
东北地区人参锈腐病菌的分子鉴定
2019-07-08 09:46:38傅俊范刘欣然李自博周如军韩月泠
沈阳农业大学学报 2019年3期
傅俊范,刘欣然,李自博,周如军,王 丹,韩月泠
(沈阳农业大学 植物保护学院,沈阳110161)
人参(Panax ginseng C.A. Meyer)为五加科人参属多年生草本药用植物[1],具有极高的药用和经济价值。 我国人参主产区集中在东北地区的辽宁、吉林和黑龙江三省。 近年来,随着农业生态种植结构调整,国家对森林资源的保护日益加强,农田平地栽参面积不断增大,逐渐取代传统的林地栽参,成为我国人参产业重点发展方向。由柱孢属真菌[Cylindrocarpon destructans (Zinns.) Scholten]引起的人参锈腐病近年来在东北人参产区发病严重,且危害随参龄增加逐年加重,直接影响人参产量和商品价值,限制了人参产业的发展[2]。 人参锈腐病菌可侵染人参的主根、侧根、须根及芽胞等部位,罹病人参植株地上部矮小且叶片变黄或变红,人参根部呈红锈色椭圆形或其他不规则形状。 发病严重时,锈腐病斑汇合成片,人参表皮遭到严重破坏,病菌侵入根内组织呈干腐状,最终枯萎死亡。据统计,目前在我国人参主产区锈腐病重病地块,发病率高达70%~80%[3]。 由于人参锈腐病菌具有潜伏侵染的特性,可在土壤中长期存活或在病残体中越冬,作为次年病害的初侵染源[4],因此该病在连作参地中发生更为严重,也是导致人参连作障碍的主要病害之一[5]。
严雪瑞等[6]基于传统的形态学比较分析,将长白山区分离得到的12 株锈腐病菌划分为4 个致病力不同的类群,即毁灭柱孢(Cylindrocarpon destructans),人参柱孢(Cylindrocarpon panacis),钝柱孢(Cylindrocarpon obtusisporum)和人参生柱孢(Cylindrocarpon panicicola),其中毁灭柱孢(C. destructans)和人参柱孢(C. panacis)致病力相对较强,钝柱孢(C. obtusisporum)和人参生柱孢(C. panicicola)致病力相对较弱。 由于真菌无性阶段形态相近,表型形态鉴定困难,rDNA-ITS (internal transcribed spacer,ITS)以其信息位点丰富、序列长度保守等特点已被广泛应用于真菌种间及种内分类分型研究当中[7]。 SONG 等[8]利用rDNA-ITS 和mt SSU rDNA 两种序列分析方法对来自韩国不同地区的15 株锈腐病菌进行了种内鉴定, 并根据其致病力差异, 将其分为两个不同的类群。 近年来,人参锈腐病菌的相关报道主要集中在病原菌的形态鉴定,生物学分析及药剂筛选等方面,分子生物学研究相对较少,尤其是利用分子手段确定东北人参主产区人参锈腐病菌的种类及致病力差异的研究尚未见报道。
本研究旨在真菌分类学基础上[9-10],通过rDNA-ITS 序列分析明确来自东北人参主产区的45 株锈腐病菌的主要种类,并探究种类间形态及致病力的差异,为人参锈腐病的综合防控提供理论基础。
1 材料与方法
1.1 材料
供试3年生人参购买自辽宁省抚顺市清原满族自治县大苏河乡。 分别从辽宁省(抚顺市新宾县红升乡、丹东市宽甸县太平哨镇、本溪市桓仁县华莱镇和抚顺市清原县大苏河乡)、吉林省(集安市榆林镇、白山市松江河镇、集安市台上镇、通化市西江镇和集安市太王乡)和黑龙江省(虎林市虎林县和哈尔滨市清河县)人参主产区共11 块参田的3年生人参罹病植株上分离纯化病原菌。
供试PDA 培养基为马铃薯200g,琼脂17g,葡萄糖20g,蒸馏水1000mL。试验采用的试剂及仪器:新型植物基因组 DNA 提取试剂盒(天根,北京)、DNA Marker-C(生工,上海)和 2×Es Taq MasterMix(Dye)(康为,北京),PCR 仪(ABI,美国)、通用电泳仪(ABI,美国)、凝胶成像分析系统(UVP,美国)和生物显微镜(NIKON,北京)。
1.2 方法
1.2.1 人参锈腐病菌分离 采用五点取样法,在11 块参田,每块随机选取5 个小区,每个小区采集5~10 株病参用于病原菌的分离。 采用改良的方中达[11]组织分离法,切取感病参根病健交界处3mm×3mm 表皮,经75%乙醇表面消毒10s 后,置于2%次氯酸钠水溶液中消毒60s,无菌水冲洗3 次,置于PDA 平板上,3 次重复。 20℃黑暗培养3~5d 后长出白色菌落,采用单孢分离法对病原菌进行纯化。
1.2.2 病菌rDNA-ITS 序列分析 将供试的45 株人参锈腐病菌取直径5mm 菌饼置于铺有玻璃纸的PDA 培养基上,20℃黑暗培养15d, 刮取玻璃纸表面菌丝, 放置于1.5mL 的离心管中, 采用试剂盒提取法提取菌丝DNA,-20℃保存备用。 选择真菌通用引物ITS1(5'-TCCGTAGGTGAAC-CTGCGG-3')和ITS4(5'-TCCTCCGCTTATTGATAT-GC-3')进行PCR 扩增。 反应体系和程序参考孙嘉曼[12]的方法。 PCR 产物由上海生工生物工程技术服务有限公司测序,测序结果在 GenBank(http://www.ncbi.nlm.nih.gov/)中进行 BLAST 比对分析,并用MEGA6.0 软件以邻接法构建系统发育树。
1.2.3 病菌形态学观察 将5mm 菌饼置于PDA 平板上,20℃黑暗培养11d 后观察病原菌的培养性状,包括菌落直径、菌落颜色及气生菌丝情况。菌落直径采用十字交叉法测定,3 次重复。显微镜检测观察病菌子实体形态并拍照。
1.2.4 病菌致病性测定 采用针刺法对人参进行室内离体接种。 用脱脂棉蘸取75%乙醇对人参根表皮擦拭消毒,待酒精完全蒸发后,用无菌的解剖针对每个供试人参主根刺伤5 点,伤口直径0.25mm,深1.00mm[13],用浓度为每升1×106个孢子的人参锈腐菌孢子悬浮液进行接种,3 次重复,以接无菌水处理组作为对照;将接种后的人参置于直径9cm 的培养皿中,在皿底部铺设蘸有无菌水的纱布,保持湿度为95%~100%。 28d 后调查发病结果。
病害严重度分级标准参考RAHMAN 等[14]的方法:1 为无明显损伤;2 为褐色病斑直径为0.9mm;3 为深褐色病斑直径为1.0~4.0mm;4 为黑色病斑直径为4.0~7.0mm;5 为黑色病斑直径大于7.0mm,病斑相连呈不规则大斑块;6 为整株侵染发病。 并计算病情指数。

1.3 数据分析
试验数据采用SPSS19.0 软件进行单项方差分析,采用Duncan 氏新复极差法进行差异显著性检验p<0.05。
2 结果与分析
2.1 人参锈腐病菌分离结果
采用五点取样法,共采集到150 个人参锈腐病样,分离纯化得到45 株菌。从辽宁省抚顺市新宾县红升乡罹病 人 参 根 部 分 离 纯 化 得 到 15 株 菌 (XB004,XB040,XB008,XB006,XB048,XB047,XB050,XB007,XB042,XB049,XB051,XB011,XB009,XB052,XB046); 从丹东市宽甸县太平哨镇罹病人参根部分离纯化得到 5 株菌(KD018,KD020,KD019,KD014,KD016); 从辽宁省本溪市桓仁县华莱镇罹病人参根部分离纯化得到 2 株菌(HR001,HR002); 从辽宁省抚顺市清原县大苏河乡罹病人参和西洋参根部分离纯化得到10 株菌(R13,R7,R5-4,R5-3,X5,X7,X25-5,X25-4,X25-1,X25-3)。 从吉林省集安市榆林镇罹病人参根部分离纯化得到 2株菌(Yl036,YL026);从吉林省白山市松江河镇罹病人参根部分离纯化得到2 株菌(FS027,FS028);从吉林省集安市台上镇罹病人参根部分离纯化得到2 株菌(JA030,JA031);从吉林省通化市西江镇罹病人参根部分离纯化得到2 株菌(TH029,TH002);从吉林省集安市太王乡罹病人参根部分离纯化得到3 株菌(YC022,YC024,YC025)。 从黑龙江省虎林市虎林县罹病人参根部分离纯化得到1 株菌(HL023);从黑龙江省哈尔滨市清河县罹病人参根部分离纯化得到1 株菌(QH043)。

表1 东北人参主产区45 株人参锈腐病菌的rDNA-ITS 序列分析Table 1 Dendrograms of 45 Cylindrocarpon isolates with rDNA-ITS
2.2 人参锈腐病菌的ITS序列分析
采用ITS1/ITS4 引物对45 个菌株进行PCR 扩增, 均获得约500bp 的核苷酸序列, 将供试菌株的序列与Genbank 中已知种的菌株序列进行BLAST 比对,加入Campylocarpon fasciculare 作为外源参考序列[15-19]构建系统发育树(图 2)。结果表明,XB046 等 28 株病原菌与毁灭柱孢(C. destructans)(AY677271)相似性达到 99%,聚在同一分支;R13 等3 株病原菌与毁灭柱孢毁灭变种(C. destructans var. destructans)(AM419065)相似性达到99%,聚在同一分支;YL026 等 14 株病原菌与强壮土赤壳(I. robusta)(AY295331, JF735264)相似性达到 99%,聚在同一分支(图1,表1)。 因此将所有供试菌株鉴定为3 个种:毁灭柱孢毁灭变种(C. destructans var. destructans),共有 3 个;毁灭柱孢(C. destructans),共有 28 个;强壮土赤壳(I. robusta),共有 14 个。
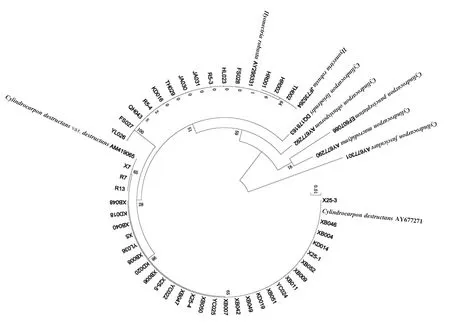
图1 基于rDNA-ITS 序列采用NJ 法建立人参锈腐病菌及其相关病原菌的系统发育树Figure 1 The phylogenetic tree of Cylindrocarpon and other related isolates based on rDNA-ITS sequence using NJ method
2.3 人参锈腐病菌形态学观察
在PDA 培养基上,45 株供试菌株大部分菌落初呈白色或米色,后变棕灰色、浅褐色、深褐色等不同颜色,菌落边缘整齐,多数呈匍匐状,气生菌丝繁茂;毁灭柱孢毁灭变种(C. destructans var. destructans)较其他种相比菌落颜色最深,为深褐色,而强壮土赤壳(I. robusta)颜色最浅,为棕灰色;不同供试菌株间生长速率差异较大,病原菌培养 11d 后菌落直径为(4.21±0.16)~(7.78±0.25)cm,其中菌株 R5-4 生长速度最快,菌落直径为(7.78±0.25)cm,菌株R13 生长速度最慢,菌落直径为(4.21±0.16)cm;3 个种的菌落生长速率差异显著,其中强壮土赤壳(I. robusta)生长速度最快,毁灭柱孢毁灭变种(C. destructans var. destructans)最慢。3 种菌都产生大分生孢子和小分生孢子:大分生孢子圆柱形,直或弯曲,两端钝圆,1 个或多个隔膜;小分生孢子,椭圆形居多,无隔膜(图 2,表 2)。
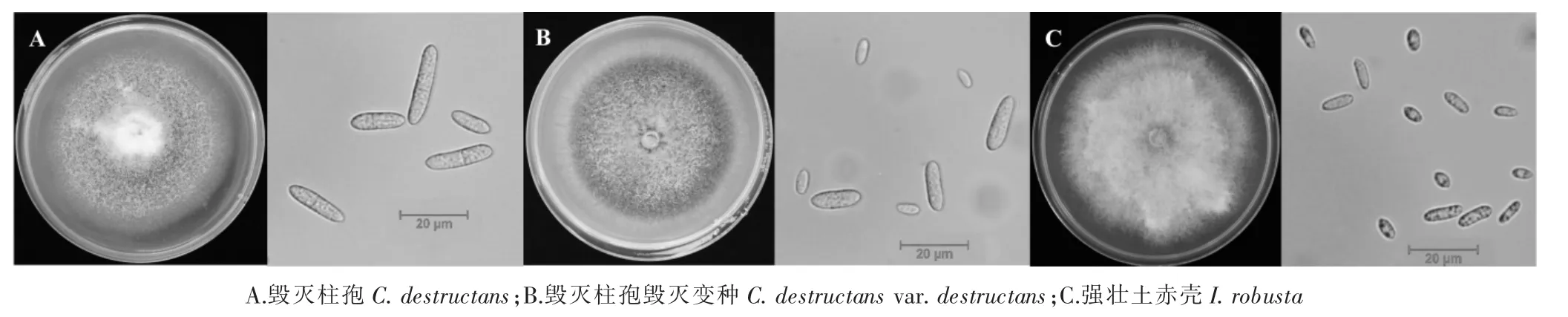
图2 人参锈腐病菌典型菌落形态(左)及分生孢子形态(右)Figure 2 Typical colonies (Left) and macroconidia (Right) of Cylindrocarpon isolates from ginseng root rust rot
2.4 人参锈腐病菌致病力分析
将供试菌株按照不同组别进行离体接种,各组别间表现出不同的致病力,其中毁灭柱孢毁灭变种(C. destructans var.destructans)致病性最强,在接种人参2d 后可见明显黄锈色病斑;其次为毁灭柱孢(C. destructans),接种人参3d 后出现病症, 病斑较小; 强壮土赤壳(I.robusta)致病力相对较弱,供试人参均在接种后5d 出现锈黄色病斑,个别接种点出现轻微腐烂。 随着时间推移,参根表面病斑扩展程度不均,以毁灭柱孢毁灭变种(C. destructans var. destructans)最为严重,致病力最强,其他种均表现出不同程度致病力(图3)。

表2 人参锈腐病菌形态和致病力Table 2 Morphological data and pathogenicity test of Cylindrocarpon isolated from ginseng root rust rot

图3 人参离体接种锈腐病菌发病症状比较Figure 3 The symptoms to ginseng with pricking method
3 讨论与结论
人参锈腐病是人参产业中最为严重的病害之一,尤其在我国东北人参主产区流行成灾,严重制约了我国人参产业健康可持续发展。 1966年戚佩坤[2]首次在中国报道3 种引起吉林地区人参锈腐病的病原菌,即C. destructans,C. panacicola 及 C. sp.;徐中志等[4]报道了 4 种人参锈腐病菌 , 分别 为 C. destructans,C.panacicola 和两个柱孢属未知种病原菌;严雪瑞[6]将长白山区分离得到的人参锈腐病原菌分为4 个种,即C.destructans,C. panacis,C. obtusisporum 和C. panicicola。 以往对人参锈腐病菌的鉴定主要以病原菌的形态特征为依据,该方法易受多种外界因素影响。 近年来,利用特异性PCR、rDNA-ITS 序列分析等分子技术辅助鉴定病原菌种类的方法已得到广泛应用[20],通过将形态学与分子技术结合,可以进一步提高鉴定的准确性[21],如张鸿雁等[15]利用形态学结合分子鉴定的方法明确了引起黑龙江地区人参锈腐病的病原菌为C. destructans。 本研究以CHAVERRI 等[10]的分类方法为基础,结合病菌形态特征和rDNA-ITS 基因序列分析技术,对采集到的45 株人参锈腐病原菌进行了鉴定。鉴定结果表明,引起东北地区人参锈腐病的病原菌有3 种,分别是C. destructans,I. robusta 和C. destructans var. destructans。 在辽宁地区采集分离的32 个菌株中,75%为C. destructans,是该地区的主要致病菌,与前人的研究结果一致[2,4,6];吉林地区为I. robusta 和C. destructans;在黑龙江地区为I. robusta。 其中,在吉林和黑龙江地区采集分离到的病原菌种类与严雪瑞和张鸿雁等的报道存在差异,这可能是因为人参锈腐病是典型的土传性病害,病原菌的种类易受到气候、土壤微生物群落结构及栽培模式等影响[22],由此导致吉林和黑龙江地区人参锈腐病菌的种群结构发生了变化。
人参锈腐病菌的致病性测定结果表明,不同种病原菌存在致病力差异,其中C. destructans var. destructans致病力最强,其次为C. destructans,I. robusta 最弱。 有报道指出,C. destructans 侵染人参后产生的致病酶[13]和毒素[23]是该病原菌重要的致病因子。 EVANS 等[24]从C. destructans 中分离出了一种毒素nectrolide,低浓度的nec-trolide 可以延缓植株生长, 甚至可以使植物幼苗根部变黑, 这可能是C. destructans var. destructans 及C. destructans 致病性较强的原因。
通过将传统病原菌鉴定手段与现代分子鉴定技术相结合的方法,对采自东北人参主产区45 株人参锈腐病原菌的主要种类进行划分,结果为C. destructans, I. robusta 和C. destructans var. destructans 3 种。 本研究结果初步揭示了东北地区人参锈腐病原菌的种群结构组成, 为进一步研究人参锈腐病菌致病力分化机制及群落遗传进化关系奠定基础,也可为该病害的综合防控提供理论依据。
