
内蒙古赛罕乌拉国家级自然保护区中华斑羚行为节律初步研究
2019-07-08 02:16杨婧陈照娟乌力吉王晓玲穆丽光向昌林刘波陈琛鲍伟东
四川动物 2019年3期
杨婧,陈照娟,乌力吉,王晓玲,穆丽光,向昌林,刘波,陈琛,鲍伟东*
(1. 北京林业大学生物科学与技术学院,北京100083; 2. 内蒙古赛罕乌拉国家级自然保护区管理局,内蒙古大板025150)
由于受人类活动影响,野生动物栖息地日趋破碎化,种群数量和分布范围持续下降(Pettorellietal.,2010)。自然保护区类似于被人类社区包围的生物多样性孤岛,生活于此类片段化环境中的野生动物必须实时调整行为策略,才能较好地利用有限资源以保证种群长期稳定(Bocedietal.,2014)。动物行为是动物对环境变化做出的最直接反应,环境条件中的物理、生物因素以及动物自身的生理状况决定动物的行为类型及适应对策(Abáigaretal.,2018)。动物的活动节律是行为生态学中的一个重要内容,行为的时间分配和节律可以反映动物个体的营养状态、社会地位以及生存压力等,了解这些生活史特征,对于制定濒危物种的保护措施至关重要(Rowcliffeetal.,2015)。
红外相机作为非损伤性的物种调查和记录手段,于20世纪90年代开始被应用于野生动物研究(Karanth & Nichols,1998)。与传统监测方法相比,红外相机技术具有对动物干扰小(Schipper,2007;Mendozaetal.,2011)、能捕获难以发现的物种(Carboneetal.,2001;Trolle & Kéry,2003;刘雪华等,2018)、影像资料便于存档检索等优点(O’Connelletal.,2011),成为野生动物调查监测与行为节律研究的有力工具。近年来,国内相关研究和保护机构开始广泛使用红外相机,如对野生白冠长尾雉Syrmaticusreevesii的活动节律和时间分配的研究(赵玉泽等,2013)和对浙江凤阳山-百山祖国家级自然保护区内兽类和鸟类多样性进行的红外相机调查(李佳等,2018),分析红外相机获得资料建立了四川卧龙国家级自然保护区野生水鹿Rusaunicolor的行为谱(张晋东等,2018)。
中华斑羚Naemorhedusgriseus为国家Ⅱ级重点保护野生动物,被世界自然保护联盟(IUCN)评估为易危(VU),同时被列入《濒危野生动植物种国际贸易公约(CITES)》附录Ⅰ(王应祥,2003;吴建平等,2005;蒋志刚等,2016)。中华斑羚生性机警,主要生活于山顶多岩石地带,野外条件下很难近距离观察,有关该物种野外行为的观察报道较少。对其近缘物种的行为研究也局限于圈养条件下的观察(刘振生等,2005),缺乏自然状态下的行为特征分析。与圈养条件不同,动物在野外状态下被捕食的风险明显提高,捕食风险是动物做出行为决策时需要权衡的一个重要因子(边疆晖,樊乃昌,1997)。动物正常活动时,依据信息源的变化确定环境中的捕食风险,并依据自身条件权衡捕食风险,通过空间选择、活动节律和栖息地利用等行为的变化降低捕食风险(魏万红等,2004)。本研究选取内蒙古赛罕乌拉国家级自然保护区一个孤立分布的中华斑羚种群,在捕食者出现明显恢复的背景下,研究中华斑羚行为节律的季节变化对栖息地破碎化和捕食风险的适应,对该保护区内中华斑羚的生态状况进行初步评估,为保护区管理者制定相应的保护措施提供理论依据。
1 研究地区与研究方法
1.1 研究地区
内蒙古赛罕乌拉国家级自然保护区位于内蒙古自治区赤峰市巴林右旗北部,地理坐标为118°18′~118°55′E,43°59′~44°27′N,总面积100 446 hm2,位于中温带半湿润温寒气候区,年均气温2 ℃,7月最热,平均气温18 ℃,1月最冷,平均气温-18 ℃(李桂林,2005)。现有哺乳动物38种,隶属于6目14科31属。保护区内中华斑羚的主要捕食者为欧亚猞猁Lynxlynx,张帅(2014)通过分析红外相机影像资料和活动痕迹估测其在保护区的数量至少为8只。
1.2 研究方法
2015年9月15日—10月6日,每天从日出到日落约9 h不间断地在山脚下架设高倍单筒望远镜(SWAROVSKI ATX,25~60×80),直接搜寻目标区域内出现的中华斑羚个体,并观察记录行为类型、持续时长、发现地点等。由于发现个体较少,于2016年5—12月,在中华斑羚分布最集中的栖息地青羊城至五间房后沟一带布设10台红外相机(LTL-5210A)进行24 h不间断连续拍摄以获取个体行为(图1),相机设置为20 s录像,5 s间隔,每月定期更换所有相机的电池及内存卡。
红外相机布设于常年有中华斑羚活动的地区(周许伟,2015),从而避免由于相机数量较少引起拍摄的行为内容不足,有效影像总时长为292 min 20 s(表1)。然而,由于雌雄性中华斑羚均长角,没有显著的性别区分特征,野外自然条件下难以开展不同性别之间的行为比较。另外,本研究获得影像中幼年个体比例少,所以仅对成年个体的行为进行了初步分析。
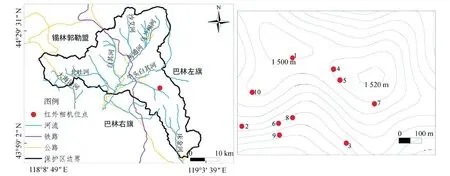
图1 研究区域及红外相机布设位点Fig. 1 Survey area and camera trap sites
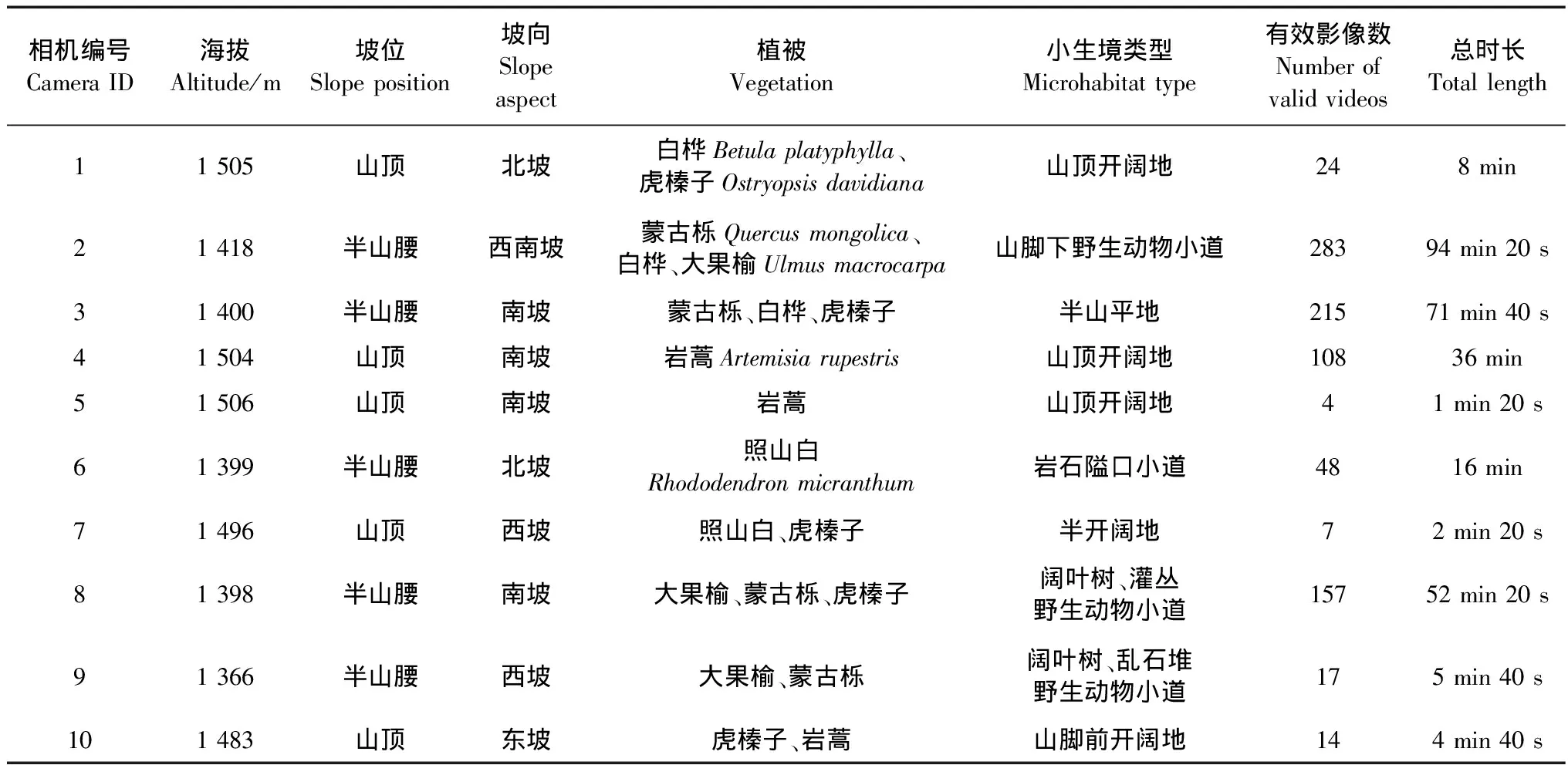
表1 红外相机布设位点小生境信息Table 1 Microhabitat information of camera trap sites
中华斑羚的行为模式分类尚无参考,本研究借鉴圈养条件下赤斑羚Naemorhedusbaileyi的行为模式(谢燕,2006;郑建清,2011;侯晓云,2014;王爱善等,2014),并结合观察到的行为特点加以优化,构建野外自然条件下中华斑羚的行为谱。采用Excel进行数据汇总,分析中华斑羚在观察期内各类行为发生的比例,以月为单位对数据进行分析。
2 研究结果
2.1 中华斑羚活动频次及持续时长
单筒望远镜直接观察共记录到中华斑羚7次,各种行为持续时间共计133 min 40 s;红外相机共拍摄到中华斑羚1 526次,各种行为持续时间共计234 min 46 s(表2)。
2.2 中华斑羚行为谱
野外观察记录中华斑羚的行为类型主要包括站立、卧息、移动、摄食、修饰、发情和其他行为(表3)。
2.3 中华斑羚行为类型比例分配
根据行为谱将各时间段内中华斑羚的行为分类并进行分析。摄食行为和站立行为在所有月份中均有出现,移动行为除2015年9—10月未观察到外,其他月份中均有出现。2015年9、10月单筒望远镜观察记录到的数据显示,摄食与卧息行为属于主要行为类型。2016年5—12月红外相机监测数据显示,摄食、站立、移动行为是中华斑羚的主要行为。摄食行为中,2016年11月比例(79.51%)最高,2015年10月比例(15.48%)最低;站立行为中,2016年8月比例(31.27%)最高,2016年11月比例(5.25%)最低;移动行为中,2016年12月比例(18.3%)最高,2016年8月比例(5.56%)最低;发情行为仅在2016年10月、12月零星出现,是占比最低的行为类型(图2)。
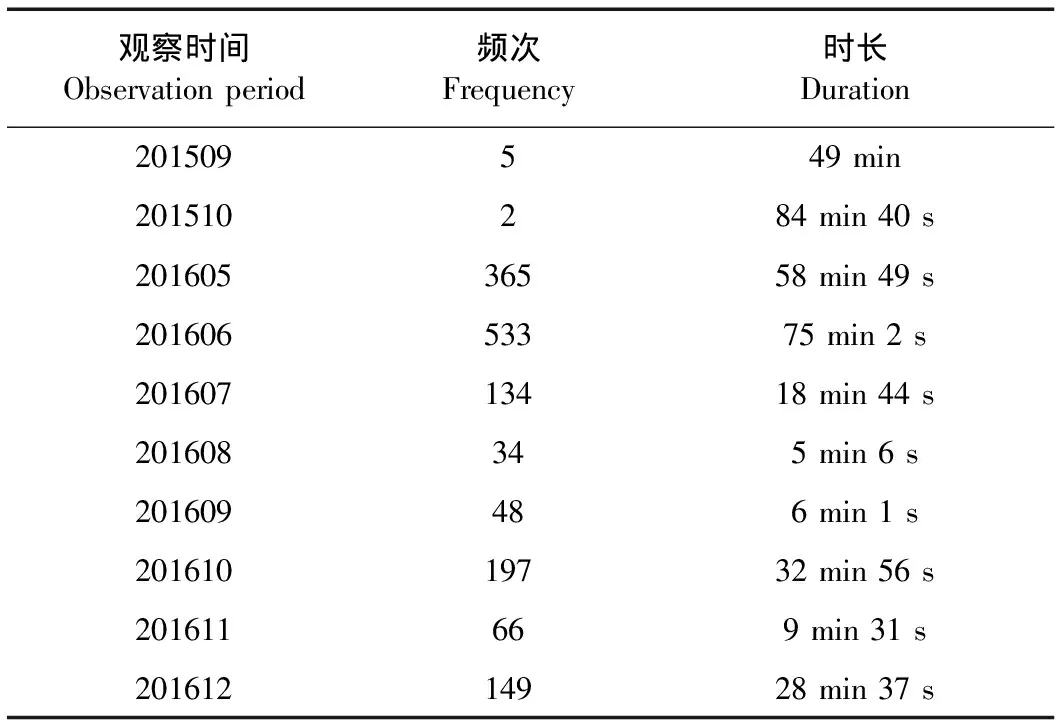
表2 中华斑羚活动频次及持续时长Table 2 Frequency and duration ofNaemorhedus griseus activities

表3 中华斑羚行为谱及描述Table 3 Behavior ethogram of Naemorhedus griseus

图2 中华斑羚行为类型比例分布
Fig. 2 The proportion distribution ofNaemorhedusgriseusbehavior types
3 讨论
本研究对中华斑羚活动节律的观察发现,2015年9—10月采用单筒望远镜远距离观察,未能记录到反刍等细微的行为,行为类型较单一。后期借助红外相机获得了较丰富的行为类型,可见研究手段的改进有助于获得更加细致的数据资料。
3.1 中华斑羚行为特点
动物的行为时间分配可反映食物的可得性、环境条件及行为学特征(March,1981),动物的相对活动强度与红外相机拍摄概率正相关,红外相机在某时间段拍摄的几率越高,意味在此时间段内动物活动强度也越高(Lietal.,2010)。中华斑羚摄食、移动行为的月度变化与气温及食物资源的季节性差异相关,8月食物充足,动物无需高强度采食,且多在阴坡处乘凉,活动强度小;秋、冬季降温和植物干枯导致食物资源匮乏,动物需加大摄食强度获得能量,活动时间延长,表现为典型的季节适应特点。中华斑羚的站立、卧息行为常常伴随反刍行为发生,这与以往对大角羊Oviscanadensis的研究相同(Pelletier & Festa,2004),因此,可用卧息及站立行为的时间表达反刍行为的发生时间(连新明等,2012)。发情行为仅在2016年10月、12月偶有出现,中华斑羚的发情行为出现频次较少可能是野外条件下红外相机无法做到对其发情行为过程的全记录,且口吻部与生殖区接触与日常行为中的嗅尾极为类似,因而无法准确判定。本研究记录到发情行为,说明采用的红外相机方法具有一定可行性,后期将通过布放更多相机来获取交配行为的影像资料予以确认。
3.2 中华斑羚与圈养赤斑羚行为比较
由于国内外对于中华斑羚行为的研究欠缺,有关该物种的自然行为特点无从考证,只能参考近缘物种的相关研究。郑建清(2011)对动物园繁殖场10只圈养赤斑羚的行为时间分配和活动规律研究发现,赤斑羚大部分时间处于趴卧和采食,站立行为次之,运动和修饰行为较少,且采食行为和站立行为受到饲养员喂食和打扫卫生过程的影响。与之相比,自然状态下中华斑羚的摄食行为比例最高,取食时间不固定,站立和移动行为次之,卧息行为较少,这些差异与动物的生活状态息息相关,这与天山盘羊Oviskarelini行为的研究结果一致(郭松涛等,2003)。自然状态下的中华斑羚生活空间大、食物来源相对匮乏,需要不断运动取食,趴卧时间少。孙立新和盛和林(1989)对动物园5头圈养喜马拉雅斑羚Naemorhedusgoral观察发现,其排粪场所固定,排粪前有嗅地行为,排尿时偶尔也有嗅地行为,排尿场所不固定。在自然状态下,中华斑羚活动区域大,排粪、排尿场所不固定。这些结果显示,了解动物自然状态下的行为表达,有助于对保护区生态评估提供有效依据,也为养殖管理、丰容建设提供参考资料。
3.3 中华斑羚警戒行为对捕食风险的适应
捕食风险是捕食者对猎物的威胁和限制,猎物对捕食风险的判断是形成行为决策的基础(路纪琪,张知彬,2004),也是评估猎物对捕食者适应能力的体现(魏万红等,2004)。研究表明,警戒行为是野生动物重要的反捕食策略之一,主要表现为抬头凝视或环视周围环境(李忠秋,2016),影响捕食者和猎物视觉能力的地表覆盖物及捕食者的存在和出现等,都显著影响猎物摄食活动时间和对食物资源的摄取率(Lathametal.,2018),外界的捕食风险越高,动物用于警戒行为的时间比例也越高(Bednekoff & Lima,2004)。在本研究区域内,中华斑羚的天敌是欧亚猞猁,通过核查红外相机影像,发现中华斑羚的站立、卧息和摄食等行为常常伴随警戒行为,8月草木繁茂,一定程度上阻挡了中华斑羚的视野,警戒次数明显增加;11月草木凋零,中华斑羚在山顶较高区域视野比较开阔,较少的警戒次数即可满足需要。此外,中华斑羚生性机警,栖息地常处于裸岩周围,表明保护区内的中华斑羚已对该区域生态条件及捕食风险产生了有效适应,形成了相应的反捕食机制。本研究对于深入了解野生动物对捕食风险的适应行为具有重要参考价值。
猜你喜欢
环球时报(2022-05-23)2022-05-23
金桥(2021年4期)2021-05-21
当代水产(2021年1期)2021-03-19
四川蚕业(2021年1期)2021-02-12
小学生优秀作文(趣味阅读)(2020年3期)2020-06-04
当代水产(2019年9期)2019-10-08
江西农业学报(2019年8期)2019-09-04
电子制作(2019年7期)2019-04-25
电子制作(2019年7期)2019-04-25
小学生作文(中高年级适用)(2018年5期)2018-06-11
