。但不同Al3+处理组幼苗的叶片均没有出现毒害症状。
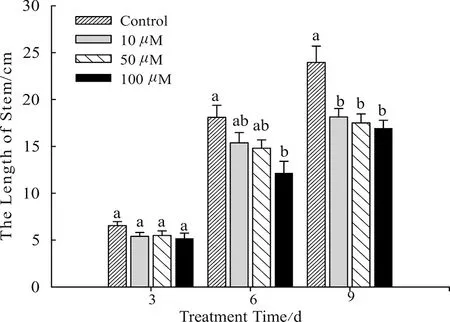
图1 不同浓度Al3+处理对蚕豆株高的影响(n=10)
整个实验过程中,对照组根生长良好,主根粗壮,侧根发达,根系繁茂呈白色。与对照组相比,10 μM Al3+处理组的幼苗侧根生长减少,根系颜色呈微褐色;随着处理时间的延长,6 d后根生长与对照组差异显著。在50 μM Al3+处理组中,根长及生长量低于对照组,根系颜色呈棕色,根尖膨大,有螺纹状裂纹。100 μM Al3+处理组主、侧根短且易断,卷曲呈鱼钩状,侧根稀少,根系呈黑褐色,根螺纹现象加重。
图2显示了不同Al3+浓度处理对蚕豆根生长的影响。10 μM Al3+处理6 d后,其根生长与对照组根长相比有显著差异(P<0.05)。50 μM 和100 μM Al3+处理组的根生长,在实验过程中均显著低于对照组。在Al3+处理的3个组合中,随着Al3+浓度的升高,对根生长的抑制作用越明显,其中100 μM Al3+处理组幼苗根生长量最少。
3.2 Al3+胁迫对叶绿素荧光参数的影响
Fv/Fm表示PSⅡ最大光化学量子产量,反映PSⅡ反应中心的最大光能转换效率,通常在叶暗适应20 min后测得。整个实验中,对照组及铝处理组叶片的Fv/Fm值无显著性差异(P>0.05)。各Al3+处理组与对照组叶片的Fv/Fm值在0.80~0.83之间,属正常范围(表1)。

图2 不同浓度Al3+处理对蚕豆根长的影响(n=10)
Fv′/Fm′是PSⅡ实际光化学效率,它反映开放的PSⅡ反应中心原初光能捕获效率。100 μM Al3+处理3 d的蚕豆幼苗叶片中Fv′/Fm′值降低,与对照组及10 μM、50 μM处理组相比差异显著(P<0.05)。6 d后,尽管各Al3+处理组与对照组的Fv′/Fm′值无显著差异(P>0.05),但Al3+处理组Fv′/Fm′值低于对照组。50 μM 和100 μM Al3+处理9 d后,幼苗叶片中的Fv′/Fm′值比对照组显著降低(P<0.05)。
ΦPSⅡ为光合电子传递量子效率,表示PSⅡ实际光化学量子产量,它反映PSⅡ反应中心在有部分关闭情况下的实际原初光能捕获效率。ETR表示表观光合电子传递速率,与光合作用过程中光反应电子传递有关。实验中,不同浓度Al3+处理后蚕豆幼苗叶片ΦPSⅡ和ETR参数的变化趋势一致。10 μM 处理组与对照相比无明显差异(P>0.05)。50 μM Al3+处理6 d后,幼苗叶片的ΦPSⅡ和ETR值下降,与对照组相比差异显著(P<0.05)。实验过程中,100 μM Al3+处理的幼苗叶片,ΦPSⅡ和ETR值始终显著低于对照(P<0.05)。
qN代表非光化学淬灭,反映的是PSⅡ天线色素吸收的光能不能用于光合电子传递而以热的形式耗散掉的光能部分。表1显示了Al3+对蚕豆幼苗叶片的qN影响中,各处理组的非光化学淬灭系数qN都呈增长趋势。相同处理时间内,蚕豆幼苗叶片的qN随Al3+浓度增加而上升。在100 μM Al3+处理下,qN值显著高于对照组(P<0.05)。50 μM Al3+处理3~6 d的叶片与对照组相比,qN值显著增加(P<0.05)。10 μM 处理组与对照相比无明显差异(P>0.05)。
3.3 Al3+胁迫对叶绿素含量的影响
Al3+胁迫下,蚕豆幼苗叶片光合色素含量的变化如表2所示。结果表明,实验过程中,各Al3+处理组的叶绿素a(Chla)、叶绿素b(Chlb)含量以及叶绿素a+b含量与对照组相比无明显差异(P>0.05)。
从表2可以看出,50 μM和100 μM Al3+处理3 d后的蚕豆幼苗叶片Chla/b与对照组相比,显著降低(P<0.05)。实验中Al3+处理6 d后,3个处理组与对照相比,其蚕豆幼苗叶片Chla/b值下降,但无明显差异(P>0.05)。处理9 d后100 μM Al3+处理组Chla/b明显下降(P<0.05)。据以上结果分析,Al3+处理使蚕豆幼苗叶片光合色素含量下降,高浓度Al3+(100 μM)处理组Chla/b明显降低。

表1 不同浓度Al3+对蚕豆叶片叶绿素荧光参数的影响
注:不同字母表示差异显著,相同字母表示差异不显著(P<0.05),平均值±标准误差,n=3
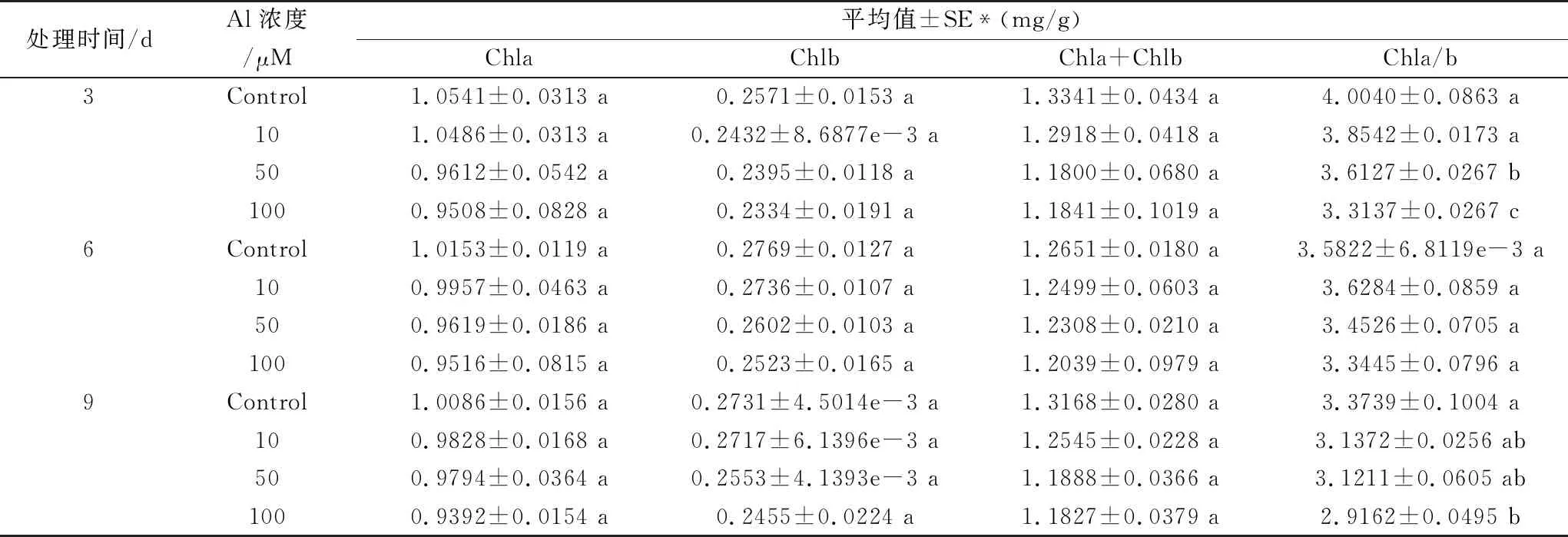
表2 不同浓度Al3+处理对蚕豆叶片光合色素含量的影响
注:不同字母表示差异显著,相同字母表示差异不显著(P<0.05),平均值±标准误差,n=3
4 讨论
4.1 Al3+胁迫对蚕豆幼苗生长的影响
本实验结果表明,不同浓度Al3+对蚕豆幼苗生长发育有明显的影响,其中,最明显的毒害特征是抑制株高和根生长。结果显示,Al3+胁迫蚕豆幼苗9 d 后,10 μM和50 μM Al3+处理组的株高显著低于对照组;在整个实验过程中,100 μM Al3+处理下蚕豆植株株高生长受到明显抑制。在根中,10 μM Al3+处理6 d后,幼苗根生长才受到抑制,而50 μM和100 μM Al3+胁迫下,植株根长受到明显抑制。铝抑制植物幼苗生长的结果与多数学者研究结果一致。一般认为,由于根系直接接触铝溶液,因此最先受到毒害。水稻在铝胁迫下,地上部分和根的鲜重显著下降。增加Al3+浓度和延长处理时间,小麦幼苗地上部分的高度趋于下降,小麦根生长受到显著抑制。植物根系是吸收和积累铝的主要器官。松苗根部的铝含量约是地上部分的4~9倍,转移到地上部分的铝很少。酸性条件下增加Al3+浓度和处理时间,植物根短粗,根尖膨大,弯曲成钩状,根系体积下降,根部细胞结构与功能受到破坏,影响植物水分吸收,营养元素的吸收运输也受抑制,使幼苗生长受到阻碍。我们的结果显示,Al3+胁迫对植物叶片形态和叶片颜色影响不明显,叶绿素含量变化没有差异,说明铝主要积累在植物根部,向地上部分器官转运较少,因而对植物茎叶的毒害较轻。
4.2 Al3+胁迫对叶绿素荧光参数和叶绿素含量的影响
叶绿素荧光参数可以反映叶片光合功能内在特征。相对于无胁迫条件,植物长期处于不良环境中,会出现叶绿素含量降低,光合作用速率下降等症状。随着不利条件出现,荧光参数变化可作为外界胁迫及不利因素征兆的指示值。实验结果中,不同浓度Al3+处理对蚕豆幼苗叶片Fv/Fm比值影响不大,且各处理组与对照组无显著性差异。但是,Fv/Fm比值的测得相对滞后于胁迫引起的变化,同时Fv/Fm 不能显示PSⅡ中活性中心与非活性中心的反应信息。因而Al3+胁迫下蚕豆PSⅡ的影响还需分析其他参数。
实验结果中10 μM和50 μM Al3+下蚕豆幼苗Fv′/Fm′、ΦPSⅡ和ETR参数变化不大;100 μM Al3+处理的Fv′/Fm′、ΦPSⅡ和ETR参数均显著降低,而qN值上升。这表明高浓度Al3+胁迫对蚕豆叶片PSⅡ的光能转换效率、实际光化学效率及电子传递效率影响明显。叶绿素作为光合作用中最重要的色素,在光合作用中发挥吸收、传递和转化光能的重要作用,其含量高低与光合作用密切相关。叶绿素含量结果显示,100 μM Al3+对蚕豆光合色素Chl a/Chl b比值降低,表明高浓度Al3+胁迫导致蚕豆幼苗叶片光合色素捕获及转化光能的能力下降,不利于光合作用的正常进行。植物叶片PSⅡ原初光能转换效率降低,可能是叶片PSⅡ活性中心受损,光合作用原初反应过程受抑制。100 μM Al3+胁迫下蚕豆的ETR参数下降表明光合电子传递受阻导致,而PS II反应中心天线色素吸收的过量光能通过热能耗散,因而非光化学淬灭系数qN上升。所以非光化学荧光碎灭是一种自我保护机制,对光合机构起一定的保护作用。
猜你喜欢
小读者(2021年2期)2021-03-29
少儿科学周刊·少年版(2021年17期)2021-01-17
阅读(科学探秘)(2020年8期)2020-11-06
阅读(低年级)(2020年4期)2020-06-21
绿色科技(2019年2期)2019-05-21
小资CHIC!ELEGANCE(2018年28期)2018-09-14
学生天地(2018年9期)2018-09-06
女性天地(2016年10期)2017-04-25
分析化学(2016年12期)2017-02-04
人生与伴侣·共同关注(2016年24期)2016-08-23
