
2,4-表油菜素内酯缓解水稻花期高温胁迫的生理机制
2019-06-13 03:02高健王亚梁孙磊赵渊刘连盟侯雨萱王玲黄世文
中国稻米 2019年3期
高健 王亚梁 孙磊 赵渊 刘连盟 侯雨萱 王玲黄世文
(中国水稻研究所/水稻生物学国家重点实验室,杭州310006;*通讯作者:wangling03@caas.cn;huangshiwen@caas.cn)
自从工业革命以后,CO2排放量增加,地表温度不断上升。IPPC(2013)报告指出,在100多年来,全球地表平均温度升高了0.85℃,预计未来100年左右,温度大约会再提高0.3℃~4.8℃[1]。据估计到21世纪末,温度上升可能使我国水稻的产量降低41%[2]。长江中下游地区是我国主要的水稻种植区域,该地区水稻受到高温的影响非常明显。有研究表明,2017年合肥市水稻开花期在7月22日至28日期间遇到38℃以上的高温,导致部分水稻品种的结实率下降15%~25%[3]。在水稻的生育期中,花粉母细胞减数分裂期和开花期对高温胁迫最敏感,在这两个时期内遭遇高温,会导致水稻产量显著下降[4-5]。
目前,培育耐热型水稻品种是抵御高温最重要的方法,耐热品种受高温胁迫伤害较小,易形成大穗,结实率高[6]。同时,调整栽培措施也是一种重要的农艺措施。比如,分期播种错开水稻花期高温,或者高温逆境时进行深水灌溉[6-7]。但是耐热型水稻品种少,培育耐热型水稻品种时间缓慢,而调整栽培措施也会产生一些问题,比如,调整播期能够避开高温,但避开高温的同时可能增大病虫害发生的可能,因此,喷施化学调控剂不失为一种有效的方法[8-9]。已报告能减轻植物高温伤害的化学调节剂有激素类、渗透压调节剂、抗氧化剂、信号分子、多胺和微量元素等[10]。与其他类型的化学调节剂相比,激素类在植物抗逆方面的作用更为明显。已有研究表明,喷施水杨酸、茉莉酸甲酯等植物激素可通过提高植物的抗氧化酶活性、光合作用以及可溶性糖含量等来避免或减轻作物受到高温伤害[6,11]。
油菜素内酯(Brassinosteroids,BRs)是多羟基化类固醇类植物激素,能影响植物对外界信号的反应,在植物生长发育方面有着重要作用。BRs不仅能影响植株大小、雄性生育力、开花和光合作用等[12-13],而且可能会增加各种生物和非生物逆境的适应性[12,14],被认为是“辅助植物修复”的潜在激素。2,4-表油菜素内酯(EBR)是BRs的一种,在减轻植物高温胁迫方面有着优良的效果。谢云灿等[13]研究表明,喷施EBR不仅能改善大豆品质,还能减轻大豆高温胁迫下的伤害,提高产量。张永平等[15]研究表明,EBR有利于高温胁迫下甜瓜幼苗抗氧化酶活性的提高和对光能的捕获与转换,对甜瓜幼苗的生长有利。BRs在其他作物耐热性方面的研究已有一定的报道,但是在缓解水稻高温逆境方面的研究甚少。本研究以耐热型水稻品种黄华占为试验材料,探究2,4-表油菜素内酯缓解水稻高温热害的作用机制,为化学调控减轻水稻高温伤害提供理论依据。
1 材料与方法
1.1 参试材料
供试水稻品种为耐热型品种黄华占[16]。
1.2 试验设计
在水稻破口期,将盘栽苗移入人工气候箱进行高温胁迫处理。高温处理为:0∶00-7∶00,31℃;7∶00-9∶00,32.5℃;9∶00-14∶00,38℃;14∶00-16∶00,36.5℃;16∶00-24∶00,31℃。常温对照处理为:0∶00-7∶00,25℃;7∶00-9∶00,26.5℃;9∶00-14∶00,32℃;14∶00-16∶00,30.5℃;16∶00-24∶00,25℃。人工气候箱中的相对湿度控制在75%左右。高温处理5 d后,将水稻植株移置自然环境下。
本试验共设4个处理:NT-CK,常温喷施清水;NT-EBR,常温喷施 0.55 mg/mL 的 EBR;HT-CK,高温胁迫下喷施清水;HT-EBR,高温胁迫下喷施0.55 mg/mL的EBR。在水稻花粉母细胞减数分裂中后期进行药剂喷施处理,每隔3 d喷1次,共喷施3次,每次喷到药液在叶片自然滑落为止。每个处理3次重复。
1.3 测定项目及方法
1.3.1 花粉表型观察和产量因素
于高温处理后第3 d,从每穗上、中、下部位各取3个颖花,用1%I2-KI溶液染色在荧光显微镜下(OLYMPUS,BX61)观察花粉活力,取所有当天开花的颖花在体式显微镜下(OLYMPUS,SZX9)观察花药破裂率,计算花粉育性和花药破裂率[花粉活力(%)=染色花粉粒数/花粉粒总数×100;花药破裂率(%)=(待测花药数-未破裂花药数)/待测花药数×100]。每个处理3次重复。于水稻成熟后,收取20个穗子,考查千粒重、每穗总粒数和实粒数。每个处理3次重复。
1.3.2 相关生理指标
高温处理第3 d,取新鲜颖花用于生理指标的测定。超氧化物歧化酶(SOD)活性测定采用Giannopolitis和Ries的方法[17];POD活性测定采用Maehly和Chance的方法[18];CAT活性测定采用李仕飞等的方法[19];丙二醛含量的测定采用Dhindsa等方法[20];脯氨酸含量的测定采用张殿忠的方法[21];可溶性糖含量的测定用蒽酮比色法[22]。
1.3.3 内源激素含量
高温处理3 d后,采用符冠富等的方法[7]测定内源激素含量。称取0.1 g颖花鲜样,加4℃放置的1 mL提取液(80%甲醇中含有1 mmol/L二叔丁基甲苯酚),冰上匀浆后,4℃提取4 h,4℃ 10 000 r/min离心10 min,定容5 mL,安捷伦C18SPE处理后氮吹,用样品稀释液溶解,采用酶联免疫吸附法(Enzyme Linked Immuno Sorbent Assay,ELISA)测定ABA和GA含量。
1.3.4 ABA合成相关基因OsNECD2的表达分析
高温处理3 d后,取颖花鲜样,用常规TRIGOL法提取RNA。cDNA的合成用TaKaRa的实时定量反转录试剂盒(PrimeScriptTM RT Master Mix),使用Applied Biosystems公司的ABI7900进行相关基因表达量的测定。PT-PCR运行程序:预变性95℃ 1.5 min;95℃ 10s,60℃ 30 s,72℃ 20 s,40 个循环。采用 2-△△CT进行相对表达量的计算。OsNECD2的上游引物序列5’-AGCACAGAGGATTGCACCATT-3’,下游引物序列 5’-TGTTGTCTCCAAGCCTTGATTT-3’;内参基因ubi的上游引物序列 5’-GCTCCGTGGCGGTATCAT-3’,下游引物序列 5’-CGGCAGTTGACAGCCCTAG-3’。
1.4 统计分析
采用 Microsoft Excel 2010和SAS 9.2进行数据的统计和分析,用Duncan’s法检验处理间差异。
2 结果与分析
2.1 喷施EBR对产量构成因素的影响
从表1可见,与常温对照相比,高温胁迫后水稻结实率降低42.79%,而对每穗总粒数、千粒重没有显著影响;在常温下,喷施EBR对结实率、穗总粒数以及千粒重均无显著影响。高温胁迫下,喷施EBR相对喷施清水使水稻结实率提高了24.09%,而喷施EBR对穗总粒数和千粒重无显著影响。
2.2 喷施EBR对花粉活力以及花药开裂的影响
花粉活力和花药开裂率是影响水稻结实的重要因素。各个处理下的花粉活力以及花药开裂率的调查表明,与常温对照相比,高温胁迫后水稻花药开裂率降低36.87%,穗子中、下部位的花粉活力分别降低8.38%和21.1%;在常温下,喷施EBR对花粉活力和花药开裂并无显著影响。在高温胁迫下,喷施EBR相对喷施清水,花药开裂率提高55.4%,穗子中部和下部的花粉活力分别提高了8.64%和24.2%(表1)。
2.3 对抗氧化酶的影响
SOD、POD、CAT能清除植物体内产生的活性氧(ROS),防止膜脂质过氧化。测定结果表明,与常温对照相比,在高温胁迫下SOD、POD、CAT的酶活性分别上升了29.83%、101.37%和91.73%,常温下喷施EBR后SOD的酶活性提高了51.76%,对POD和CAT无显著影响。高温胁迫下,喷施EBR相对喷施清水SOD、POD酶活性分别上升了20.4%和30.9%,对CAT无显著影响(表2)。
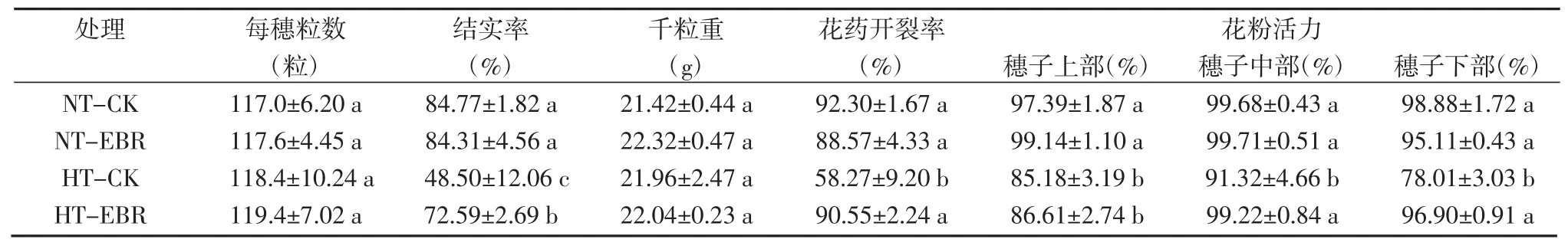
表1高温胁迫下EBR对水稻产量及花粉育性的影响
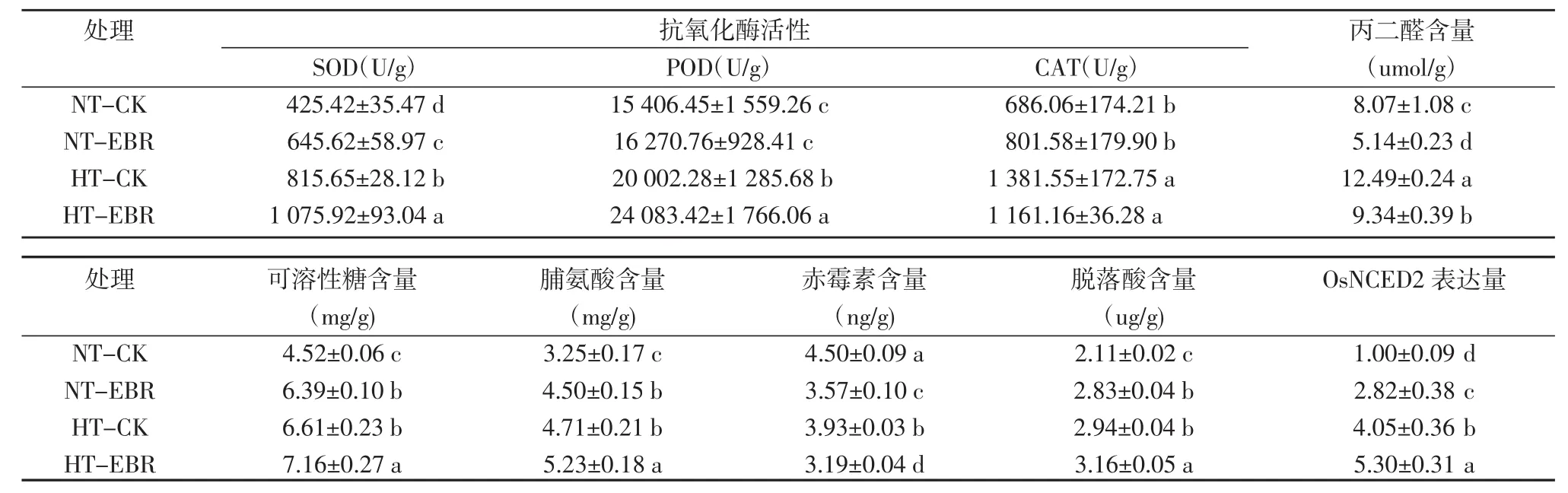
表2高温胁迫下EBR对水稻生理指标的影响
2.4 对丙二醛含量的影响
MDA是评价脂膜受损程度的指标之一。测定结果表明,与常温对照相比,高温胁迫下MDA含量显著上升54.77%,常温下喷施EBR MDA降低了36.23%;高温胁迫下喷施EBR相对喷施清水MDA下降了25.14%(表 2)。
2.5 对可溶性糖和脯氨酸含量的影响
可溶性糖和脯氨酸的积累有助于保持细胞渗透平衡。测定结果表明,与常温对照相比,高温胁迫下可溶性糖和脯氨酸的含量显著上升了46.23%和45.13%,常温下喷施EBR后可溶性糖和脯氨酸含量升高了41.34%和38.57%;而高温胁迫下喷施EBR后可溶性糖和脯氨酸含量分别提高8.23%和11.02%(表2)。
2.6 对GA和ABA含量的影响
植物激素间的相互作用可以调控植物的生长发育。测定结果表明,与常温对照相比,高温胁迫下GA含量下降12.54%,ABA含量上升39.25%,常温下喷施EBR后GA含量下降20.69%,ABA含量上升了32.27%;高温胁迫下,喷施EBR后GA含量下降18.81%,ABA含量上升7.48%(表2)。
2.7 对ABA合成基因OsNCED2表达量的影响
OsNCED2是控制ABA的合成代谢途径的关键基因。基因表达量分析表明,与常温对照相比,高温胁迫下,OsNCED2表达量上升305.16%,常温下喷施EBR后OsNCED2表达量上升了181.58%;高温胁迫下,喷施EBR后OsNCED2表达量上升30.67%(表2)。
3 讨论
花粉的萌发、授粉、花药开裂等生理活动易受到高温影响,从而导致水稻的败育,结实率下降,产量降低。高温胁迫下,颖花观察是高温指标中的重要因素。张桂莲等[23]的研究发现,在高温胁迫下,水稻花药开裂受到抑制,花粉粒数减少,花粉活性降低,导致小穗育性降低,结实率下降,这与本研究的结果一致。在本研究中,常温下喷施EBR对花粉活力、花药开裂无显著影响,而高温胁迫下喷施EBR能使水稻花药和穗子中、下部位的花粉的活性提高,有利于水稻结实,这说明高温胁迫下花粉育性的提高是由于喷施了EBR的缘故,EBR有助于维持水稻本身的抗逆性。
高温胁迫会导致植物体内的活性氧(ROS)的过量产生[24],如超氧自由基(O2-)、羟基自由基(OH)和过氧化氢(H2O2)等,导致膜脂质过氧化,蛋白质变性和核酸损伤[25]。为了阻止高温胁迫下ROS对细胞的损害,植物中的一系列的抗氧化活性酶,包括SOD、、POD、CAT、APX,以及AsA和GSH等非酶系统的活性上升,可能有效的清除ROS,防止膜脂质过氧化,减轻高温伤害[26-27]。在本研究中,喷施EBR诱导了水稻SOD和POD的酶活性增加,有助减轻高温下ROS积累带来的损伤。然而,尚不清楚EBR是否直接或间接调节植物对氧化应激的反应[28]。
在胁迫条件下的植物已经进化出在胞质溶胶中积累相容的渗透物用于细胞内维持渗透稳态,这些渗透物包括脯氨酸、可溶性糖以及可溶性蛋白[25]。这些渗透物不仅在渗透调节中起作用,其中脯氨酸还能参与氮代谢、能量代谢以及保护生物大分子,可溶性糖能保持细胞膜和原生质胶体的稳定,并为有机物的合成提供碳架[29]。高温逆境提高了脯氨酸和可溶性糖含量,EBR处理促进了这两种渗透保护剂的积累,表明EBR可能会促进渗透物的积累,这可能与这些植物的耐热性提高密切相关。
植物激素间的相互作用可以调控植物的生长发育,已有众多研究揭示了BR和其它激素的相互作用[30-31]。GA和BR都是促进生长的激素,但是它们之间存在拮抗关系[32]。有研究表明,在水稻根部,当施加过量活性BR时,该激素主要通过GA失活基因GA2ox-3的上调诱导GA失活,导致激素水平降低和生长抑制[33]。DELLA家族蛋白,在GA信号分子中在N端具有DELLA结构域的蛋白。有研究表明,GA含量的降低有利于DELLA的积累,并且DELLA家族蛋白在环境压力下能参与植物适应性生长[34]。在本研究中,外源EBR的处理使得GA含量降低,这可能是外源EBR使水稻能维持较高抗逆性的原因。ABA可以增强抗氧化酶活性、调节光合作用、促进细胞吸水、调节气孔、缓解逆境伤害[35-37]。而高温处理可以使ABA含量增加[38],从而调整植物适应高温胁迫。OSRK1/SAPK6和SAPK4参与ABA信号传导,而喷施EBR可能通过影响这些蛋白磷酸化水平来影响ABA信号传导[39]。在本研究发现,喷施EBR GA含量下降,ABA含量上升,这说明EBR能通过内源激素的变化来达到减轻高温对水稻的伤害的目的。
9-顺-环氧类胡萝卜素双加氧酶(NCED)是水稻和拟南芥ABA生物合成的限速酶,在水稻中存在5个NCED基因,包括OsNCED1、OsNCED2、OsNCED3、Os-NCED4和OsNCED5,其中仅有OsNCED3基因表达受干旱胁迫诱导[40],但是在我们的研究中,高温可以诱导OsNCED2基因的表达,喷施EBR进一步诱导Os-NCED2的表达,OsNCED2是否是高温下ABA的生物合成关键基因以及喷施EBR是否是影响OsNCED2的表达进而提高ABA的含量还需进一步研究。
4 结论
本研究表明,在高温胁迫下,喷施EBR能通过提高抗氧化酶的活性、渗透保护剂的积累,以及内源激素的调节,来维持花粉育性和花药的正常,从而减轻高温对水稻结实的伤害。本研究提供了EBR缓解水稻高温伤害的技术支持,对高温逆境下水稻的保产措施有着参考作用。
猜你喜欢
中老年保健(2022年3期)2022-11-21
麦类作物学报(2022年8期)2022-08-08
哈尔滨轴承(2021年1期)2021-07-21
安徽化工(2021年3期)2021-05-29
保鲜与加工(2021年1期)2021-02-06
棉花学报(2020年3期)2020-08-08
科学与财富(2019年15期)2019-10-21
蔬菜(2019年5期)2019-05-21
江苏农业科学(2016年8期)2017-02-15
天津农业科学(2015年9期)2015-09-02
