
野生多花黄精种群表型变异研究
2019-06-12 07:16:54周新华曾平生武晓玉姚甲宝熊光康李家彧
中南林业科技大学学报 2019年6期
周新华,曾平生,武晓玉,姚甲宝,熊光康,李家彧
(中国林业科学研究院亚热带林业实验中心,江西 分宜 336600)
遗传多样性是指地球上所有生物所携带的遗传信息的总和,它体现了物种基因的丰富程度,是研究物种多样性及生态系统多样性的基础[1-2]。生物种内遗传多样性研究有多种遗传标记方式,包括表型、细胞学、生化和分子标记4 大类,表型相较于其他三种方式可以更直观地揭示生态系统多样性和生物多样性[3-5]。生物的表型变异是基因在环境因子的影响下不同表达的结果,是生物的遗传受制于环境条件的遗传表征是遗传多样性的主要研究内容[6]。利用表型性状研究种群的遗传多样性具有简便、快速和节省费用等优点,尽管表型性状不完全等同于基因位点,但表型变异是基因型与环境因子共同作用的结果,更具有综合性和实用性[7]。目前,国内外学者在表型变异研究方面已在木本植物、藤本植物和草本植物[8-23]开展了相关研究,为相关植物资源的保护利用或遗传育种奠定了坚实的基础。
多花黄精Polygonatum cyrtonema Hua 属百合科黄精属的多年生草本植物,是药食两用的大宗传统中药材,具有较高的观赏、药用和经济价值[24-25]。多花黄精主要分布在贵州、湖南、四川、湖北、河南、浙江、广东、福建、安徽、江西等南方省份,生长在海拔500~2 100 m 的林下、灌丛或山坡阴处[25]。多花黄精植株形态优美,根茎含有特有的黄精多糖、皂苷等药用活性物质,故在生产实践中可用作园林绿化草本植物,也是当今林下经济发展中重要的林下药材栽培品种。然而,多花黄精之前的研究主要集中在种子的萌发[26]、无性组培育苗[27-30]、块茎药用成分分析[31-32]和人工林下栽培[33-35]等方面,但作为遗传育种工作基础的地理种源、群落和形态变异暂未见相关研究报道。以不同地理种群的野生多花黄精为研究对象,从全国分布区内选取了具有代表性的9 个样点,系统测定与其生长表型形状密切相关的12 个性状,运用统计学分析方法分析其相关指标,阐明不同地理种群间及其自身内部的变异规律,为多花黄精中药材产业利用、开发和保护提供支撑。
1 材料与方法
1.1 样品采集和数据收集
在对中国植物志、中国数字植物标本馆等文献资料检索和野外实地调查的基础上,作者于2014—2017年的10—11月间进行多花黄精植株的野外收集工作。野生植株的收集地点分别位于江西省的鹰潭市(JXYT)、新余市(JXXY),湖北省的宜昌市(HBYC),浙江省的丽水市(ZJLS),安徽省的黄山(AHHS),贵州省的凯里市(GZKL),广西省的桂林市(GXGL),湖南省的衡阳市(HNHY),四川省的南充市(SCNC),共计9个地理种群的135 株。植株收集要求生长年限5 a以上,单株无病虫害,生长态势正常,无明显的缺陷,同时每个地理种源收集的单株要求间隔距离大于30 m 以上,种群保有单株数量在100 株以上。由于野生多花黄精生长在原生环境较好的高山荫蔽处,植株收集工作比较困难,故植株收集时要求尽可能多地收集多花黄精单株,9 个种群的植株收集数量至少保证每个种群在15 株以上。通过查阅中国自然资源数据库(http://www.data.ac.cn/zy/show/shi3.asp)获得相关调查区域的气候资源数据,并在调查时记录各点的地理经纬坐标和海拔等数据,所获得数据具体见表1。

表1 野生多花黄精9个种群的地理位置及主要气候因子Table 1 Geographical locations and main conditions for nine populations of Polygonatum cyrtonema
1.2 表型性状测定方法
每个种群选取长势良好的健康单株5 株,统计其叶片数,并利用电子游标卡尺测定每株的叶长、叶宽、地径、果实纵径、果实横径,测量时精确到0.01mm,计算叶片的长宽比和果实的横纵比;利用LI-3000C 便携式叶面积测定仪(LI-COR,Lincoln,USA)测定每株每片叶的叶面积,并计算其全株叶面积,测量精确到0.01 cm2;利用不锈钢直尺测定每株株高,测量精确到0.1 cm,每个野生种群测量5 株,重复3 次,每个单株的叶长、叶宽、叶柄长、果实纵径、果实横径、叶面积等指标按单株取平均值。每个种群随机选取100 粒种子,带回实验室用电子分析天平测定其质量,换算成千粒重,重复5 次。
1.3 数据处理和统计分析
数据的整理和统计分析采用社会统计学分析软件SPSS25.0 进行。对所选取的12 个表型形状指标进行9 个地理种群间的单因素方差分析和多重比较分析,比较表型分化系数等得出种间表型性状变异特征[36];运用表型性状的平均值和标准差,计算出表型性状的变异系数,然后用变异系数来衡量多花黄精种群内的变异水平;运用相关分析来分析各表型性状与生态因子之间的相互关系;运用聚类分析,计算出不同种群遗传距离的大小。所有数据在统计分析时,均确保在同一量纲上进行。
2 结果与分析
2.1 野生多花黄精生长表型在种群间和种群内的变异
野生多花黄精生长表型指标在种群间和种群内的差异结果见表2。分析结果可知,所选取的12 个表型性状除果实横纵比外,其余性状指标无论是在种群间还是种群内都存在显著(P <0.05)或极显著(P <0.01)的差异,这就说明不管是在种群间还是在种群内,野生多花黄精大部分生长表型性状指标存在变异。9 个不同种群的12 个生长表型性状指标的平均值、标准差及多重比较结果(表4)表明,除果实横纵比外的各表型性状间均存在显著差异。其中,江西新余种群的叶片宽最小、叶片长宽比最大,其值分别为30.19 mm 和5.60,与其它几个种群间的差异较为显著;湖北宜昌种群的果实横径最小,其值为6.88 mm,与四川南充种群间差异极显著;浙江丽水种群的叶片数、叶片长宽比、全株叶面积、地径和株高值最小,其值分别为10.67、3.08、364.61 cm2、4.56 mm 和57.11 cm,叶片宽值最大,其值为37.56 mm;安徽黄山种群的叶片长、叶面积和全株叶面积值最大,其值分别为178.99 mm、49.43 cm2和897.33 cm2,果实纵径值最小,其值为6.28 mm;贵州凯里种群的叶片数、株高最大,其值分别为20.67、105.00 mm,显著大于江西鹰潭、江西新余、浙江丽水和广西桂林种群;湖南衡阳种群的叶片长、叶面积和种子千粒重值最小,其值分别为112.16 mm、27.16 cm2和12.23 g;四川南充种源的地径、果实横径、果实纵径和种子千粒重均最大,其值分别为10.66 mm、9.76 mm、11.11 mm和16.43 g,均显著高于大多数种群。
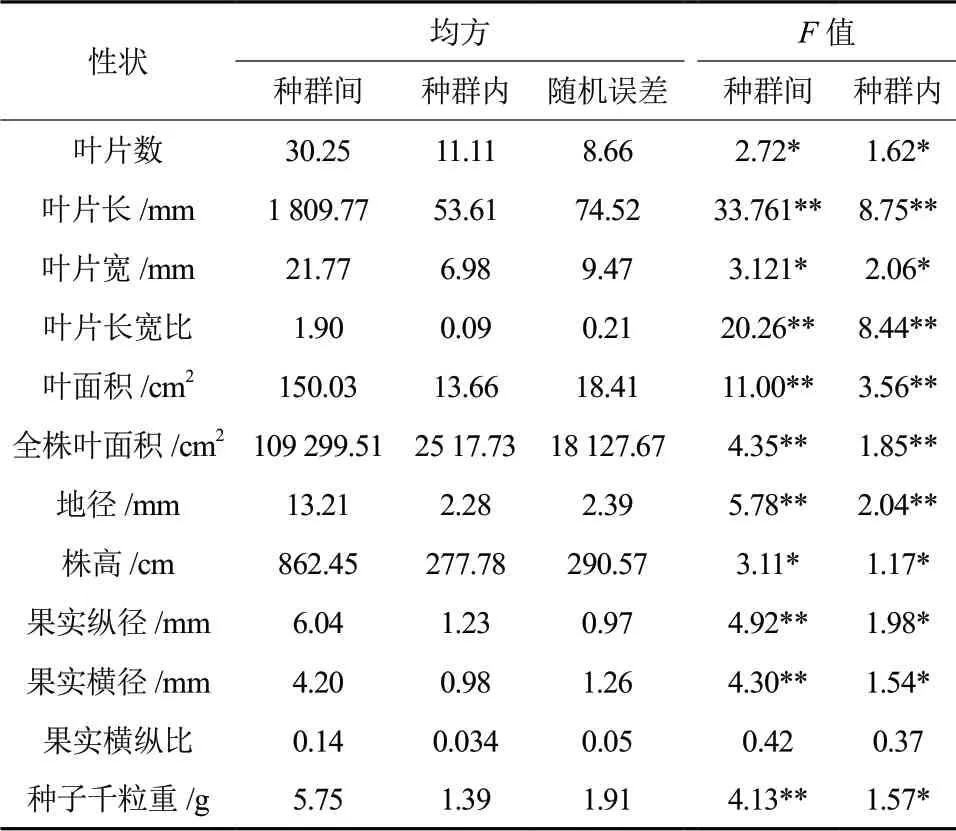
表2 野生多花黄精各种群间和种群内全株、叶片、果实、种子表型性状的方差分析结果Table 2 Variance analysis of phenptypic traits of whole plant,leaf,fruit and seeds among and within Polygonatum cyrtonema Hua populations
9 个不同种群野生多花黄精的12 个表型性状指标在种群内方差分量、种群间方差分量和分化系数情况见表3。数据显示:就各性状的平均方差分量百分比来看,种群内占36.24%、种群间占15.37%、随机误差占48.39%,种群内的平均方差分量百分比远大于种群间的平均方差分量百分比,这说明生长表型性状的种群内变异远大于种群间的变异;就各性状的平均表型分化系数来看,种群内贡献值是71.16%、种群间贡献值是28.84%,这就说明在野生多花黄精的表型变异中,种群内部的变异远远大于种群间的变异,即变异主要存在不同种群内部。
2.2 不同地理种群野生多花黄精表型性状变异特征分析
不同地理种群野生多花黄精的各表型性状在种群内的平均变异系数为12.16%,变异幅度为4.76%~23.63%(表5)。叶片数、全株叶面积、地径、株高和种子千粒重等5 个指标的变异系数都大于16%,远超所有指标的变异系数平均值;叶片长、叶片宽、叶片长宽比、单叶面积、果实纵径、果实横径和果实横纵比等7 个表型性状的变异系数均不超过12.16%。另外,进一步分析不同地理种群野生多花黄精所有指标的平均变异系数可知,江西新余、浙江丽水和广西桂林3 个种群的变异系数较大,其值分别为14.45%、15.99%和19.76%,远大于平均变异系数12.16%,这就表明了新余、丽水和桂林种群的表型变异和多样性均较为丰富;湖南衡阳和四川南充2 个种群的变异系数较小,其值分别为8.21%和6.39%,远小于平均变异系数12.16%,说明这2 个地理种群的表型变异较弱,多样性也较低;江西鹰潭、湖北宜昌、安徽黄山和贵州凯里4 个种群的变异系数幅度为10.15%~12.94%,比较接近平均变异系数12.16%,说明这4 个地理种群的多样性大体一致。

表3 野生多花黄精表型性状的方差分量及其种群间表型分化系数Table 3 Variation components and phenotypic differentiation coefficients among Polygonatum cyrtonema Hua populations
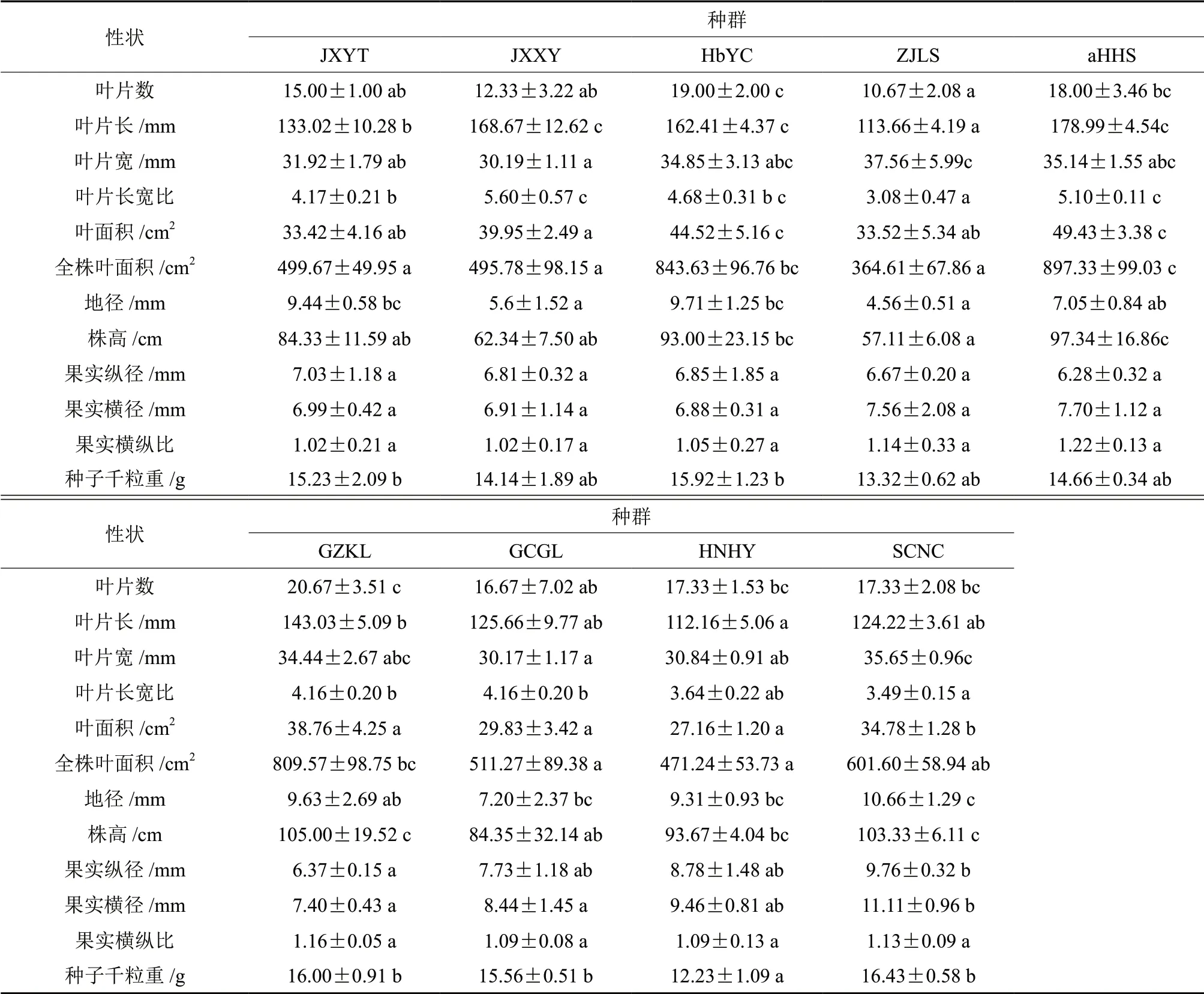
表4 野生多花黄精9个种群表型性状的平均值、标准差及多重比较Table 4 The mean value,standard deviation and multiple comparison of phenotypic traits of nine populations in Polygonatum cyrtonema Hua
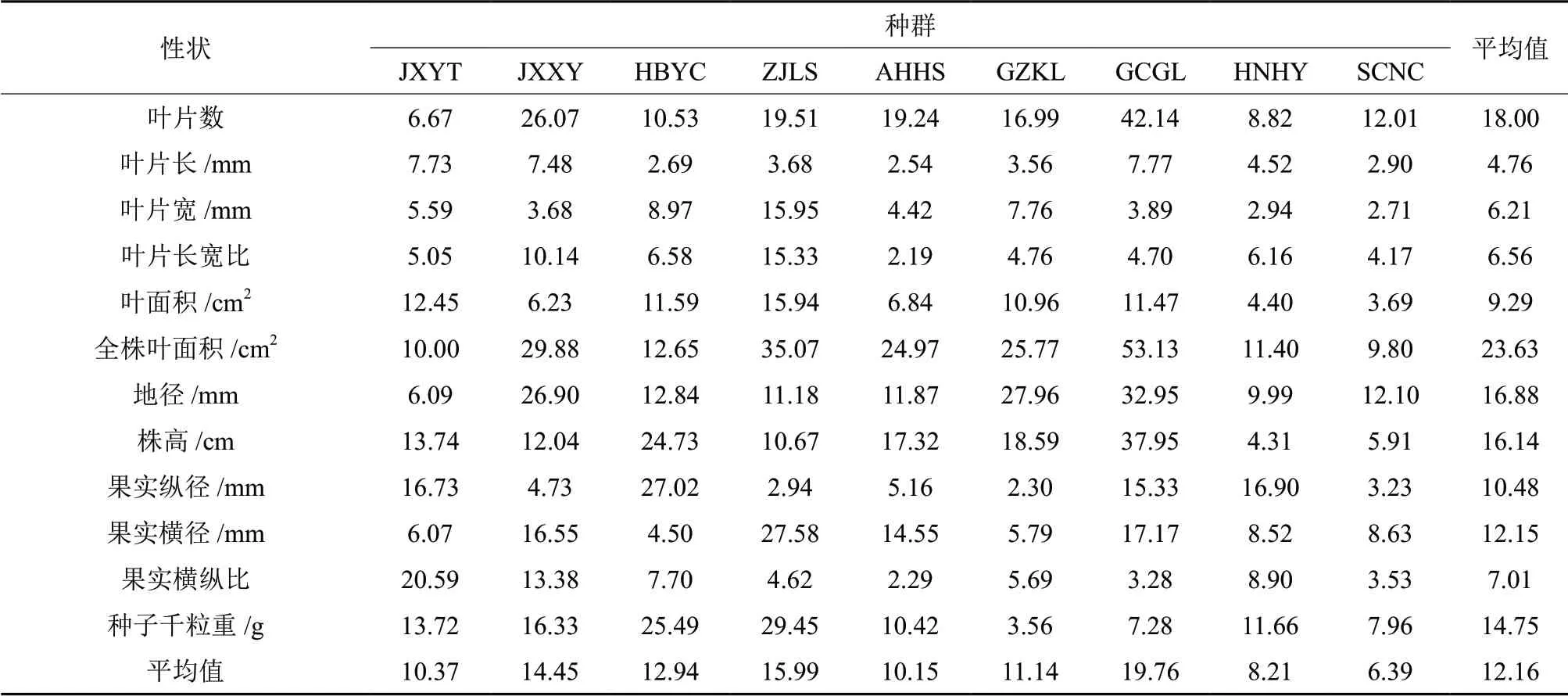
表5 野生多花黄精种群表型性状变异系数Table 5 Variation coefficients of phenotypic traits in Polygonatum cyrtonema Hua populations
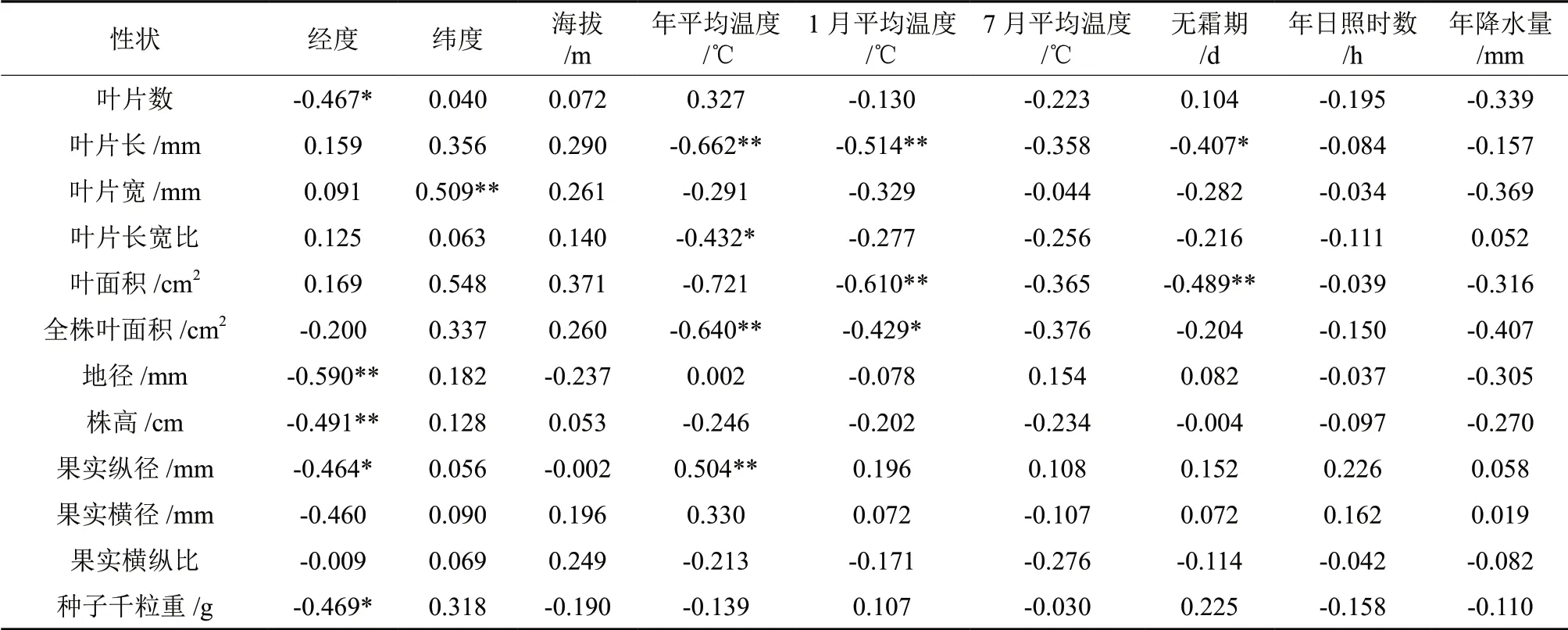
表6 野生多花黄精13个表型性状与生态地理因子的相关系数Table 6 The correlation coefficient of Polygonatum cyrtonema Hua 13 phenotypic traits and geographical factor
2.3 不同野生多花黄精种群表型性状的地理变异
野生多花黄精全株、叶片、果实和种子12 个表型性状与采集地点的生态地理因子的相关分析见表6。叶片数、地径、株高、果实纵径和种子千粒重与经度均呈现显著或极显著的负相关关系,说明在经度较高的地区多花黄精拥有更少的叶片数,更小的地径、株高、果实纵径和种子千粒重;叶片宽与纬度间呈现极显著的正相关关系,说明随着纬度的增大叶片宽呈现下降趋势;叶片长、叶片长宽比和全株叶面积年平均温度间呈现显著或极显著的负相关关系,果实纵径与年平均温度呈现极显著的正相关关系,说明温度越低叶片长、叶片长宽比和全株叶面积越大,果实纵径越小;叶片长、叶面积和全株叶面积与1月平均温度呈现极显著的负相关关系,说明1月份平均温度越高,叶片长、叶面积和全株叶面积越小;叶片长和叶面积与无霜期呈现显著或极显著的负相关关系,说明无霜期越长,叶片长和叶面积越小。12 个表型性状均与海拔、7月平均温度、年日照时数和年降水量无显著相关关系,说明这些地理生态因子对多花黄精表型性状的影响不大。
2.4 不同地理种群野生多花黄精表型的聚类分析
利用所选取的12 个表型形状,采用欧氏距离法对9 个野生多花黄精种群进行聚类分析,聚类结果见图1。从种群关系聚类树状图可以看出,9 个野生多花黄精种群明显可以分为3类,江西鹰潭、广西桂林、湖南衡阳和四川南充种群归为第Ⅰ类,江西新余和浙江丽水种群归为第Ⅱ类,河北宜昌、贵州凯里和安徽黄山种群归为第Ⅲ类。仔细分析发现,在地理分布上相近的种群反到没有能聚为一类,因此,笔者将9 个野生多花黄精种群的12 个表型性状的Eucidean distances 与地理距离进行相关分析,相关分析数据显示欧氏距离与地理距离之间的相关性不显著(R=0.162,P=0.421)。这可能与野生多花黄精的自然生长环境有关,多花黄精喜欢阴凉、潮湿的生长环境,一般分布在林下、灌木下和山坡阴处,这种选择性的生长环境与地理位置的连续改变引起的生境改变相关系不大。这说明小生境的影响大于大气候对多花黄精表型变异的影响。
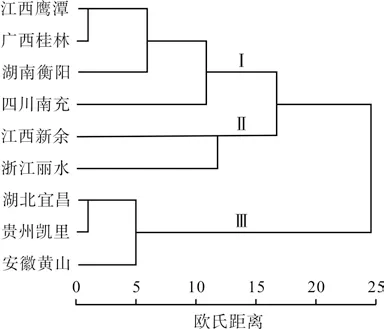
图1 基于12 个表型性状的野生多花黄精种群关系树状图Fig.1 Dendrogram of wild Polygonatum sibiricum population based on 12 phenotypic traits
3 讨 论
3.1 不同地理种群野生多花黄精表型变异来源
通过对野生多花黄精9 个地理种群的叶片数、叶片长、叶片宽、叶片长宽比、单叶面积、全株叶面积、地径、株高、果实纵径、果实横径、果实横纵比和种子千粒重等12 个表型性状的深入研究可以发现,野生多花黄精的这些表型性状除果实横纵比外的性状均在种群间和种群内存在显著或极显著的差异,这与紫茉莉[36]、肉苁蓉[37]、丹参[38]、黄芩[39]等药用植物的研究结果相似,这是不同地理种群的野生多花黄精自身遗传基因和所处生态环境共同作用的结果。种群间表型分化研究结果表明,种群内的变异远大于种群间的变异,种群内的变异是野生多花黄精表型变异的主要来源。12 个表型性状的变化幅度为17.19%~44.41%,平均分化系数为28.84%,种群间的变异仅贡献28.84%,远远小于种群内的变异贡献71.16%,引起这一现象的原因可能有两个:其一是9 个野生多花黄精种群在地理分布上相隔距离较远,除江西新余和江西鹰潭种源是同一个省份的,相隔距离为200 km 左右,其它种群均分布均在不同省份,相隔距离均大于江西新余和鹰潭的距离,这就导致不同种群间在基因交流时受阻于距离;另一个原因是同一种群的野生多花黄精生长在同质的生境条件下,这样就导致了同一种群内的多花黄精分布较为集中,种群内部的基因交流就相对容易实现。因此,基于上述原因,野生多花黄精种群内的遗传分化较种群间的遗传分化更容易实现,这就说明了多花黄精表型变异主要来源于种群内部。
3.2 不同地理种群野生多花黄精表型变异特征
变异系数又称离散系数,是衡量测量值变异程度的一个统计量,其值越大,则测量值的变异程度也就越大,则对应的物种多样性也就越丰富。对9 个野生多花黄精种群的叶片数、叶片长、叶片宽、叶片长宽比、单叶面积、全株叶面积、地径、株高、果实纵径、果实横径、果实横纵比和种子千粒重等12 个表型性状的变异系数深入研究发现,所有表型性状的变异系数幅度在4.76%~23.63%之间,平均变异系数为12.16%,变异系数的变化范围较大,说明多花黄精种群内表型性状的变异程度较高。种群内的变异程度也高,则说这一种植物种群内可能存在的遗传变异较大,同时也能更好地适应生存环境的改变。
野生多花黄精叶片数、全株叶面积、地径、株高和种子千粒重等5 个表型性状的变异系数分 别 为18.00%、23.63%、16.88%、16.14% 和14.75%,远大于平均变异系数,这5 个指标的稳定性排名为种子千粒重>株高>地径>叶片数>全株叶面积,这与滇重楼[40]和兰州百合[41]的种子、叶片和茎的相关研究性状是一致的,这就表明野生多花黄精种子的稳定性最高,茎的稳定性其次,叶片的稳定性最差。
3.3 不同地理种群野生多花黄精表型变异规律性
植物在不同的生态环境中,往往会以不同的形态来应对以适应,这对提高该物种的抗逆性和适应环境的能力是非常有利的。以往在许多植物群落的研究中常常会发现,植物的表型性状随生态地理因子的改变而出现一些规律性的变化。本研究发现,纬度、经度、年平均温度、1月平均温度和无霜期与多花黄精表型性状间存在一定的显著相关性,且而海拔、7月平均温度、年日照时数和年降水量与多花黄精表型性状间无显著相关性,这可能与多花黄精特殊喜阴的林下生长环境有关。随着经度的增大,野生多花黄精的叶片数、地径、株高、果实纵径和种子千粒重均减小,说明适当经度对多花黄精茎叶的营养生长和果种的生殖生长均有明显的促进作用,这与本次试验调查中贵州凯里和四川南充两个种群茎叶、果实、种子的生长表型情况一致。果实横径和果实纵径与9 个生态地理因子均无显著相关性,这说明生态环境因子对多花黄精果实表型的影响不大,而决定果实表型的因素主要是其自身的遗传因素。
3.4 野生多花黄精保护策略
多花黄精作为我国传统大宗药食两用的药用植物品种,在人们健康产业的不断发展中,传统制药、药膳保健等活动对多花黄精的需求越来越大,使野生多花黄精资源大幅度缩减,导致其种群分布面临濒危状态。因此,提出并制定相应合理的策略来保护野生多花黄精种质资源是十分必要的,具体的保护策略如下:第一,加强对现有野生多花黄精种质资源的保护力度,根据本文的研究结果,建议将浙江丽水和江西新余遗传多样性丰富的种群,设定为重点保护和人工育种单位;第二,加强对野生多花黄精生长环境的监管,对有大面积天然种群分布的林分进行保护,严禁破坏其林下生长环境,为其种群繁衍提供有利条件;第三,在对天然分布种群保护的同时,加大对多花黄精培育的研究,可以选择优良种源进行有性和无性育苗;第四,收集全分布区野生多花黄精种质资源,建立野生多花黄精全分布区种质资源保存圃。
猜你喜欢
——如意、半如意、将军帽、金螺、金夏
中国果业信息(2024年1期)2024-04-02 03:52:16
中国防痨杂志(2022年6期)2022-11-25 00:36:00
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02 01:48:06
Digital Chinese Medicine(2020年3期)2020-11-03 03:12:12
云南农业科技(2020年5期)2020-10-14 07:24:50
新疆农业科学(2020年1期)2020-02-14 03:22:46
山东农业科学(2018年1期)2018-02-03 16:59:39
江苏农业科学(2017年7期)2017-05-23 23:49:32
青苹果·教育研究版(2016年7期)2016-12-12 05:19:52
中国麻业科学(2015年5期)2015-12-28 06:22:11
