
一批新育成小麦品种(系)主要农艺性状分析及其分子标记鉴定
2019-06-14 07:14许小宛姚俭昕高雅洁张玲丽孙道杰
麦类作物学报 2019年5期
许小宛,姚俭昕,高雅洁,冯 毅,张玲丽,孙道杰
(西北农林科技大学农学院,陕西杨凌 712100)
小麦作为我国的主要口粮作物之一,其产量高低直接关系到我国的粮食安全。黄淮麦区是我国冬小麦生产的重要区域,在我国的小麦生产中占有极其重要的地位[1]。近年来,黄淮麦区育成并推广了大批优良品种,为我国的小麦育种提供了大量优异种质资源,也为我国的粮食生产做出了重大贡献。对黄淮麦区新育成小麦品种(系)进行分析评价,能够客观、全面地了解本地区的育种现状及存在问题,为新品种的推广及后续品种的选育提供理论参考。
前人关于小麦品种(系)的评价多集中在农艺性状、品质性状和抗性上[2-5],将分子标记鉴定与农艺性状相结合进行品种(系)的综合评价的研究甚少。要燕杰等[2]通过对96份小麦材料的农艺性状和品质性状进行多元评价分析,筛选出了综合农艺性状和综合品质性状优良的品种。薛 香等[4]通过对小麦的品质性状进行主成分分析,将13个品质性状浓缩成5个主成分,为小麦的品质改良提供了重要参考。孙彩铃等[5]通过对山东省区试小麦品种(系)的品质性状进行主成分分析和聚类分析,筛选出了6个综合品质较好的小麦品种(系),同时证明了主成分分析可以有效用于小麦品种(系)的综合评价。在分子标记辅助选择上,前人开发了大量分子标记可以有效用于小麦多个性状的选择。Ellis等[6]设计出了两组特异性引物,能够准确鉴别矮杆基因Rht1和Rht2的a、b两种等位变异。Korzun[7]和Worland[8]等的研究表明,SSR标记Xgwm261的扩增结果(192 bp)与系谱分析相结合,可以准确鉴定矮秆基因Rht8。Sun等[9]基于小麦PPO基因序列所设计的STS标记PPO18,能够有效检测多酚氧化酶基因PPO-A1。Zhang等[10]开发出Ppd-P11及Ppd-P12两个光周期标记,可区分光周期敏感型a和不敏感型b两种等位变异。Vp1B3是Yang等[11]基于Vp1B基因开发的一个STS标记,能够有效鉴定小麦品种穗发芽抗性,共有a(652 bp)、b(845 bp)、c(569 bp)三种等位变异,其中a类型与感穗发芽有关,b与c类型与抗穗发芽有关。此外,Kulwal等[12]在3AL上检测到一个与穗发芽抗性有关的位点Qphs.ccsu-3A.1,该位点与标记Xgwm155紧密连锁,具有A、B、C、D四种等位变异。在小麦的赤霉病鉴定方面,朱展望等[13]开发出一对Fhb1诊断性标记PFT-CAPS和His-InDel,其中PFT-Ⅰ/HIS-Ⅰ为抗赤霉病单倍型。
小麦农艺性状是由多个指标构成的复杂性状,通过主成分分析既可以达到降维的效果,又能保证所获取信息的完整性,大大简化了数据分析程序。聚类分析是一种依据遗传距离对材料进行分类的研究方法,在小麦农艺性状及品质性状的研究上应用广泛,已有大量研究证明了聚类分析在对材料分类处理时具有很高的可行性[14-15]。聚类分析可以作为主成分分析的补充,在主成分分析的基础上进行聚类分析有助于提高研究工作的准确性。在品种的选育过程中,仅通过肉眼观察,很难对品质性状和抗性等一些重要性状做到准确的把握,分子标记辅助选择技术(MAS)能够对育种材料进行基因型选择,有效克服在表型观测上的诸多不便,在作物育种中已经得到了广泛应用[16]。本研究拟从农艺性状选择和分子标记辅助选择两方面入手,对黄淮麦区2016-2017年度参加国家区试的81份小麦品种(系)及本课题组育成的10份小麦品种(系)做以全面评价,以期为新品种的推广及后续品种的选育提供理论参考。
1 材料与方法
1.1 材 料
供试材料(表1)为81份2016-2017年度国家黄淮南片区试品种(系)和10份由本课题组育成的小麦品种(系)。
所有材料均于2016年10月份种植于陕西杨凌区试试验站和西北农林科技大学南阳小麦试验站,实验采用随机区组设计,3次重复,各小区中每个品种种植3行,行长2 m,行距25 cm,株距3 cm。常规田间管理。
1.2 方 法
1.2.1 农艺性状调查
性状调查按照国家小麦良种区域试验统一记载标准,在大田记载株高、旗叶长、宽、穗长、小穗数、穗颈长、芒长等性状,室内考种记录穗粒数、籽粒长、宽、千粒重等性状。株高、穗粒数和千粒重数据来源于《2016-2017年度国家冬麦区黄淮南片水地品种试验总结》。
1.2.2 整穗发芽率(GP)的测定
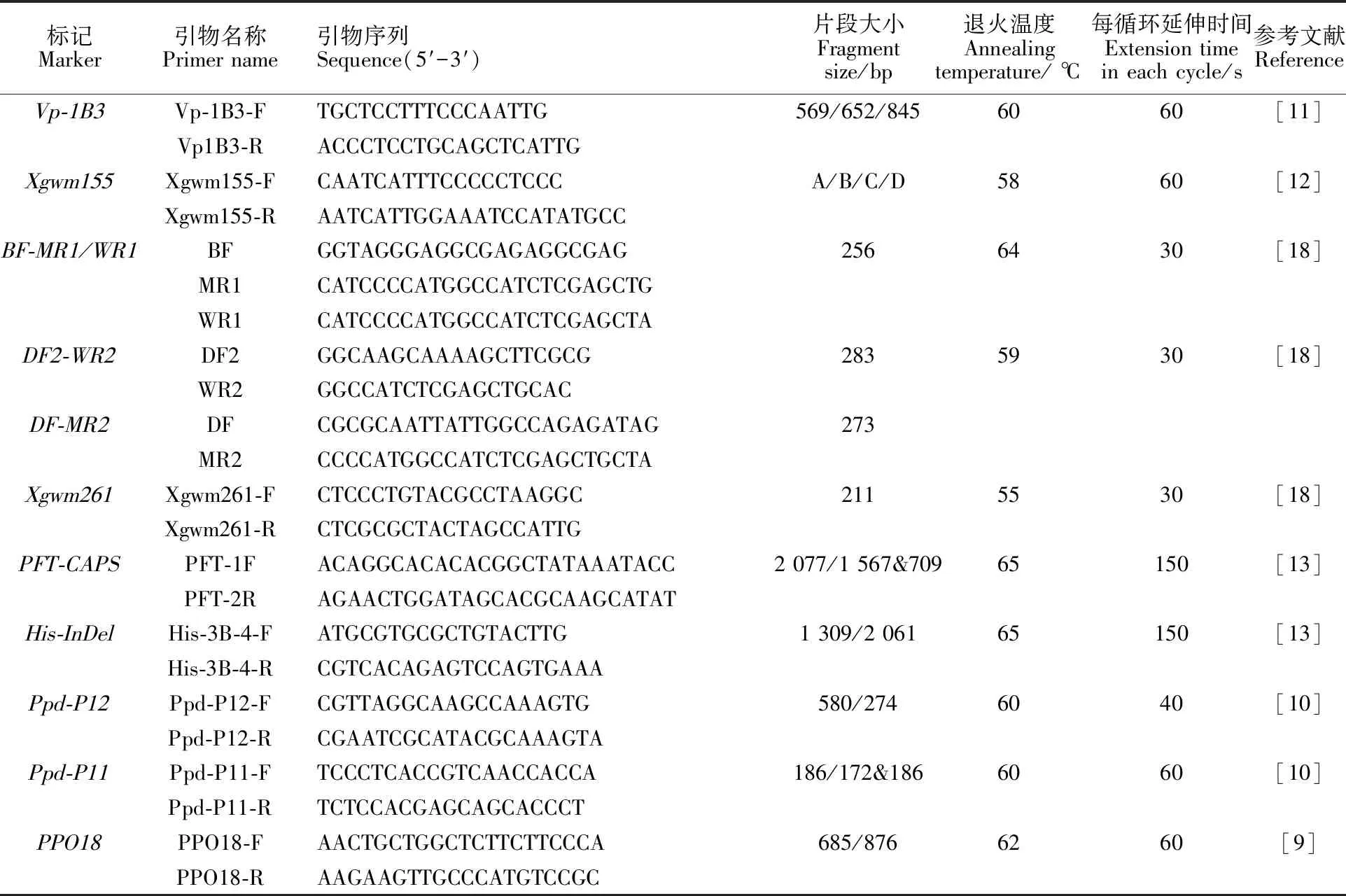
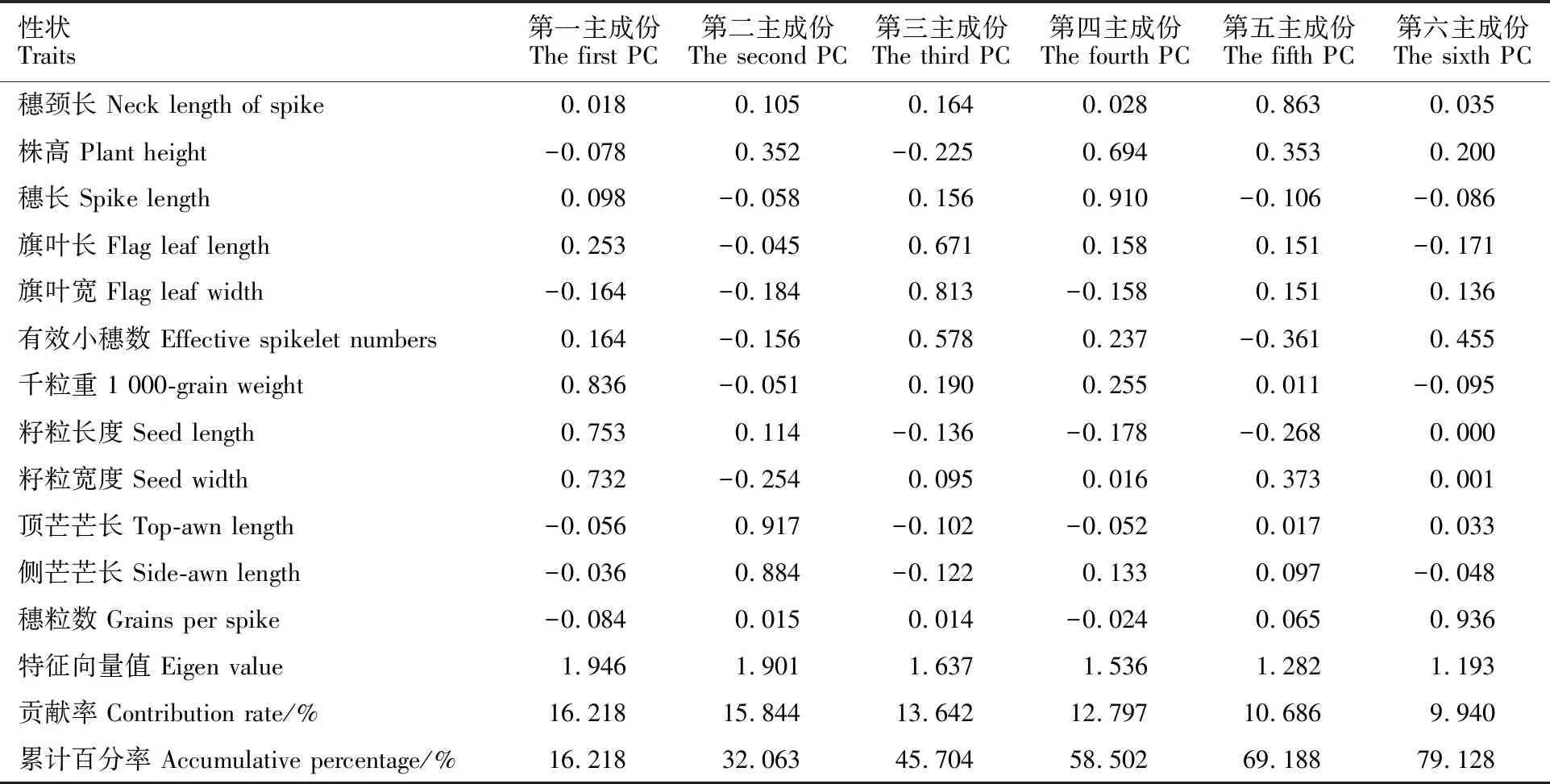
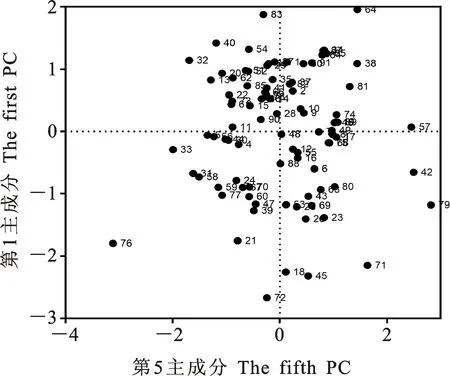
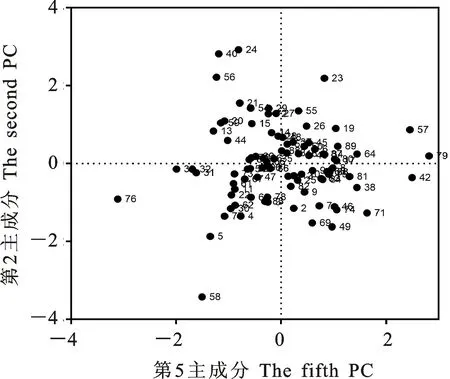
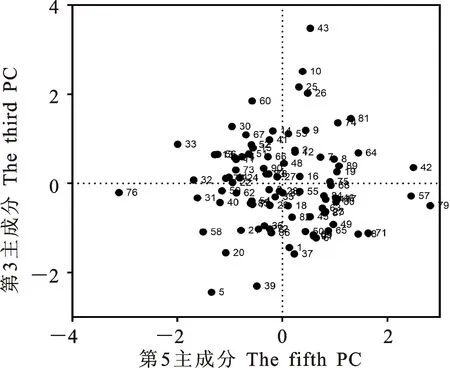
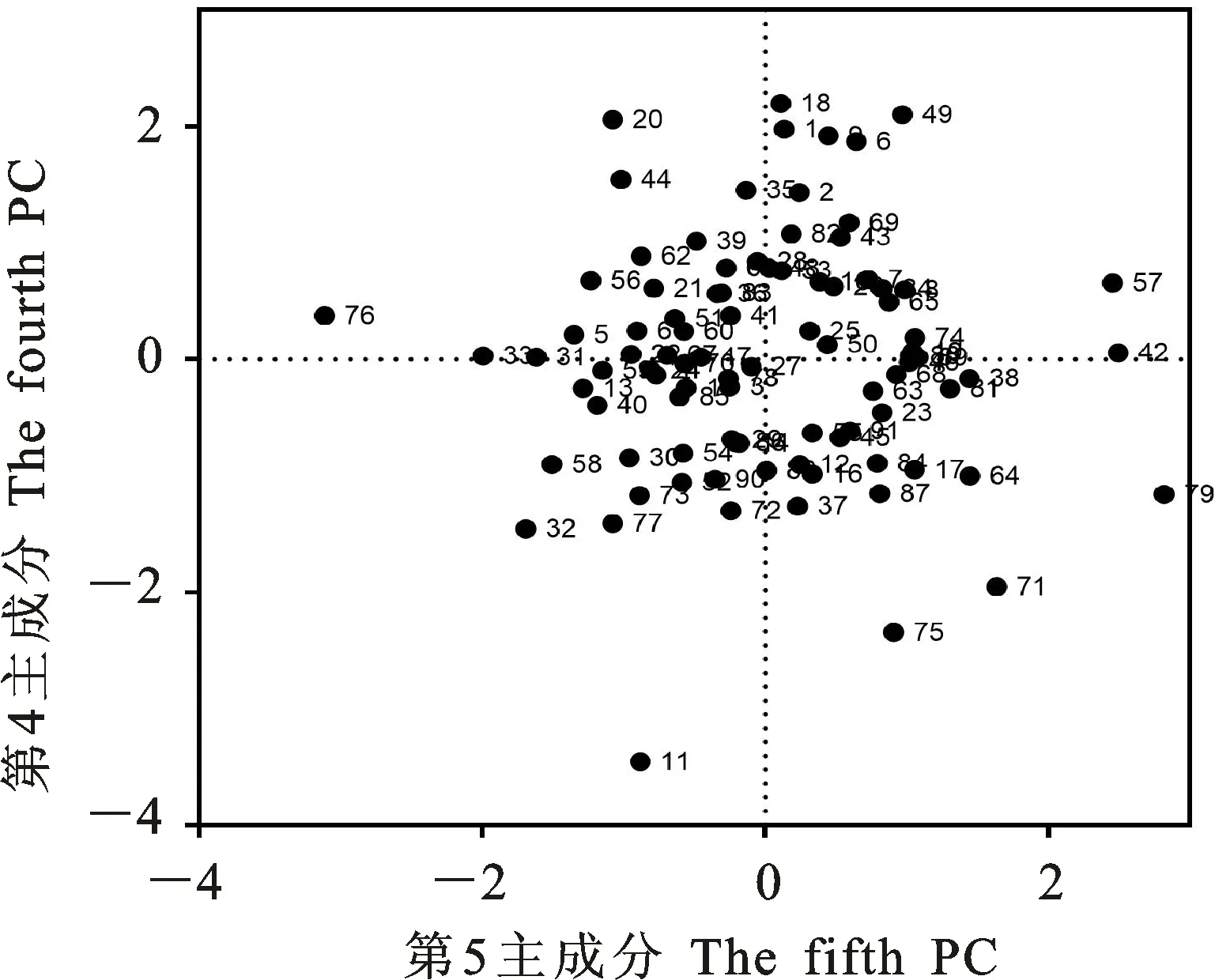
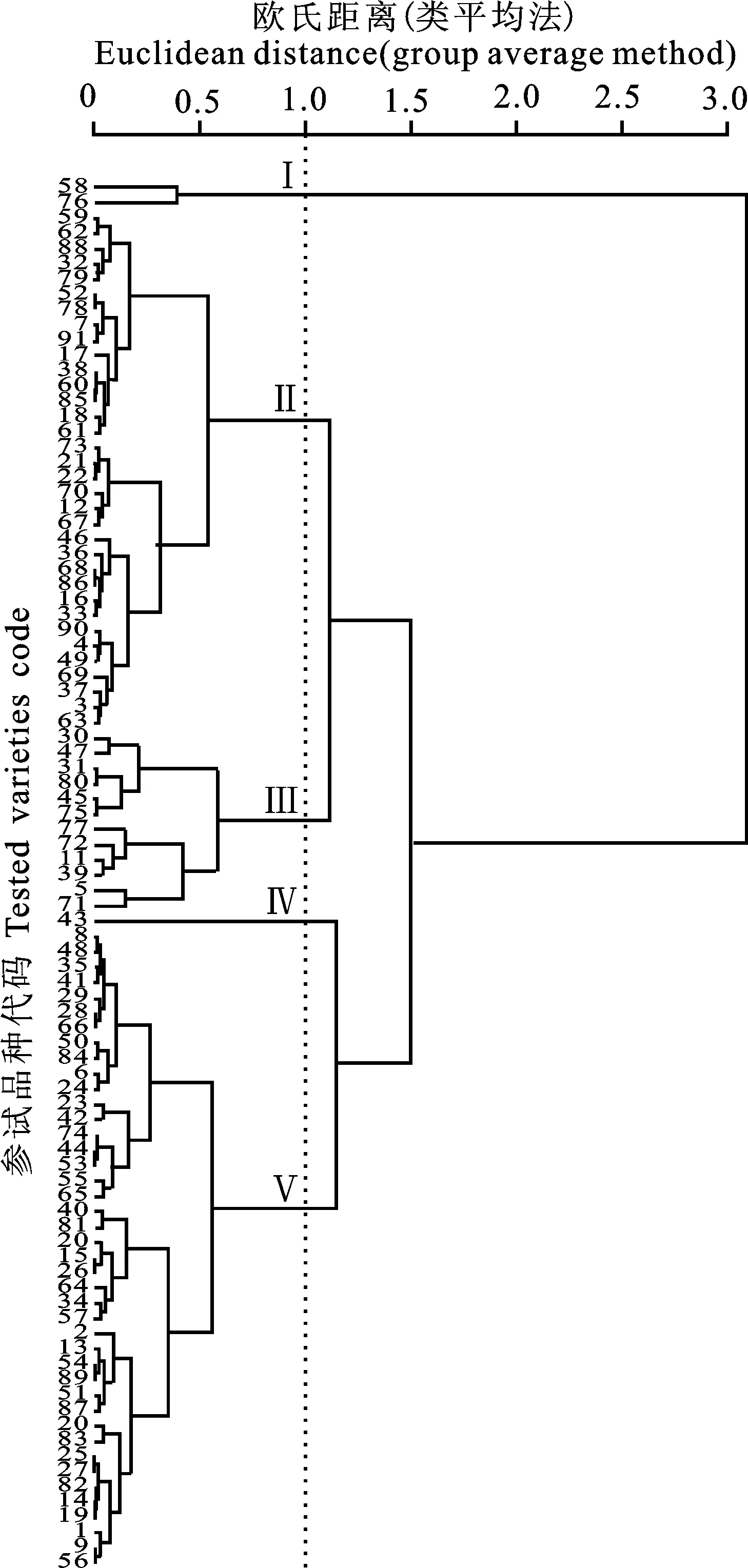
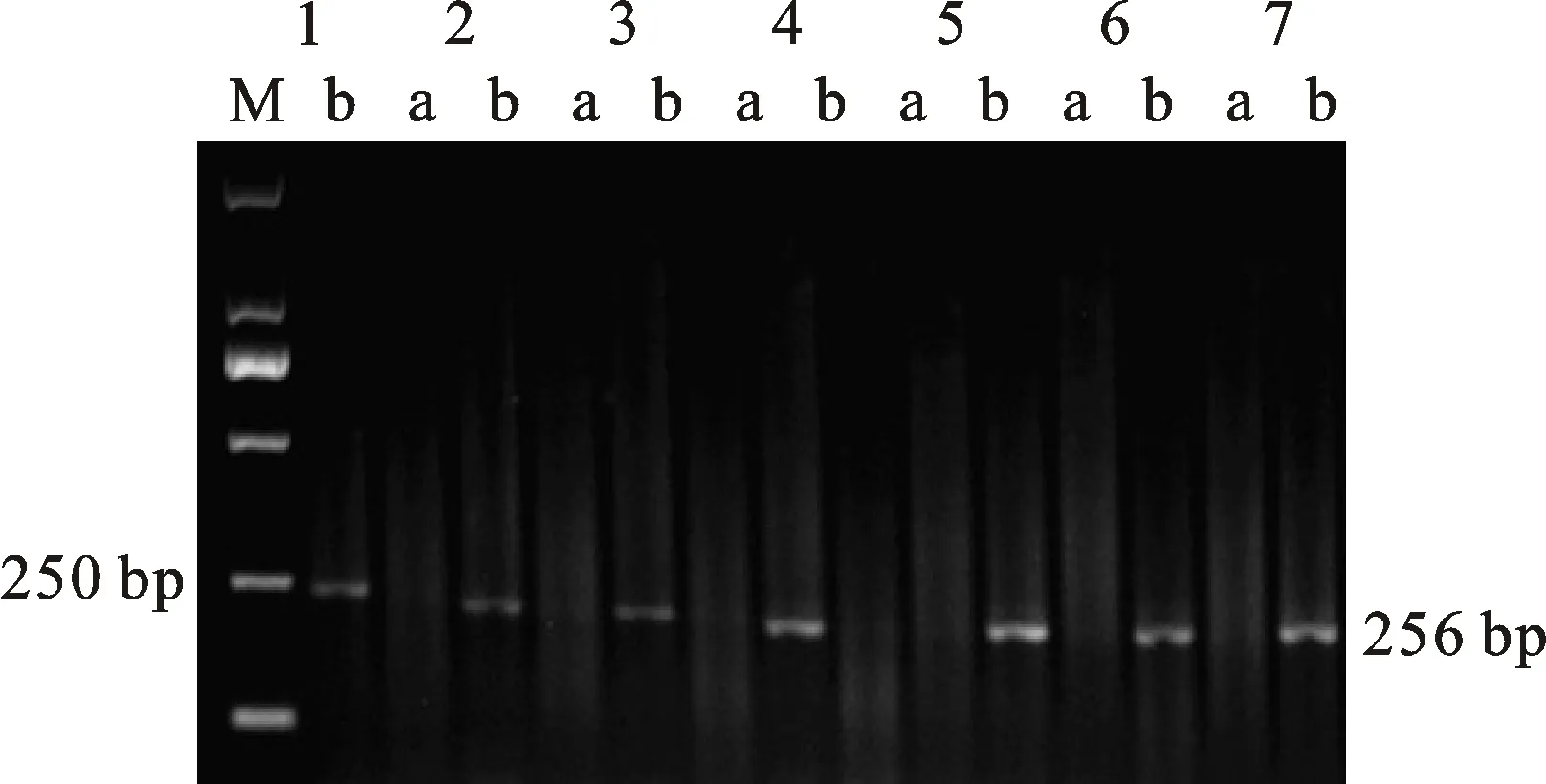
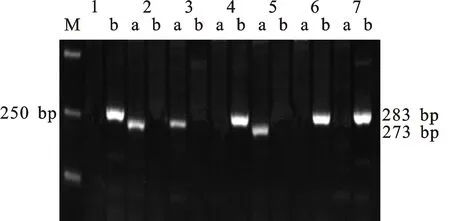
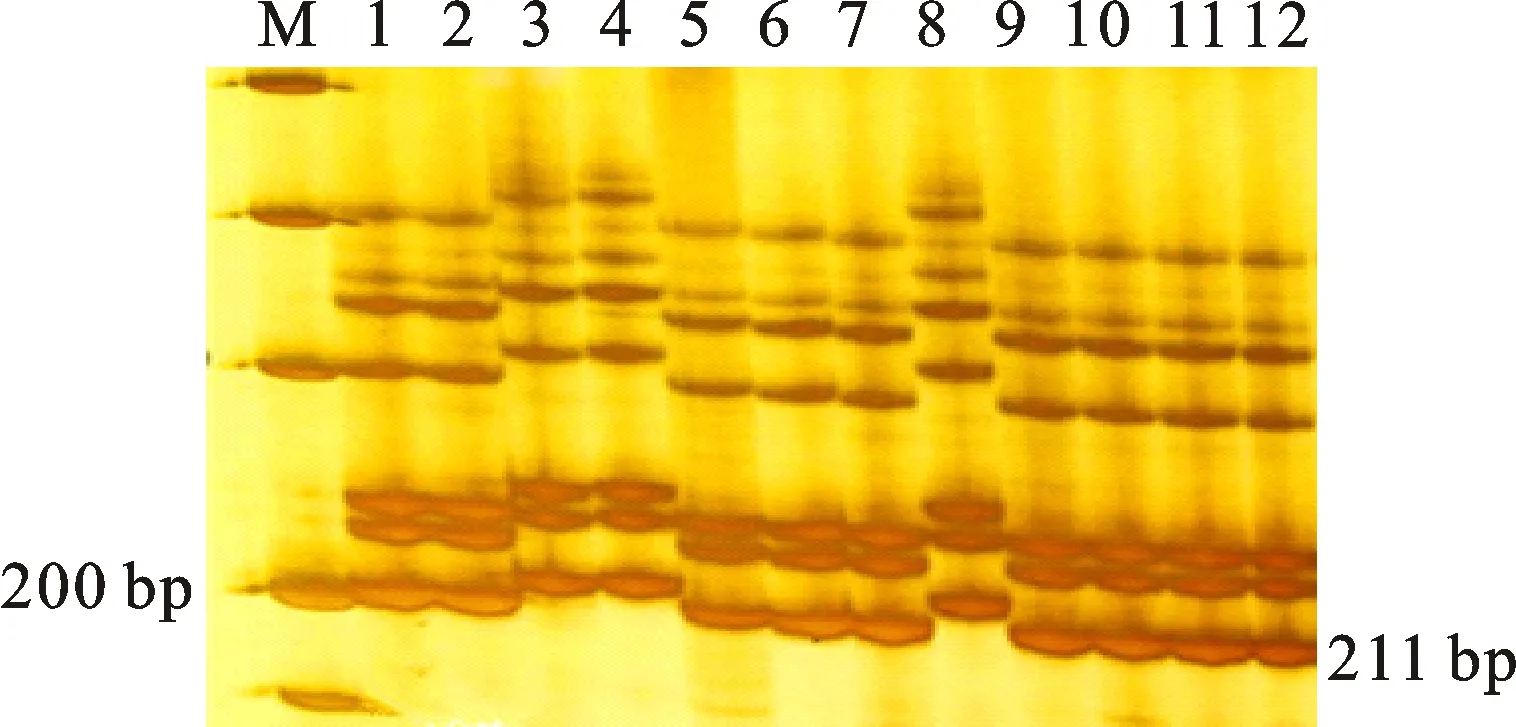
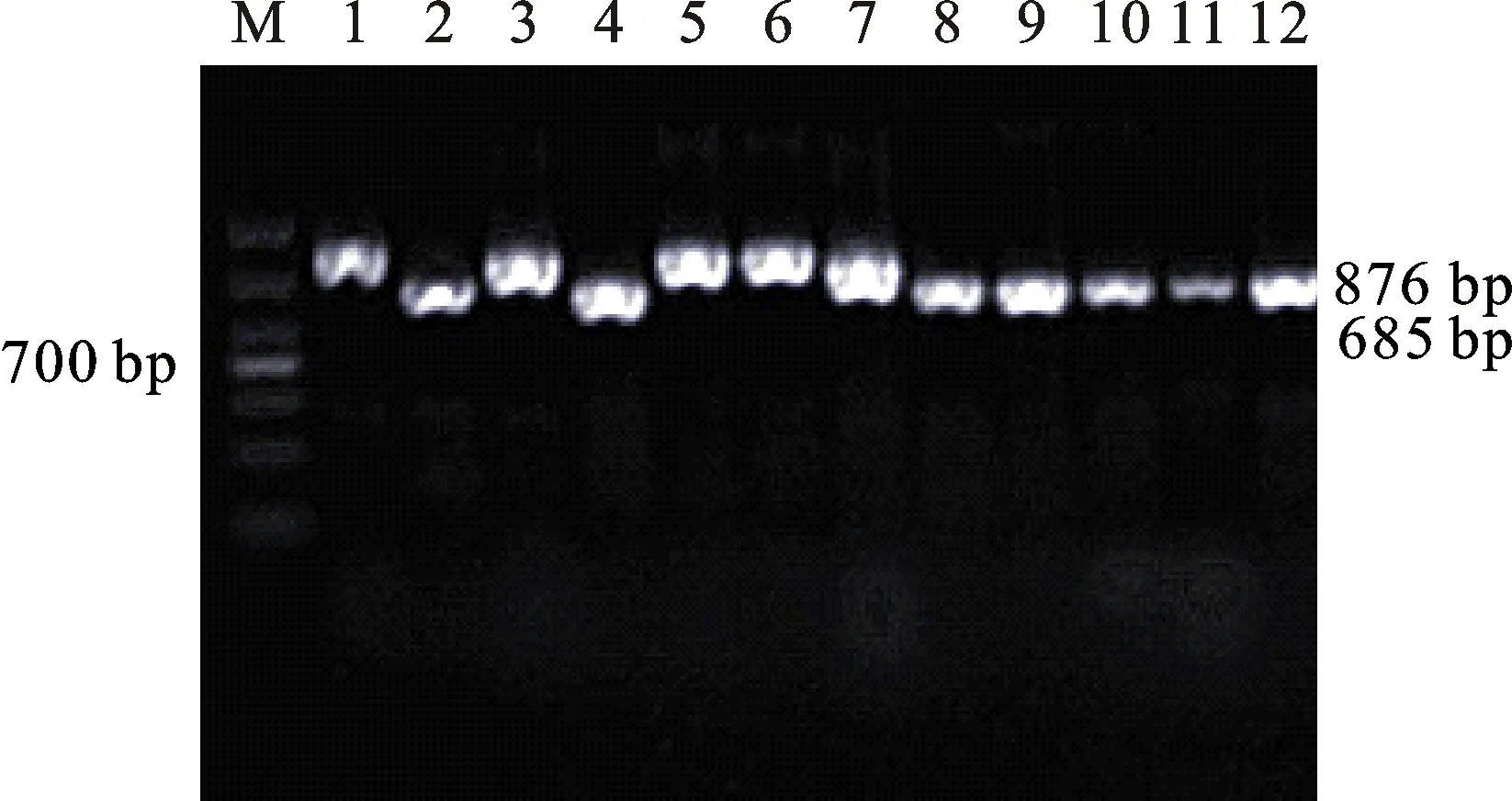
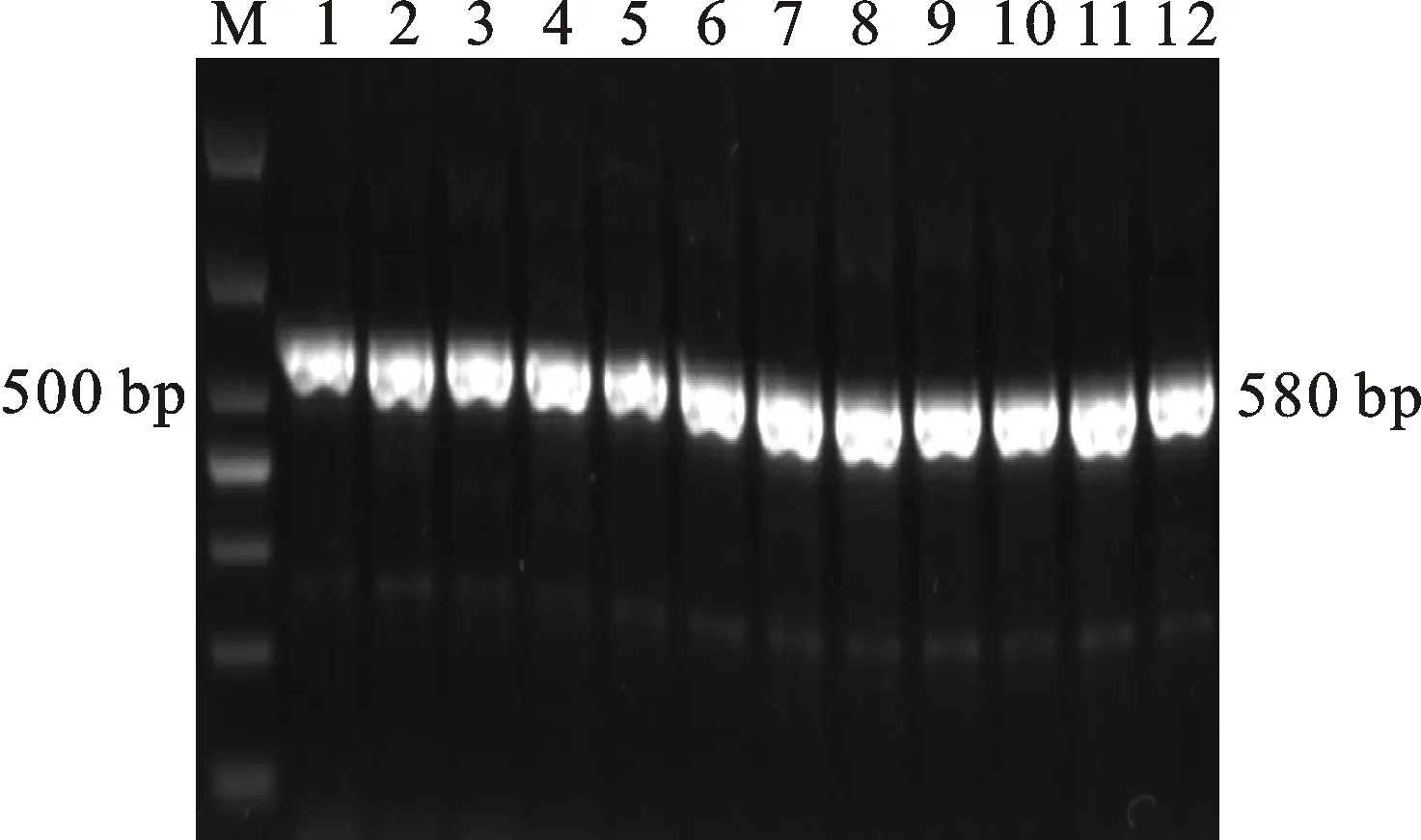
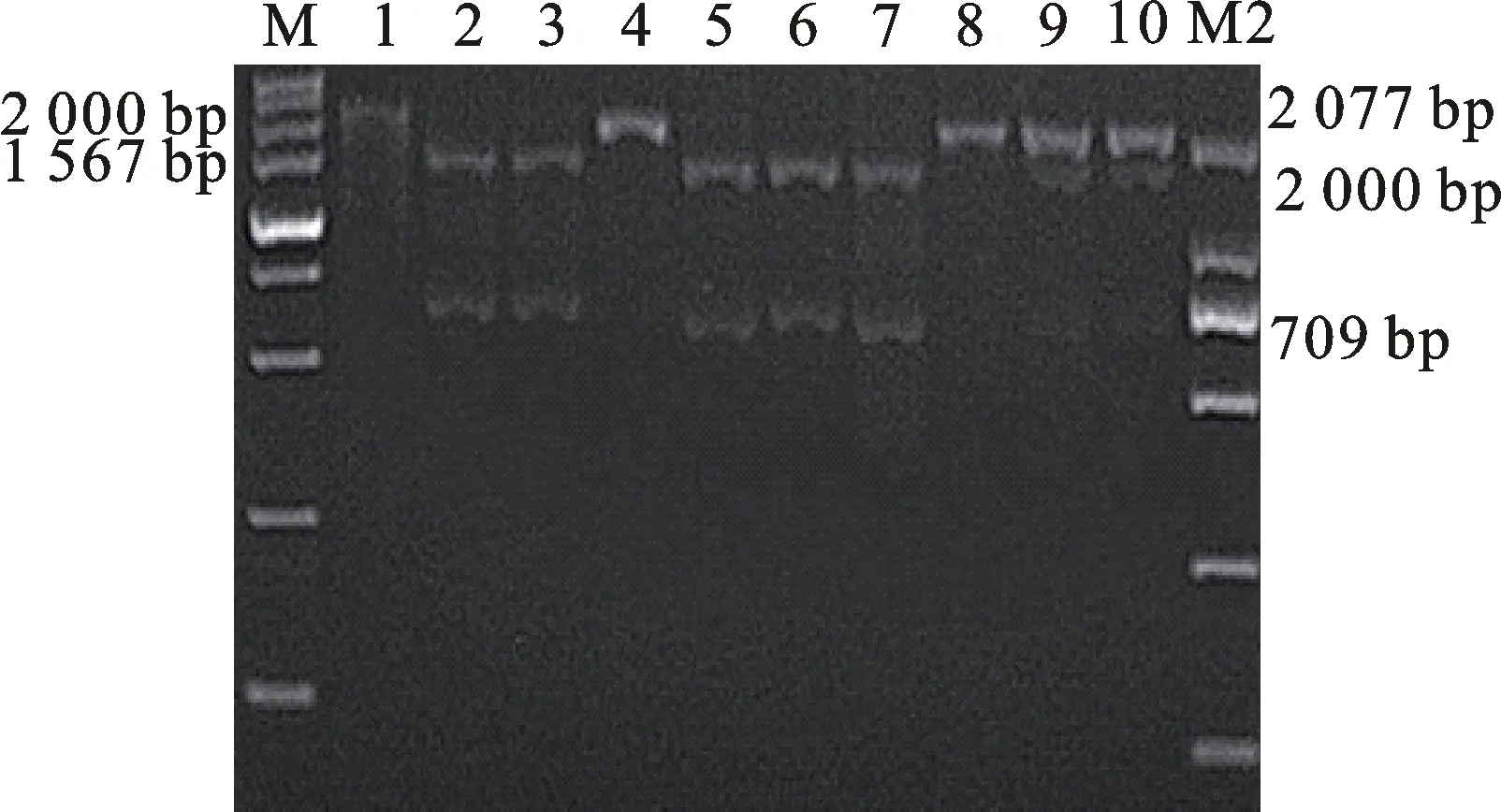
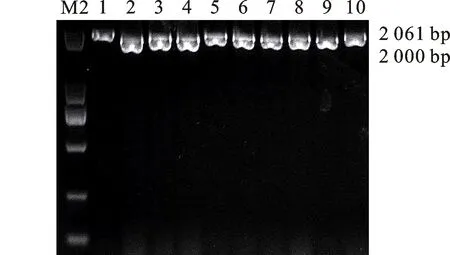
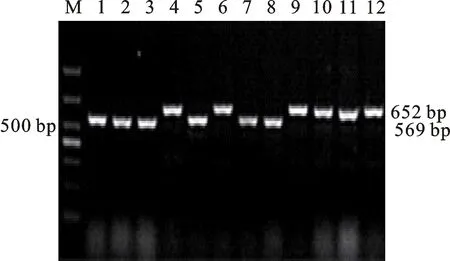
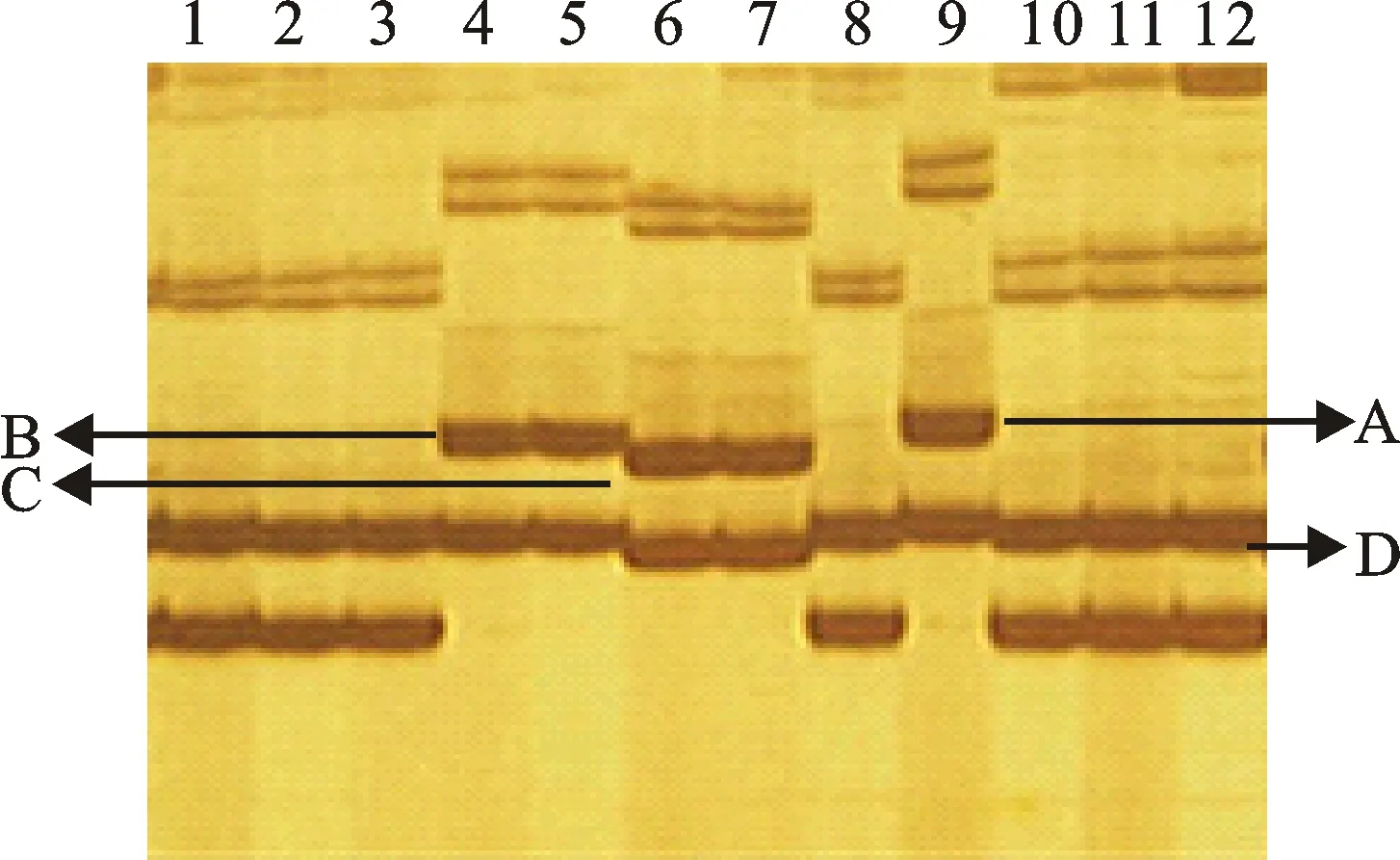
本研究用整穗发芽率(GP)来评价小麦的穗发芽抗性,参照Yang[11]等的分类标准稍作改进,将材料的穗发芽抗性分为高抗(GP≤20%)、抗(20% 表1 供试小麦品种(系)名称Table 1 Wheat cultivars(lines) tested in 2016-2017 1~81:2016-2017年度国家黄淮南片区试小麦品种(系);82~91:本课题组育成的品种或品系。 1-81:Wheat cultivars originated from Huang-Huai wheat growing region which participated in the National Adaptability Test of Wheat Varieties in 2016-2017; 82-91:Cuitivars or lines developed by our research group. 每个品种(系)取主茎穗2个进行编号、挂牌,而后分别置于不同的纸杯中,于纸杯中加水至整穗完全浸没,10 h后将水倒出,每隔4 h喷水1次,确保整穗完全湿润。重复5次。7 d后统计整穗发芽率(GP),以芽长达到籽粒长度视为发芽,反之则为不发芽,最终发芽率为两个穗子发芽率的平均值。 1.2.3 DNA提取及PCR扩增 用CTAB法从幼苗中提取小麦基因组DNA[17]。10对分子标记分别用于检测矮秆基因(Rht1、Rht2、Rht8)、赤霉病抗性基因(Fhb1)、光周期基因(PpD-A1、PpD-D1)、多酚氧化酶基因(PPO18)、穗发芽抗性基因或QTL位点(Vp1B、Qphs.ccsu-3A.1),标记引物均由上海生工生物工程有限公司合成,详见表2。 PCR反应体系为15 μL,包括2×Mix混合液7.0 μL,上、下游引物各1.0 μL(2 μmol·L-1),ddH2O 4.0 μL ,DNA模板2.0 μL(100 ng· μL-1)。标记PFT-CAPS和His-InDel的扩增程序为:95 ℃预变性3 min;94 ℃变性30 s,65 ℃退火30 s,68 ℃延伸2 min 30 s ,33个循环;68 ℃终延伸7 min,4 ℃保存。其他标记的扩增程序为95 ℃预变性5 min;94 ℃变性1 min,58~65 ℃退火1 min,72 ℃延伸0.5~1.0 min,36个循环;72 ℃延伸10 min,4 ℃保存。标记WMS155、Xgwm261用6%的聚丙烯酰氨凝胶电泳检测,其他8个标记均用1.5%的琼脂糖凝胶电泳检测。 1.2.4 数据统计与分析 用Excle 2010进行数据统计;SPSS 20.0进行主成分分析;GRAPHPAD 7.00进行二维排序分析;R语言进行聚类分析。 2.1.1 主要农艺性状的主成分分析 对91份供试材料的12个重要农艺性状进行主成分分析,利用SPSS 20.0数据处理软件计算出各主成分的特征值和贡献率,提取到特征值大于1的主成分6个,累计贡献率为79.128%,详见表3。其中,第一主成分的特征值为1.946,贡献率为16.218%,载荷较高的农艺性状分别是千粒重、粒长,主要反映的是籽粒性状,因此命名为籽粒性状构成因子。向量间关系表明,籽粒越大,千粒重越高,但由于籽粒大小与穗粒数有一定的负相关关系,因此穗粒数会有所减少。第二主成份的特征值为1.901,贡献率为15.844%,载荷较高的农艺性状分别是顶芒芒长和侧芒芒长,主要反映芒长相关性状,因此命名为芒长构成因子。该主成分的向量间关系表明,芒长与籽粒性状、旗叶面积等均成负相关,而麦芒越长,与株高相关的性状越大,穗粒数越多。第三主成分的特征值为1.637,贡献率为13.642%,载荷较高的农艺性状分别是旗叶宽和旗叶长,主要反映旗叶相关性状,因此命名为旗叶构成因子。向量间关系表明,旗叶面积增大则株高和籽粒长度下降,而产量相关性状有所升高。第四主成分的特征值为1.536,贡献率为12.797%,载荷最高的性状是穗长,因此命名为穗长构成因子。向量间关系表明穗长增长则有效小穗数、千粒重等增加,但穗粒数和籽粒长度有一定下降。第五主成分的特征值是1.282,贡献率为10.686%,载荷最高的性状是穗颈长,该性状与株高相关,因此命名为株高构成因子。向量间关系表明,穗颈长增加则穗长、有效小穗数明显下降,千粒重和穗粒数略有升高但不明显。第六主成分的特征值是1.193,贡献率为9.94%,载荷最高的性状是穗粒数,因此命名为穗粒数构成因子。向量间关系表明穗粒数增多则有效小穗数增多,株高升高。 表2 引物和PCR检测结果Table 2 Primers and the fragment lengths of PCR detection 2.1.2 二维排序分析 以第五主成分值为横坐标,分别以第一、二、三、四、六主成分值为纵坐标利用GRAPHPAD 7.00绘图软件绘制二维排序图(图1、图2、图3、图4、图5)。第五主成分是株高控制因子,较小为好,第一主成分是籽粒性状构成因子,较大为好,因此第五主成分值较小且第一主成分值较大的品种主要分布在图1的左上角,这些品种整体表现为株高较矮且籽粒较大。第二主成分是芒长构成因子,由于芒长与千粒重、穗长及旗叶面积等性状成负相关关系,所以第二主成分适中较好,因此符合条件的品种主要分布在图2纵坐标轴的左边,围绕横坐标轴附近。第三主成分是旗叶性状构成因子,由于旗叶面积增大则株高和籽粒长度下降,且旗叶过大不利于植株通风、透光,容易引发病虫害,因此第三主成分适中较好。符合该特点的品种主要分布在图3纵坐标轴的左边,围绕横坐标轴附近。第四主成分是穗长构成因子,较大为好,因此符合该特点的品种主要分布在图4的左上角,这些品种表现为株高较矮、穗子较大。第六主成分是穗粒数构成因子,由于穗粒数与籽粒大小、千粒重在一定程度上呈负相关,因此穗粒数应适中偏大但不可过大,符合该特点的品种主要分布在纵坐标轴的左侧,围绕横坐标轴附近及横坐标轴上方,整体表现为穗粒数与籽粒大小及千粒重等性状结合较好。 表3 入选的6个主成分及其特征向量Table 3 Principal component analysis of agronomic traits 图1 第5、1主成分二维排序图 图2 第5、2主成分二维排序图 图3 第5、3主成分二维排序图 由以上分析可知,参试材料中综合性状较好的小麦品种编号为3、4、13、15、28、29、31、33、35、41、44、48、51、54、59、61、62、66、70、78、82、83、85、86、88、90。所对应的品种是郑麦1860、中农麦 4007 、秦禾麦2号、珍麦3号、圣麦 108、郑麦 151、豫农186、郑品麦22号、禾丰3号、泉麦31、安科1502、中育1220、丰德存麦16号、泰禾麦5号、淮麦4046、洛麦27、郑育麦21、怀川365、瑞华1426、郑麦119、西农979、西农585、西农B2、西农B3、西农B5、西农B7。 图4 第5、4主成分二维排序图 图5 第5、6主成分二维排序图 2.1.3 聚类分析 在主成分分析的基础上,以供试材料的主成分得分乘以对应主成分所占总贡献率的权重得到综合得分,利用综合得分进行聚类分析,在欧氏距离1.0处将供试材料聚为5类(图6),各类群的被测指标平均值具体见表4。第Ⅰ类包含2份材料,主要特点是株高最矮,穗长较长,旗叶面积较小,千粒重较轻,穗粒数最多,籽粒饱满度较差。第Ⅱ类包含34份材料,主要特点是株高较矮,穗长较短,千粒重中等,穗粒数较少。第Ⅲ类包含12份材料,主要特点是株高较矮,穗长最短,旗叶面积较小,千粒重最轻。第Ⅳ类包含1份材料,主要特点是株高、穗长、旗叶面积、穗粒数均最大,千粒重较高,籽粒饱满度稍差。第Ⅴ类包含42份材料,整体呈现为株高稍高,穗长较长,旗叶面积适中,千粒重最高,穗粒数较多。综上所述,第Ⅴ类材料虽株高较高但丰产潜力较好,第Ⅰ类、第Ⅱ类和第Ⅲ类材料株高虽然较矮但丰产性较差,第Ⅳ类材料虽然也具有较高的丰产潜力但其株高过高,农艺性状较差。因此,第Ⅴ类材料可以作为优质种质资源在生产上得以运用,其他材料则可以作为育种上的优良亲本,用于品种改良。 表4 各类群小麦农艺性状的平均值Table 4 The average value of agronomic traits in different clusters 图6 农艺性状系统聚类图 2.2.1 矮秆基因鉴定结果 矮杆基因检测结果表明,参试的91份材料中,有7份含有矮秆基因Rht1,占比为7.69%,代表品种有西农585、阜麦18、郑麦151、西农528等(图7);84份材料含有矮秆基因Rht2,占比为92.31%,代表品种有西农20、泉麦29、郑麦136、周麦18等(图8);79份材料含有Rht8,占比为86.81%,代表品种有西农979、西农585、泉麦29、中麦247等(图9)。在91份参试材料中,只含Rht1的材料仅有1份,为郑麦151,占比为1.10%;只含Rht2的材料有10份,占比为10.99%;只含Rht8的材料有2份,占比为2.20%;兼具Rht1和Rht8的材料有5份,占比为5.49%;兼具Rht2和Rht8的材料有73份,占比为80.22%;兼具Rht1和Rht2的材料仅有2份,占比为2.20%;同时含有Rht1、Rht2和Rht8的材料仅有1份,为安科1403,占比为1.10%。 2.2.2 多酚氧化酶基因鉴定结果 在PPO-A1位点,91份参试材料中有27份为PPO活性较低的PPO-A1b基因型,频率为29.67%;有64份为PPO活性较高的PPO-A1a基因型(图10),频率为70.33%。 M:DL2000; 1~7:部分供试材料; a:Rht-B1a基因的扩增条带; b:Rht-B1b基因的扩增条带。 M:DL2000; 1-7:Part materials tested;a:Band ofRht-B1a;b:Band ofRht-B1b. 图7矮秆基因Rht1在部分参试材料中的检测结果 Fig.7The detection results of dwarfing geneRht1in some tested materials M:DL2000; 1~7:部分供试材料; a:Rht-B1a基因的扩增条带; b:Rht-B1b基因的扩增条带。 M:DL2000; 1-7:Part materials tested;a:Band ofRht-B1a;b:Band ofRht-B1b. 图8矮秆基因Rht2在部分参试材料中的检测结果 Fig.8The detection results of dwarfing geneRht2in some tested materials M:DL500; 1~12:部分供试材料。 M:DL500; 1-12:Part materials tested. 图9矮秆基因Rht8在部分参试材料中的检测结果 Fig.9The detection results of dwarfing geneRht8in some tested materials 2.2.3 光周期基因鉴定结果 PpD-A1(图11)和PpD-D1(图12)两个光周期基因均具有a、b两种等位变异,PpD-A1a基因型(580 bp)和PpD-D1a基因型(186 bp)对光周期反应不敏感,PpD-A1b基因型(274 bp)和PpD-D1b(172 bp &186 bp)基因型对光周期反应敏感,经检测,参试材料均为PpD-A1a+PpD-D1a基因型,因此91份材料均为光周期非敏感型材料。 M:DL1000; 1~12:部分供试材料。 M:DL1000; 1-12:Part materials tested. 图10多酚氧化酶基因PPO-A1在部分参试材料中的检测结果 Fig.10The detection results of polyphenol oxidase genePPO-A1in some tested materials M:DL1000; 1~12:部分供试材料。 M:DL1000; 1-12:Part materials tested. 图11光周期基因PpD-A1在部分参试材料中的检测结果 Fig.11The detection results of photoperiod genePpD-A1in some tested materials M:DL1000; 1~12:部分供试材料。 M:DL1000; 1-12:Part materials tested. 图12光周期基因PpD-D1在部分参试材料中的检测结果 Fig.12The detection results of photoperiod genePpD-D1in some tested materials 2.2.4 赤霉病基因鉴定结果 91份供试材料中,经标记PFT-CAPS扩增后共检测到17份材料含有2 077 bp/2 076 bp片段(图13),进一步经DraⅠ内切酶酶切后发现,有5份材料为PFT-Ⅰ型(2 077 bp),分布频率为5.49%;有12份材料为PFT-Ⅱ型(1 567 bp& 709 bp),分布频率为13.19%。在His位点,91份供试材料均检测到2 061 bp片段(图14),说明这些材料均非His-Ⅰ型(1 309 bp)。由此可知,供试的91份材料均不含Fhb1基因。 M1:DL5000; M2:DL2000; 1~10:部分供试材料。 M1:DL5000; M2:DL2000; 1-10:Part materials tested. 图13PFT-CAPS标记在部分参试材料中的检测结果 Fig.13Detection results ofPFT-CAPSmarker in some tested materials M2:DL2000; 1~10:部分供试材料。 M2:DL2000; 1-10:Part materials tested. 图14His-InDel标记在部分参试材料中的检测结果 Fig.14Detection results ofHis-InDelmarker in some tested materials 2.2.5 穗发芽抗性基因鉴定结果 91份供试材料中,有37份材料含抗穗发芽基因Vp1Bc(569 bp),分布频率为40.66%,平均发芽率为45.5%;有54份含感穗发芽基因Vp1Ba(652 bp),分布频率为59.34%,平均发芽率为57.6%;未出现Vp1Bb(845 bp)基因型材料(图15)。对不同基因型进行方差分析发现,Vp1Ba基因型的发芽率明显低于Vp1Bc基因型,呈显著差异(F=16.97,P<0.01)。 用Xgwm155标记在91份供试材料中共扩增出A、B、C、D四种片段类型,其中扩增出A片段类型的仅有1个,占供试材料的1.10%,整穗发芽率为75%;扩增出B片段类型的有16个,占供试材料的17.58%,整穗发芽率均值为52.9%;扩增出C片段类型的有19个,占供试材料的20.88%,整穗发芽率均值为44.7%;扩增出D片段类型的有55个(图16),占供试材料的60.44%,整穗发芽率均值为55.0%。方差分析表明,扩增出B型片段的品种与扩增出C型和D型片段的品种发芽率没有显著差异,而扩增出C型和D型片段的两类品种的发芽率具有显著性差异(F=7.318,P<0.05)。由于含有A带型的材料太少,所以含有该带型材料的发芽率还有待验证。 M:DL1000; 1~12:部分供试材料。 M:DL1000; 1-12:Part materials tested. 图15Vp1B3标记在部分参试材料中的检测结果 Fig.15Detection results ofVp1B3marker in some tested materials 1~12:部分供试材料。 1-12:Part materials tested. 图16WMS155标记在部分参试材料中的检测结果 Fig.16Detection results ofWMS155markerin some tested materials 结合整穗发芽实验,从参试的91份材料中筛选出21份SGR≤40%的抗穗发芽品种,占总数的23.08%。标记基因型方差分析表明,Vp1B3和Xgwm155均与穗发芽抗性有关。21份抗穗发芽品种中,15份为Vp1Bc型,6份为Vp1Ba型,占比分别为71.43%、28.57%;用标记WMS155扩增出A、B、C、D带型的品种个数分别为0、3、8、10,占比分别为0、14.29%、38.10%、47.62%。用两个标记同时可检测到抗性条带的有7份,占参试材料的7.69%,分别为西农20、徽研66、郑麦151、淮麦4046、安科1405、西农501、西农585,穗发芽率均值为35.3%,这些材料可作为穗发芽抗性改良的重要抗源用于后续育种。 3.1.1 主成分分析 主成分分析采用降维的思想,以较少的几个不相关的变量来反映多变量信息,将其用于农艺性状的评价分析时,可实现用几个综合指标来把握若干性状的整体信息,为数据分析提供便利[19]。本研究通过对91份参试材料进行主成分分析,将12个复杂农艺性状浓缩成6个互为独立的综合因子,可以解释79.128%的表型变异。就本研究的6个主成分因子而言,对小麦产量的贡献率依次是:籽粒性状构成因子>芒长构成因子>旗叶构成因子>穗长构成因子>株高构成因子>穗粒数构成因子。千粒重是小麦的产量构成三要素之一,本研究中第一主成分载荷最高的是千粒重因子(0.836),表明了千粒重对小麦的产量起着极其重要的作用,这与张中州等[20]的研究结果相符。根据高产育种的标准及各性状间的相互关系,选择品种应满足的条件是:第一主成分较大、第二主成分适中、第三主成分适中、第四主成分较大、第五主成分较小,第六主成分适中偏大。由此可知郑麦151,西农585等26个小麦品种均为满足以上标准的优良品种。 3.1.2 聚类分析 聚类分析是一种依据遗传距离远近对材料进行分类的研究方法,实践证明,聚类分析在研究品种资源的差异和分类方面可行[14-15]。基于主成分分析进行聚类分析既可以保证生物信息的完整性又可以简化数据处理程序,得到更为可靠的结果,为以后杂交育种的亲本选配提供参考,是较为科学的数据分析方法[2,21]。本研究基于主成分分析的结果对91份供试材料进行聚类分析,采用Q型聚类,在遗传距离1.0处将供试材料聚为5类,可以较好的揭示出各类群间及类群内品种的差异性,同时也能反映出黄淮麦区新育成小麦品种(系)的遗传距离较近,遗传基础较为狭窄的现状。马艳明等[22]利用11对ISSR引物分析了我国黄淮麦区96个小麦推广品种(系)的遗传多样性,分析结果表明,参试品种间具有较高的遗传相似度,其中64.6%的品种被聚在同一类,这与本研究结果一致。推测出现这种现象的原因可能是育种工作中所用骨干亲本较为有限。因此,在后续的育种中应注重扩宽亲本选择范围,尽量选择遗传背景差异较大的合适亲本配制杂交组合。 3.1.3 二维排序分析 二维排序分析法能够简洁、直观的方式反映参试材料的分布特点及品种间的性状差异。陶爱芬等[23]基于主成分分析对红麻的产量性状和品质性状进行了二维排序分析,筛选出了在品质或产量上表现突出和兼具高产与优质的品种,明确了各参试材料的优点与不足,为后续的生产利用提供了有力参考。本研究通过二维排序分析对91份材料的12个农艺性状进行综合分析,共筛选出26份矮杆、高产且综合性状优良的品种。其中已经通过国家区试的优异小麦品种西农979和西农585同时出现在聚类分析及二维排序分析的结果中,且入选的其他品种在两种分析方法中的吻合度也很高,由此可见,基于主成分分析的二维排序分析及聚类分析在分析结果上具有很高的一致性,前者能够客观地反映各品种在农艺性状上的差异,而后者则可以反映各品种的遗传距离及各类群品种之间的差异,两种方法各有所长,但都能较好的对种质资源的综合农艺性状进行评价,这与前人的研究结果基本一致。在数据分析中两种分析方法结合起来,能够从多个角度对种质资源的农艺性状或者品质性状进行更为全面的评价,为小麦的生产运用及后续育种工作提供科学依据。 在育种实践中,将分子标记辅助选择与表型性状的选择相结合,能够在很大程度上提高育种效率。本研究共利用10个分子标记对参试的91份材料的不同性状进行鉴定。结果发现,在矮杆性状上,黄淮麦区小麦品种中Rht2和Rht8存在的比例较高,这与张德强等[18]的研究结果一致,推测可能与该地区的气候条件及近年来所育成品种的遗传基础较为狭窄有关。对多酚氧化酶活性相关基因,陈 泠等[24]的研究表明,陕西和河南的小麦品种中含PPO-A1b的比例较低。本研究中,有69%的参试材料来自河南和陕西,有29.67%的参试品种具有低活性多酚氧化酶基因,这与前人的报道是吻合的。在光周期基因上,参试品种经检测均属于光周期不敏感型,这与邓跟望等[25]的研究结果相一致,但孙道杰等[26]的研究表明,一定的光周期敏感性会增强品种的生殖发育稳定性(抽穗期稳定性),因此小麦品种应具有一定的光周期敏感性。在赤霉病抗性上,91份参试材料均不含对赤霉病抗性最强的Fhb1[27]基因,但有5个材料含有PFT-Ⅰ,后续工作中可尝试利用含有His-Ⅰ基因型的品种为亲本,对黄淮麦区小麦品种的赤霉病抗性加以改良。在穗发芽抗性上,参试材料整体表现为穗发芽抗性较差,抗性材料所占比例为23.08%,这与马文洁等[28]的研究结果一致。由此可知小麦的穗发芽抗性也是黄淮麦区品种亟待改良的性状。 综上可知,黄淮麦区近年来育成的品种(系)在产量、品质和抗性方面有一定的提升,但仍有很大的发展空间。新育成的品种中郑麦151、西农585等综合性状优良,可用于后续推广。鉴于黄淮麦区小麦品种的遗传基础较为狭窄,后续育种中应选择遗传背景较为丰富的材料作为杂交亲本,重点改良品种的穗发芽抗性、赤霉病抗性,其次也要提高对低多酚氧化酶活性材料的选育,以改良面粉(团)的色泽,满足现代人的消费需求。再者,近年来全球极端气候增多,气候不稳定性增强,为了使小麦具有稳定的抽穗期,保证粮食产量,在后续的育种中要注重培育具有适度光周期敏感性的品种。
2 结果与分析
2.1 农艺性状鉴定结果


2.2 供试材料分子标记鉴定结果

3 讨 论
3.1 小麦主要农艺性状的分析方法
3.2 小麦分子标记鉴定
猜你喜欢
当代水产(2022年7期)2022-09-20
河北科技师范学院学报(2022年2期)2022-08-26
河南农业·综合版(2022年2期)2022-03-18
中国饲料(2021年17期)2021-11-02
空间科学学报(2021年1期)2021-05-22
落叶果树(2021年6期)2021-02-12
甘肃教育(2020年18期)2020-10-28
天津农林科技(2020年3期)2020-08-13
四川蚕业(2020年2期)2020-07-10
山西中医药大学学报(2020年2期)2020-06-06
