
福建省露地及设施蔬菜烟粉虱越冬种群密度及耐寒性研究*
2019-06-12 07:46姚凤銮丁雪玲卢学松何玉仙翁启勇
中国生态农业学报(中英文) 2019年6期
姚凤銮,郑 宇,丁雪玲,卢学松,何玉仙**,翁启勇**
(1.农业部福州作物有害生物科学观测实验站/福建省作物有害生物监测与治理重点实验室/福建省农业科学院植物保护研究所福州 350013;2.闽台作物有害生物生态防控国家重点实验室/福建农林大学 福州 350002)
烟粉虱[Bemisia tabaci(Gennadius)]广泛分布于除南极洲以外的各大洲,是热带、亚热带及相邻温带地区的一种重要害虫[1]。20世纪90年代,MEAM1隐种(原称为B生物型)入侵我国并迅速扩散,在广东、北京、河北、山东等地相继暴发,造成了严重的经济损失,成为我国蔬菜等作物上的重要害虫[2-3]。MED隐种(原称为Q生物型)是近年来引起人们高度重视的一种新的隐种,我国于2003年首次在云南发现[4],目前已广泛入侵我国大部分省市[5-7]。
温度是影响烟粉虱种群发展的重要因素之一[8-9],低温胁迫直接影响烟粉虱的越冬[10-11]。王勇等[10]研究发现,烟粉虱不耐低温,不能在江苏地区(30°45′0″~35°19′8″N)的露地越冬。董国等[12]认为烟粉虱在我国浙江台州(28°40′48″N)以北地区不能在室外露地越冬。崔洪莹等[11]依据等温线绘制烟粉虱越冬北界模型图,推测烟粉虱在我国的自然越冬北界分布线为浙江省金华市(29°4'48″N)、江西省南昌市(28°40′48″N)、湖北省仙桃市(30°22′12″N)以及四川 省(26°3′36″~34°19′12″N)和云 南 省(21°7′48″~29°15′0″N)的大约北纬 28°的地区。但是,周国珍等[13]的研究表明烟粉虱在湖北省(29°5′24″~33°19′48″N)露地能安全越冬。尽管如此,这些研究主要关注烟粉虱的越冬北界[10-13],而在远离越冬北界、气候较温暖且适宜露地烟粉虱越冬的地区,越冬烟粉虱的虫态、种群数量特征还鲜见报道。
福建省的纬度介于 23°30′36″~28°22′12″N,根据上述研究结果[10-13]推测,烟粉虱可以在福建地区露地越冬。然而,由于福建省横跨中亚热带和南亚热带,南北气候差异大,地形特征以山地丘陵为主,形成了多种多样的地方性气候以及与地理环境要素密切相关的区域农业生态系统,加上福建省近年来大力发展设施农业等因素,因此,除了与纬度、海拔直接相关的温度条件外,不同地区之间的作物品种布局和栽培方式的差异亦可能影响冬季烟粉虱的发生数量。另外,尽管众多研究认为杂草是烟粉虱的越冬寄主,但烟粉虱在杂草上的生存适宜性鲜见报道[14]。
目前,有关福建省烟粉虱隐种(生物型)的发生和分布、抗药性发展动态已有研究报道[7,15-16],但关于福建省烟粉虱田间种群消长动态及其冬季发生情况目前尚少见报道。越冬虫量通常与翌年害虫的发生数量关系密切[17],因此,越冬烟粉虱的虫量调查可以为该虫的科学防治提供理论依据。鉴于此,本文通过田间调查福建地区露地和温室大棚冬季烟粉虱发生情况,室内测定烟粉虱的耐寒性,以及杂草作为烟粉虱寄主的适宜性,以期明确福建地区烟粉虱的越冬规律,阐明不同地区冬季烟粉虱发生数量差异的生态学机制,增进对烟粉虱在我国南方较温暖地区越冬规律的认识,为科学制订烟粉虱综合防治技术方案提供理论依据。
1 材料与方法
1.1 调查时间和地点
福建省漳州市和南平建瓯市是福建省两个重要蔬菜生产地,因此,选择漳州市和南平建瓯市分别作为此次烟粉虱越冬调查的南北界。2017年1月9—12日和2月21—24日,在福建省南平市建瓯市小松镇(27°6′36″N,118°21′36″E,海拔 134.6 m)、三明市将乐县光明乡(26°46′48″N,117°24′0″E,海拔 210.95 m)、三明市三元区中村乡(26°9′36″N,117°41′24″E,海拔439.8 m)、福 州 市 闽 侯 县 南 通 镇(25°55′12″N,119°13′48″E,海拔 32.14 m)、龙岩市新罗区东肖镇(25°0′0″N,117°0′0″E,海拔 427.9 m)和漳州市龙海市东园镇(24°22′12″N,117°51′36″E,海拔 22.64 m)等地开展调查。其中,前三者调查地点位于中亚热带,福州和龙岩的调查地点位于南亚热带北界附近,而漳州的调查地点位于南亚热带。
1.2 越冬寄主调查方法
在6个调查地区,选择烟粉虱常年发生较为严重的露地和温室大棚蔬菜地,广泛采集蔬菜作物和杂草,每种植物采集10~15株,带回室内镜检确定烟粉虱越冬寄主。植物鉴定参照《中国农田杂草图册》[18]和《农田杂草识别及防除——幼苗和成株简明图鉴》[19]。
1.3 越冬种群密度调查方法
选取面积不少于667 m2的蔬菜温室大棚和露地,采用五点取样法调查处于生长中后期蔬菜上的烟粉虱虫量。花椰菜(Brassica oleraceaL.var.botrytisL.)的生育期为莲座期、花球形成期和采后残株期;茄子(Solanum melongenaL.)的生育期为座果期;辣椒(Capsicum annuumL.)、黄瓜(Cucumis sativusL.)和豇豆[Vigna unguiculata(Linn.)Walp]的生育期为采后残株期。每个地区调查 3~7块露地,在南平建瓯、三明将乐和龙岩新罗各调查 5~7个温室大棚。调查成虫时,每点调查 4株,每株仅调查顶部展开的 5片真叶,共调查 100片叶;调查卵和若虫时,每点调查 2株,每株寄主采集上、中、下 3片叶,共查30片叶。对于辣椒等叶片较小的作物,以1根枝条代表1片叶。调查时记录采集或观测的样地数量和生育期。将寄主叶片带回室内,在解剖镜下观察记录烟粉虱卵和若虫数量。以头·叶-1为单位表示每个样地成、若虫密度,以粒·叶-1为单位表示每个样地卵密度。
选取面积不少于667 m2的处于休耕期的蔬菜温室大棚和露地,随机调查处于生长期杂草上的烟粉虱虫量。在南平建瓯、三明将乐和龙岩新罗各调查3~6块露地,3个温室大棚。每块样地至少调查10株。调查成虫时,每株调查寄主顶部展开的 5片真叶,共调查至少50片叶;调查卵和若虫时,每株寄主采集上、中、下3片叶,共调查至少30片叶,在解剖镜下观察并分别记录卵和若虫的数量。调查时记录采集或观测的叶片数量和样地数量。以头·叶-1为单位表示每个样地成、若虫密度,以粒·叶-1为单位表示每个样地卵密度。
1.4 低温对烟粉虱各虫态存活的影响
福建省各地区2017年1—2月份日最高温和最低温情况见图1。其中,南平建瓯极端低温 0~4℃,持续时间 4~6 d;三明极端低温 3~5℃,持续时间2~5 d;三明将乐极端低温 3~5℃,持续时间 3~6 d;福州闽侯极端低温3~5℃,持续时间4~6 d;龙岩极端低温3~5℃,持续时间3~4 d;漳州龙海极端低温6~8℃,持续时间 3~5 d。鉴此,本研究中低温分别设置为0℃、4℃、8℃,处理时间分别设置为6 h、24 h、48 h、96 h和144 h。低温胁迫试验在±1℃、相对湿度(65±5)%、光照14 h/10 h(L/D)的人工气候箱(三洋MIR-154)中进行。
在25℃室温条件下,取 20~30头混合日龄MEAM1隐种烟粉虱雌成虫,用微虫笼固定于无虫花椰菜叶片背面,产卵8 h后去除成虫,饲养待用。之后,将带有1日龄卵、低龄若虫(产卵后8 d,包括1~2龄若虫)、高龄若虫(产卵后16 d,包括4龄若虫和伪蛹)的花椰菜叶碟(直径 35 mm),利用培养皿琼脂保湿法进行培养,即将叶碟正面朝下置于具透气网培养皿(直径 35 mm)的琼脂上。另选取 4~7日龄成虫(雌雄数量比为1∶1)置于上述利用培养皿琼脂保湿法培养的无虫花椰菜叶碟上。在解剖镜下观察记录初始卵、若虫和成虫数量后,在0℃、4℃、8℃条件下处理6 h、24 h、48 h、96 h和144 h,然后移到(25±1)℃、相对湿度(65±5)%、光照 14 h/10 h(L/D)的人工气候箱中(一恒 MGC-350HP)继续饲养,逐日观察存活率。卵和低龄若虫发育至下一虫龄,高龄若虫发育至成虫视为存活,否则视为死亡。每处理重复3次。

图1 各调查点2017年1—2月每日最高温和最低温Fig.1 Daily maximum and minimum temperatures of each sampling site in January and February of 2017
1.5 越冬寄主对烟粉虱生长发育的影响
分别取20~30头混合日龄MEAM1隐种烟粉虱雌成虫,用微虫笼固定于花椰菜、龙葵(Solanum nigrumL.)和辣子草(Ranunculus sieboldiiMiq.)供试寄主植株叶片背面,待产卵8 h后,移去微虫笼和烟粉虱成虫,在解剖镜下观察记录叶片初始卵量后,移至(25±1)℃、相对湿度(65±5)%、光照 14 h/10 h(L/D)的人工气候箱中(一恒 MGC-350HP)饲养,观察至成虫羽化,统计各供试寄主植物上烟粉虱的世代存活率和世代发育历期,每种寄主植物重复3次。
1.6 越冬隐种鉴定
在2017年1月,采集露地或温室大棚中栽培蔬菜和杂草上的烟粉虱成虫,并参照郑宇等[20]方法提取单头烟粉虱总 DNA,参照Shatters等[21]的方法进行隐种鉴定。MED隐种烟粉虱特异性引物为:上游引物 CTTGGTAACTCTTCTGTAGATGTGTGTT和下游引物 CCTTCCCGCAGAAGAAATTTTGTTC;MEAM1隐种烟粉虱特异性引物为:上游引物CTAGGGTTTATTGTTTGAGGTCATCATATATTC和下游引物 AATATCGACGAGGCATTCCCCCT。PCR反应条件为94℃预变性3 min,94℃变性30 s,64℃退火45 s,72℃延伸60 s,35个循环,72℃延伸 10 min。
1.7 统计分析
利用混合线性模型(linear mixed effect model)分析调查月份、调查地点、蔬菜生育期、栽培方式对蔬菜和杂草上烟粉虱越冬虫口密度的影响,模型中将田块以及寄主作为随机效应因子,以平均每片叶片上烟粉虱卵、若虫、成虫的数量作为响应变量。以 95%置信区间的重叠与否来检验同一地点、烟粉虱同一虫态虫口密度差异显著性以及低温下烟粉虱各虫态存活率之间的差异显著性。利用单因素方差分析方法(One-way ANOVA)比较龙葵、辣子草、花椰菜对烟粉虱世代存活率和发育历期的影响。对于在a=0.05水平上存在差异的处理效应,应用 Tukey法进行多重比较分析。所有分析和作图均在R语言软件上完成。
2 结果与分析
2.1 烟粉虱越冬寄主植物
调查农作物及农田杂草共 13科 31种,发现并记录到烟粉虱寄主植物6科16种(表1),分别为茄科(Solanaceae)4种、十字花科(Brassicaceae)2种、葫芦科(Cucurbitaceae)1种、豆科(Fabaceae)1种、菊科(Asteraceae)7种、唇形花科(Labiatae)1种。露地越冬寄主主要为花椰菜,温室大棚越冬寄主主要为花椰菜、番茄(Lycopersicon esculentumMill.)、茄子和黄瓜蔬菜以及龙葵、小飞蓬[Conyza canadensis(L.)Cronq.]、辣子草和胜红蓟(Ageratum conyzoidesL.)等阔叶类杂草。
2.2 烟粉虱在露地的越冬情况
福建省不同地区露地花椰菜上烟粉虱越冬虫口密度在不同调查月份(F1,27.0=148.6,P<0.001)、不同调查地点(F5,15.0=10.7,P<0.001)和不同生育期(F2,25.2=56.3,P<0.001)之间均存在显著差异(表2)。其中,1月份的越冬虫口密度显著高于 2月份;漳州龙海的越冬虫口密度显著高于其他地区(P<0.05),而其他地区之间的越冬虫口密度均差异不显著(P>0.05);花椰菜采后残株上的越冬虫口密度显著多于莲座期(t15.0=10.4,P<0.001)和花 球 形 成 期(t26.0=8.1,P<0.001),花球形成期的越冬虫口密度显著高于莲座期(t26.0=10.0,P<0.001)。在温度较低的2月份,南平建瓯、三明三元和将乐,仅零星发现卵和伪蛹;福州闽侯和龙岩新罗主要发现卵、高龄若虫和成虫;漳州龙海发现烟粉虱的各种虫态。
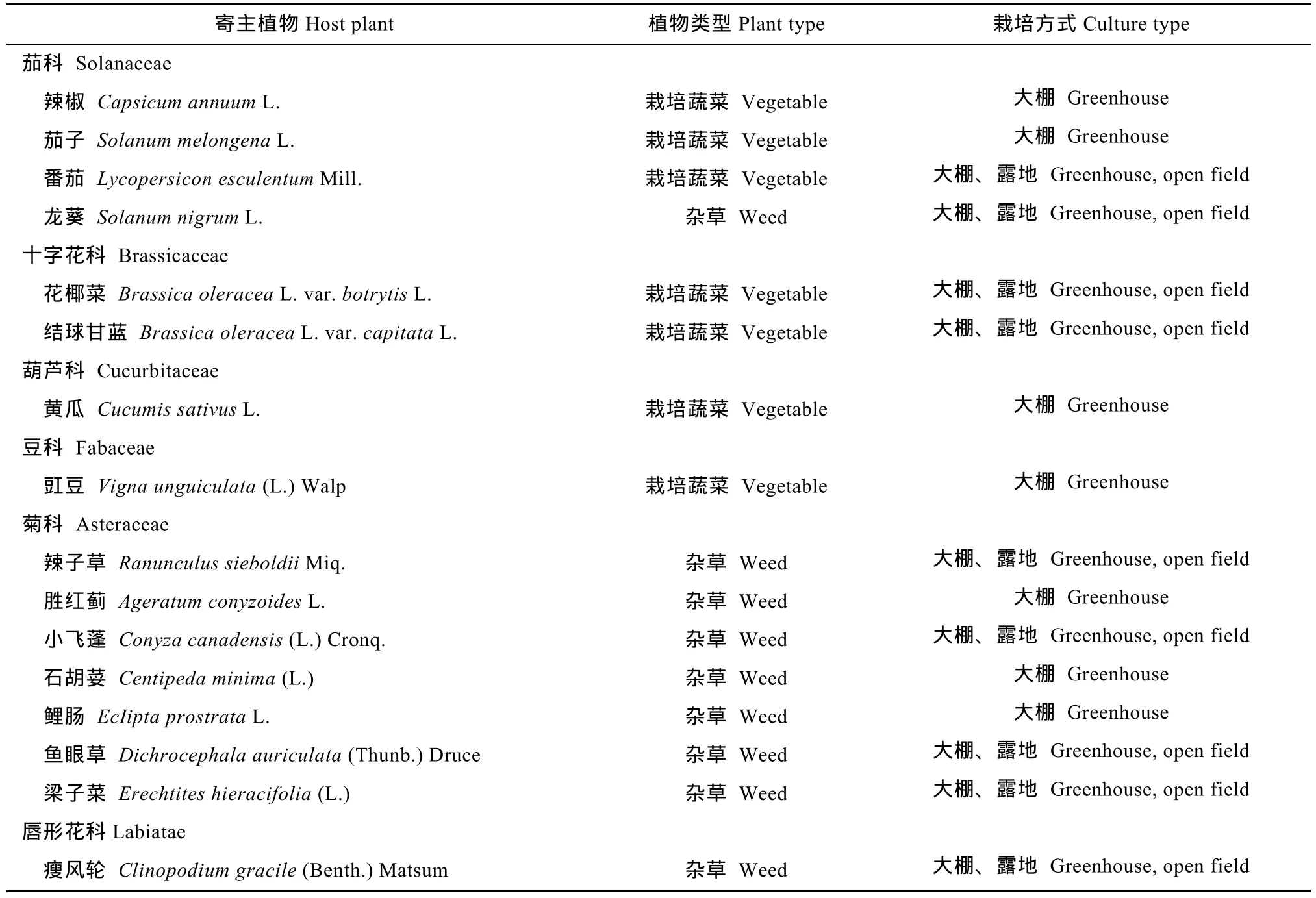
表1 福建省烟粉虱越冬寄主植物种类Table 1 Overwintering host plant species of Bemisia tabaci(Gennadius)in Fujian Province
2.3 烟粉虱在温室大棚中越冬情况
生产大棚烟粉虱越冬虫口密度显著低于休耕大棚(F1,10.2=20.4,P=0.001),两者之间的差异并不随着地区的改变而改变(F2,16.2=1.44,P=0.266)。在三明将乐的生产大棚内,莲座期和花球形成期的烟粉虱越冬虫口密度存在显著差异(F1,5=7.5,P=0.041),花球形成期烟粉虱卵、若虫和成虫数量分别为3.43粒·叶-1、1.45头·叶-1和0.29头·叶-1,分别明显高于莲座期的0.59 粒·叶-1、0.43 头·叶-1和0.08 头·叶-1(表3)。
2.4 温室大棚对烟粉虱越冬的影响
福建冬季极端低温条件下,蔬菜上烟粉虱越冬虫口密度在不同栽培方式下差异显著(F1,9=30.0,P<0.001),即露地烟粉虱越冬虫口密度显著低于温室大棚。栽培方式与不同调查地点之间没有交互作用(F1,9=2.36,P=0.158),表明在三明将乐和龙岩新罗地区均是温室大棚的烟粉虱越冬虫口密度显著大于露地(表4)。
2.5 烟粉虱在杂草上越冬情况
表5结果显示,栽培方式显著影响杂草上越冬的烟粉虱虫口密度(F1,18=95.5,P<0.001),即休耕温室大棚内杂草上烟粉虱越冬虫口密度显著高于棚外露地杂草。栽培方式与地区之间不存在交互效应(F2,18=1.0,P=0.371),即栽培方式间烟粉虱虫口密度的差异显著性不随调查地点的变化而变动。另外,在三明将乐地区,温室大棚外露地杂草上烟粉虱虫口密度与距离大棚的远近相关(χ2=6.6,df=2,P=0.038)。距离增大,虫口密度显著减小,温室大棚附近的露地杂草上虫口密度最大,而周围无温室大棚的露地烟粉虱虫口密度极低。
2.6 低温对烟粉虱存活的影响
由表6可知,在0℃、4℃和8℃低温下,处理6 h后,烟粉虱低龄若虫的存活率显著低于卵、高龄若虫和成虫;处理96 h后,成虫的存活率显著低于
卵和高龄若虫。卵和高龄若虫的存活率差异程度与低温强度及持续时间相关,在8℃低温下处理 48 h至144 h,卵的存活率显著高于高龄若虫;在4℃低温下,处理6 h至144 h,高龄若虫的存活率显著高于卵;而在0℃低温下,高龄若虫的存活率在6 h至24 h显著高于卵,在48 h至144 h两者差异不显著。
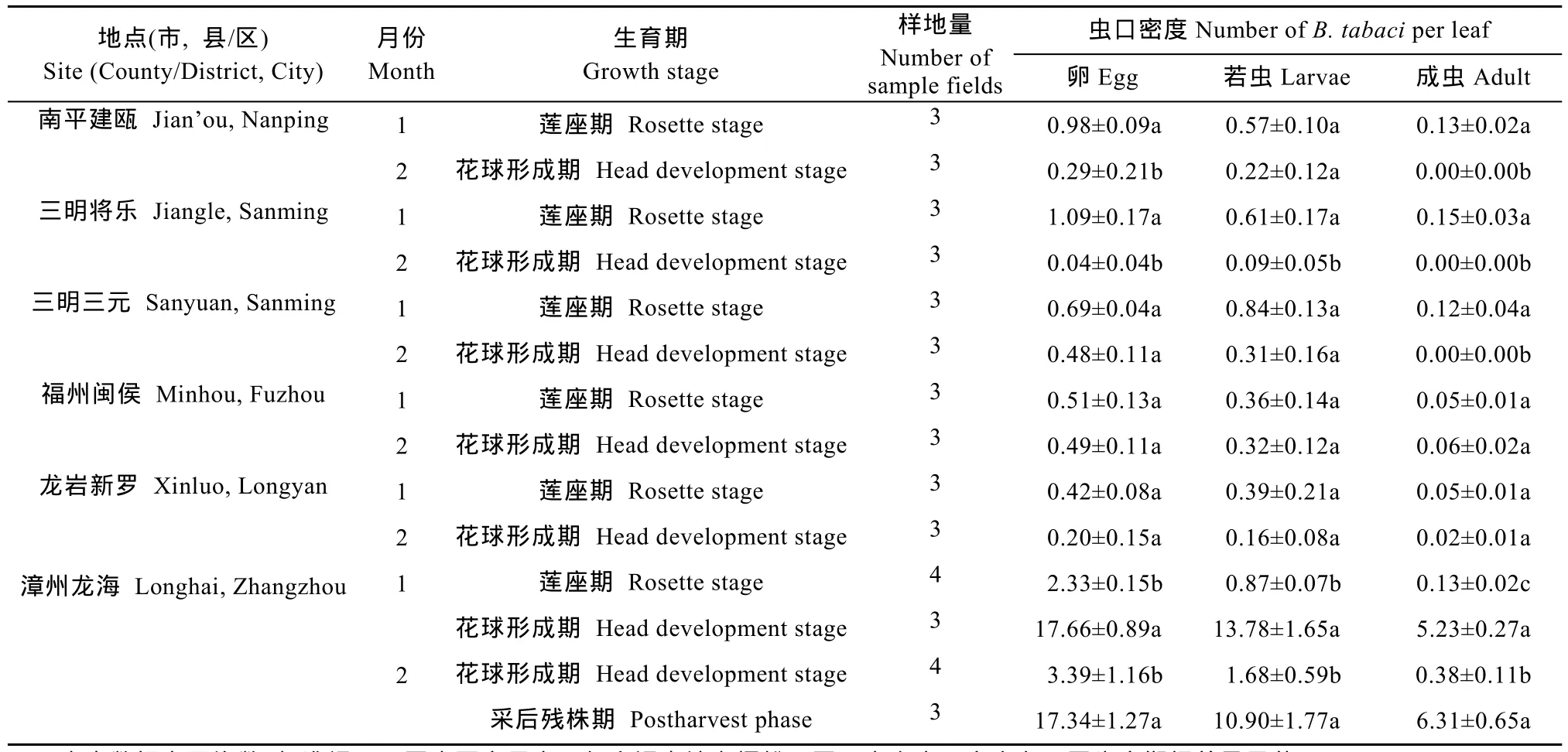
表2 福建省不同地点露地花椰菜上烟粉虱越冬虫口密度Table 2 Density of overwintering Bemisia tabaci(Gennadius)on Brassica oleracea L.var. botrytis L.grown in open fields in different investigated sites in Fujian Province
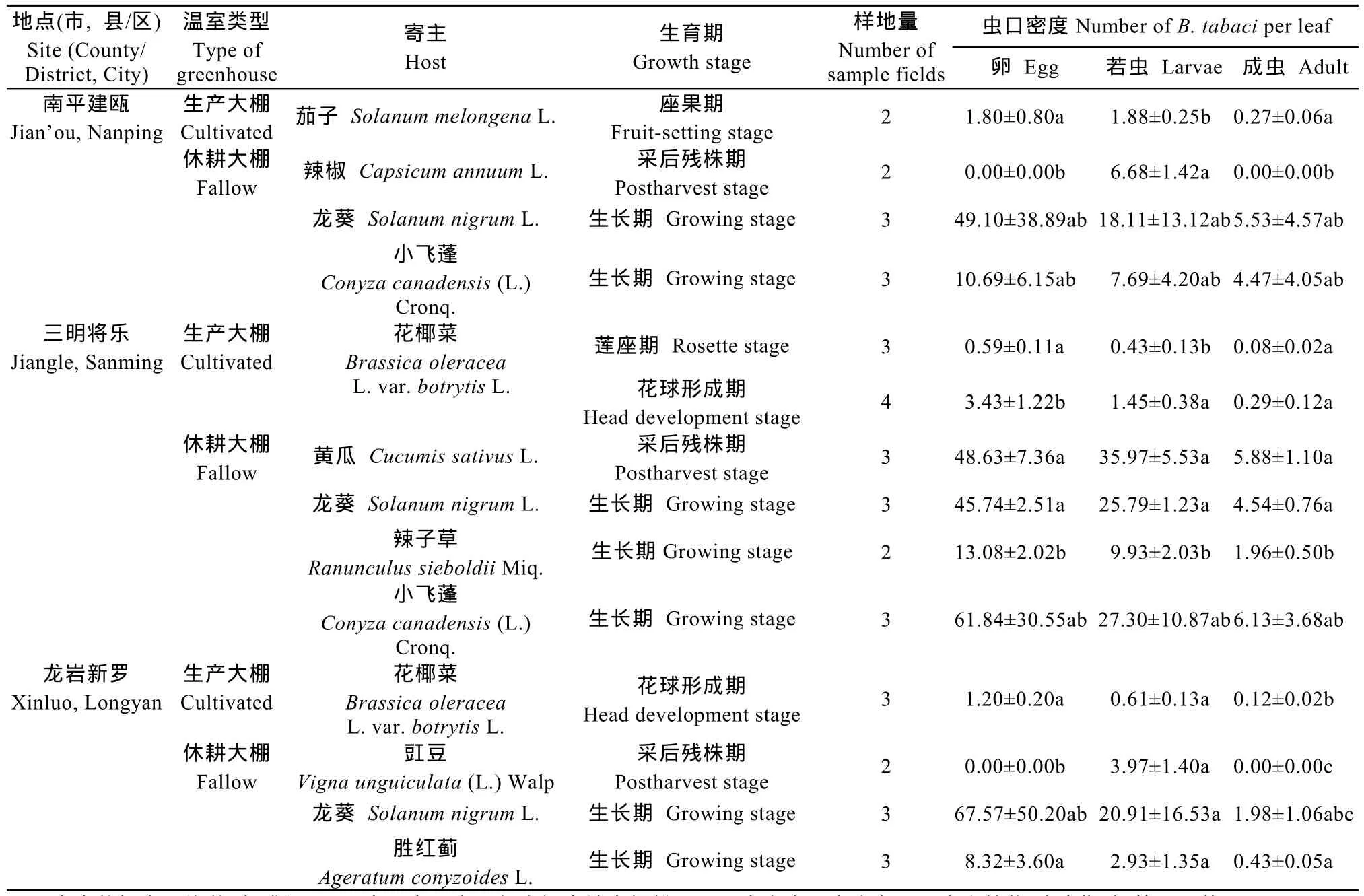
表3 福建省不同地点温室大棚中烟粉虱越冬虫口密度Table 3 Density of Bemisia tabaci(Gennadius)overwintering in greenhouses in different investigated sites in Fujian Province
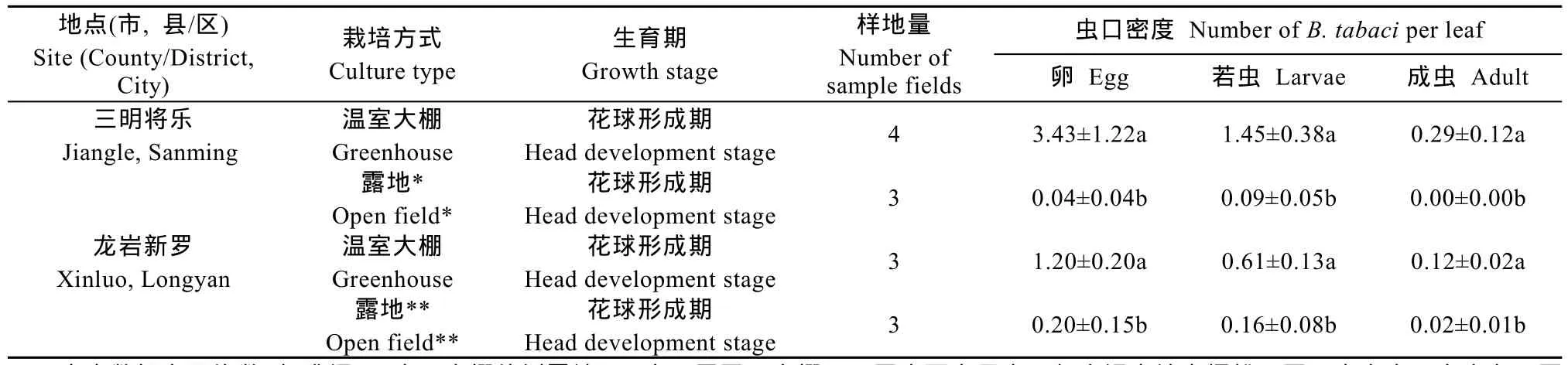
表4 福建省不同地点温室大棚和露地花椰菜上烟粉虱越冬虫口密度Table 4 Density of Bemisia tabaci(Gennadius)overwintering on Brassica oleracea L.var. botrytis L.grown in greenhouses and open fields in different investigated sites in Fujian Province
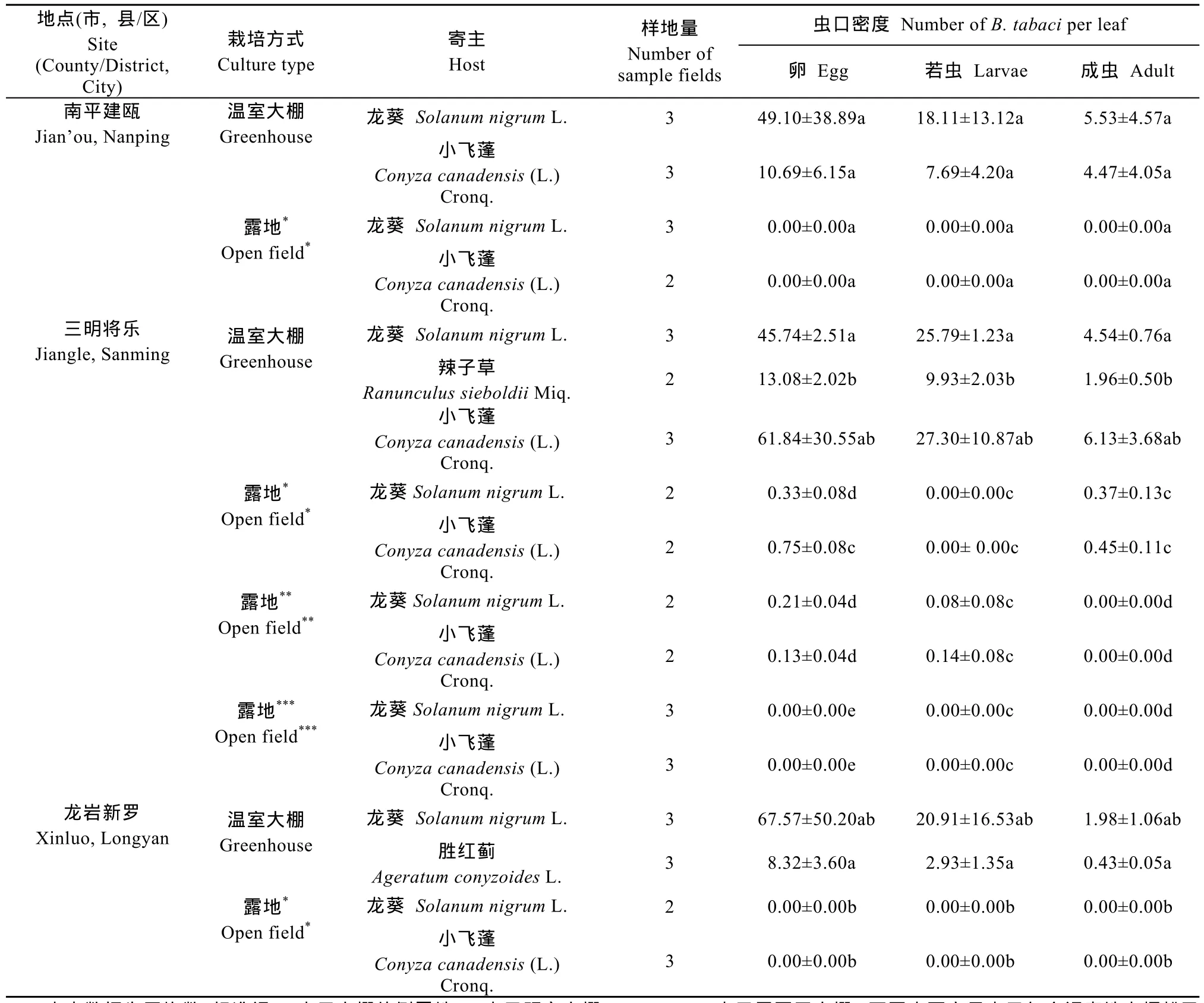
表5 福建省不同地点休耕温室大棚和露地杂草上烟粉虱越冬虫口密度Table 5 Density of Bemisia tabaci(Gennadius)overwintering on weeds grown in fallow greenhouses and open fields in different investigated sites in Fujian Province

表6 低温对烟粉虱各虫态存活的影响Table 6 Effect of low temperatures on the survival rate of Bemisia tabaci(Gennadius)at various life stages
2.7 越冬寄主对烟粉虱世代发育的影响
在室内25℃适温条件下,烟粉虱在花椰菜、龙葵和辣子草上的世代存活率分别为83.7%、79.3%和81.6%,差异不显著(F2,6=0.127,P=0.883);世代发育历期分别为18.9 d、16.7 d和18.8 d,差异显著(F2,6=22.0,P=0.002)。其中,烟粉虱在龙葵上的发育历期显著短于在花椰菜和辣子草,而在花椰菜和辣子草上的发育历期差异不显著(表7)。

表7 烟粉虱在不同越冬寄主上的生长发育Table 7 Development of Bemisia tabaci(Gennadius)on different overwintering plants
2.8 越冬烟粉虱隐种
由表8可知,福建省越冬烟粉虱隐种有MEAM1隐种和MED隐种,总数量分别为105只和111只,而温室大棚中,两隐种的数量分别为57只和111只。其中,漳州市龙海市东园镇露地花椰菜和番茄上仅发现 MEAM1隐种;三明市将乐县光明乡温室大棚黄瓜上仅发现 MED隐种;其他地区发现两种隐种混合发生,其中龙岩市新罗区东肖镇和南平市建瓯市小松镇温室大棚内MED隐种占优势,而福州市闽侯县南通镇和三明市三元区中村乡温室大棚内MEAM1隐种占优势。
3 讨论与结论
目前已记录到烟粉虱寄主植物 600多种[22-24]。何玉仙等[25]前期调查记录福建省福州、漳州农田烟粉虱寄主植物 17科 62种,主要包括十字花科、菊科、豆科、茄科、葫芦科、锦葵科(Malvaceae)等植物。越冬寄主是特指在冬季给昆虫提供营养物质和居住场所使其能安全越冬的植物。周国珍等[13]通过调查发现,湖北省烟粉虱露地越冬寄主主要有荔枝草(Salvia plebeiaR.Br.)、小飞蓬、一年蓬[Erigeron annuus(L.)Pers.]、益母草[Leonurus artemisia(Laur.)S.Y.Hu]、苦苣菜(Sonchus oleraceusL.)、马蹄金(Dichondra repensForst.)和金银花(Lonicera japonicaThunb.),且以荔枝草为主。崔洪莹等[11]发现露地上的烟粉虱寄主有荔枝草、白菜[Brassica pekinensis(Lour.)Rupr.]、甜菜(Beta vulgarisL.)、菠菜(Spinacia oleraceaL.)、莴苣(Lactuca sativaL.)、萝卜(Raphanus sativusL.)和一年蓬,而温室大棚中主要有辣椒、酸模(Rumex acetosaL.)、婆婆草(Bidens pilosaL.)、荠草[Brassica juncea(L.)Czern.and Coss.]、繁缕[Stellaria media(L.)Cyr.]和番茄,露地上荔枝草的虫量最大,大棚中婆婆草的虫量最大。本研究同样发现蔬菜作物和杂草可作为烟粉虱的越冬寄主,露地上花椰菜的虫量最多,大棚中蔬菜和杂草上均具有较多虫量,尤其是休耕大棚前茬作物残株上的虫量很大,可以扩散到棚内大量生长的龙葵、胜红蓟、辣子草、小飞蓬等阔叶类杂草上。结合烟粉虱在龙葵和辣子草上的生长发育表现(表7),我们认为这些阔叶类杂草是福建地区冬季烟粉虱重要的越冬替代寄主。Muñiz[14]同样发现龙葵是烟粉虱的适宜寄主。此外,阔叶类杂草锦葵(Malva parvifloraL.)、荠菜(Capsella bursa-pastorisL.)和田芥[Brassica kaber(DC)]也是适宜烟粉虱生长发育和繁殖的越冬寄主[14]。

表8 福建省不同地区冬季烟粉虱隐种发生情况Table 8 Number of cryptic species of Bemisia tabaci(Gennadius)overwintering in Fujian Province
低温的强度和持续时长与露地烟粉虱越冬数量密切相关[10-11]。崔洪莹等[11]和Lin等[26]发现随着温度降低,烟粉虱成虫在低温暴露下的致死中时间不断缩短。本研究亦发现随着温度降低及持续时间延长,烟粉虱各虫态的存活率呈下降趋势。2017年2月的温度较1月低,2月份蔬菜上的虫口密度也相应地显著少于 1月份蔬菜上的虫口密度。另外,我们还发现卵和高龄若虫更耐低温,在温度更低的三明和南平地区露地蔬菜上烟粉虱以卵和伪蛹越冬。这与周国珍等[13]、董国等[12]的研究发现相符。但是,除了湖北仙桃的荔枝草和江西南昌的一年蓬以烟粉虱卵为主外,崔洪莹等[11]发现在湖北、浙江和江西的露地蔬菜和杂草上,烟粉虱主要以成虫越冬。
越冬北界是烟粉虱越冬研究中重要的科学问题,也得到了较多的关注[10-13],而且多数研究选择接近中亚热带北界的地区开展试验。例如,崔洪莹等[11]认为湖北省仙桃市、江西省南昌市和浙江省金华市可以作为烟粉虱的自然越冬北界。陈春丽等[9]研究预测,12月下旬,我国除福建、广东、广西、云南、海南、台湾等省有烟粉虱田间存在,其他省的烟粉虱在露地基本上消退。实地调查发现烟粉虱在广东、福建等南方地区的一些露地蔬菜大田可以安全越冬[8-9]。本研究调查结果亦证实,在福建省南平市建瓯市小松镇和三明市三元区中村乡以南地区,烟粉虱可在露地越冬。本研究还发现,在南亚热带北界附近的福州闽侯和龙岩新罗露地蔬菜上烟粉虱各虫态均可安全越冬,但主要是卵、高龄若虫和成虫,且越冬虫口密度较小;而在南亚热带的漳州龙海露地蔬菜上烟粉虱各虫态均可安全越冬,且越冬虫口密度较大,可周年危害。随着全球气候变暖,在南亚热带北界附近的区域,烟粉虱越冬的数量可能会率先出现积极的响应,应予以关注。
尽管烟粉虱在露地上的越冬界限与冬季的低温等温线分布范围关系密切[9,11],但温室大棚对烟粉虱成功越冬具有积极的保护作用。王勇等[10]发现在越冬北界分布线以北的江苏江淮地区,烟粉虱可在双膜覆盖的大棚和日光温室等保护地设施中越冬。甚至在高纬度地区如哈尔滨和辽宁的温室大棚中,烟粉虱仍可以伪蛹越冬[27-28]。本研究发现福建各地区温室大棚蔬菜和杂草上烟粉虱各虫态均可安全越冬,且温室大棚内烟粉虱虫口密度远高于棚外露地,休耕温室大棚内烟粉虱虫口密度明显高于生产大棚。由于温室大棚中越冬有助于烟粉虱以更大的虫源基数扩散到田间进行为害,是烟粉虱周年循环为害的一个重要环节[10,26]。因此,建议在冬季休耕期要及时清除大棚内作物残株及阔叶杂草以降低越冬烟粉虱虫量。
在福建省,MED隐种烟粉虱于2013年在局部地区被发现,2014年于多个地区被发现且占比不断上升,但截至2014年福建省烟粉虱仍以MEAM1隐种为主[7]。Chu等[29]证实冬季广泛使用的温室大棚有助于增加烟粉虱的遗传多样性,促进MED隐种烟粉虱的成功定殖。温室大棚的建立可能是MED隐种烟粉虱在福建省定殖的重要因素之一[7]。本研究表明MED隐种烟粉虱可以在大棚中越冬。由于MED隐种对杂草寄主的适应性较 MEAM1隐种烟粉虱更高[14],在休耕大棚中大量生长的杂草可能也会促进 MED隐种竞争取代MEAM1隐种。
本文主要关注蔬菜高强度生产的农田生态系统,仅选取了典型作物即蔬菜和杂草进行调查,而园艺花卉中也有烟粉虱喜好的寄主植物[26,30],在花卉与蔬菜毗邻种植区域开展越冬调查,有助于进一步明确烟粉虱越冬规律。冬季烟粉虱MEAM1、MED隐种的竞争替代对翌年种群建立具有重要作用[29],加大不同栽培方式下烟粉虱取样和鉴定数量,对比MEAM1、MED隐种的耐寒性以及对常见越冬杂草的适应性,可以增进对两者竞争替代规律的认识。
综上,温度是影响烟粉虱越冬的最重要因素,烟粉虱可以在福建露地越冬。温室大棚的保温作用有助于烟粉虱越冬,尤其在休耕大棚内,前茬作物残株和阔叶杂草上留有大量各虫态烟粉虱,成为翌年春季扩散到露地为害的重要虫源。温室大棚的发展建设可能有助于MED隐种烟粉虱的定殖扩散。建议清除田间蔬菜残株和阔叶类杂草,压低烟粉虱的越冬虫口基数。
猜你喜欢
上海蔬菜(2022年6期)2022-12-31
农业工程学报(2022年10期)2022-08-22
天津农业科学(2022年5期)2022-05-31
今日农业(2021年3期)2021-12-05
环境昆虫学报(2021年5期)2021-10-19
今日农业(2021年9期)2021-07-28
今日农业(2021年4期)2021-06-09
天津农林科技(2021年2期)2021-04-16
饲料博览(2020年11期)2020-12-30
植物保护(2018年5期)2018-12-05
