
苹果树腐烂病菌对苯醚甲环唑的敏感性
2019-06-11 11:55刘娟冯浩王帅高小宁黄丽丽
植物保护 2019年1期
关键词:抗药性
刘娟 冯浩 王帅 高小宁 黄丽丽
0)
摘要為了明确苹果树腐烂病菌对苯醚甲环唑的敏感性现状,本研究采用菌丝生长速率法测定了来自7个省份7个不同年份的106株苹果树腐烂病菌对苯醚甲环唑的敏感性,并建立了敏感基线。结果表明:供试苹果树腐烂病菌菌株的EC50区间在0.003~0.123 μg/mL,敏感性频率分布呈连续性单峰曲线。经KS法检验符合正态分布,未出现敏感性明显下降的抗性群体,平均EC50 (0.044±0.029) μg/mL可作为苹果树腐烂病菌对苯醚甲环唑的敏感基线。不同地理来源的菌株对苯醚甲环唑的敏感性有显著性差异,来自辽宁省的菌株敏感性最高,河南省的敏感性最低;不同年份的菌株敏感性也有显著性差异,2009年采集的菌株对苯醚甲环唑的敏感性最高,2016年采集的菌株敏感性最低,菌株对药剂的敏感性随年份的推移逐渐降低,说明病菌的抗药性不断增强。
关键词苹果黑腐皮壳菌;菌落直径;苯醚甲环唑;EC50;抗药性
中图分类号:S 436.611.11
文献标识码:A
DOI:10.16688/j.zwbh.2018119
苹果树腐烂病(apple Valsa canker)是由苹果黑腐皮壳属真菌Valsa mali侵染引起的枝干病害[1]。该病的发生会造成树皮组织腐烂,树势削弱,甚至枝枯树死,果树寿命缩短[2]。因其侵染途径多样、潜伏性强、致病机理复杂,病害防控困难,治愈率低,被称为“苹果的癌症”[3]。
目前,生产上苹果树腐烂病主要依靠刮除病疤后涂抹药剂或全树喷淋药剂等措施进行防治,所用药剂种类繁多[45]。化学药剂因其操作简单,使用方便且见效快而成为防治苹果树腐烂病的主要药剂。其中甲基硫菌灵可以高效预防腐烂病菌的侵染[6],丁香菌酯能有效抑制苹果树腐烂病病疤的复发[7],戊唑醇能有效抑制苹果树腐烂病菌孢子萌发和菌丝生长[8]。而三唑类杀菌剂苯醚甲环唑虽然尚未在苹果树腐烂病上登记使用,但是其对腐烂病菌室内效果要强于同类杀菌剂戊唑醇[8],可以作为防治苹果树腐烂病的替代药剂。
苯醚甲环唑由先正达公司于1989年研发上市,具有高效、广谱、低毒的特点,对子囊菌门和担子菌门的病菌有很好的防治效果[9]。其作用机制是抑制真菌细胞膜上的麦角甾醇合成,破坏细胞膜结构,造成功能紊乱,从而达到杀菌的目的[1011]。由于该药剂作用位点单一,长期反复使用会引起病菌产生抗药性[12],被认为是一种具有中等抗性风险的杀菌剂。苹果黑星病菌Venturia inaequalis、小麦赤霉病菌Fusarium graminearum对该药剂已经产生了抗性[1314]。近年来,我国各地苹果产业区普遍应用苯醚甲环唑等三唑类杀菌剂防治各类真菌病害,是否有抗药性菌株尚不明确。因此,本研究通过测定采自全国的106株苹果树腐烂病菌对苯醚甲环唑的敏感性,并比较不同地理来源、不同年份的菌株敏感性差异,以期明确苹果树腐烂病菌对该药剂的敏感性现状,为病害的科学防治和病菌的抗药性监测提供重要科学依据。
1材料与方法
1.1材料
1.1.1供试菌株
本实验室自2007至2016年从陕西、山西和河南等省苹果产区的田间发病植株采集分离了106株苹果树腐烂病菌,其中陕西省54株,山西省18株,新疆维吾尔自治区10株,河南省11株,辽宁省5株,山东省4株,四川省4株,用于敏感性测定。
1.1.2供试药剂与浓度
95.6%苯醚甲环唑原药,先正达公司提供;浓度设置为0.006 3、0.013、0.025、0.050、0.100和0.200 μg/mL。
1.2方法
1.2.1含药培养基的制备
将苯醚甲环唑原药溶于丙酮制成10 000 μg/mL母液,加入终浓度为0.2%的Tween80,置于4℃冰箱中备用。用蒸馏水稀释母液,按设置浓度加入到PDA培养基中,充分混匀,配成不同质量浓度的苯醚甲环唑含药平板。
1.2.2敏感性测定
参考Yin等[15]的方法进行。被测菌株于25℃条件下在PDA平板上培养72 h,利用打孔器自菌落边缘打取大小相同、直径为5 mm的菌饼,菌丝面朝下接种于含药平板的中心。每个浓度重复3次,以不加药的PDA平板作为对照。25℃下培养72 h,用十字交叉法测量菌落直径,取平均值,计算不同浓度的苯醚甲环唑对菌丝的生长抑制率。
菌丝生长抑制率=[(对照菌落直径-处理菌落直径)/(对照菌落直径-菌饼直径)]×100%。
以X(药剂质量浓度的对数值)和Y(抑制率对应的几率值)进行回归分析得到毒力回归方程Y=a+bX以及相关系数r,并计算EC50。
1.2.3数据处理
试验数据采用Microsoft Excel 2010和SPSS 16.0进行处理。对EC50进行KolmogorovSmirnov正态性检验,参照祁之秋等的方法[16]获得苹果树腐烂病菌群体对苯醚甲环唑的敏感性频率分布图,建立对苯醚甲环唑的敏感基线。采用Duncan氏新复极差法对不同地区、不同年份的平均EC50进行差异显著性分析。
2结果与分析
2.1苹果树腐烂病菌对苯醚甲环唑的敏感基线的建立
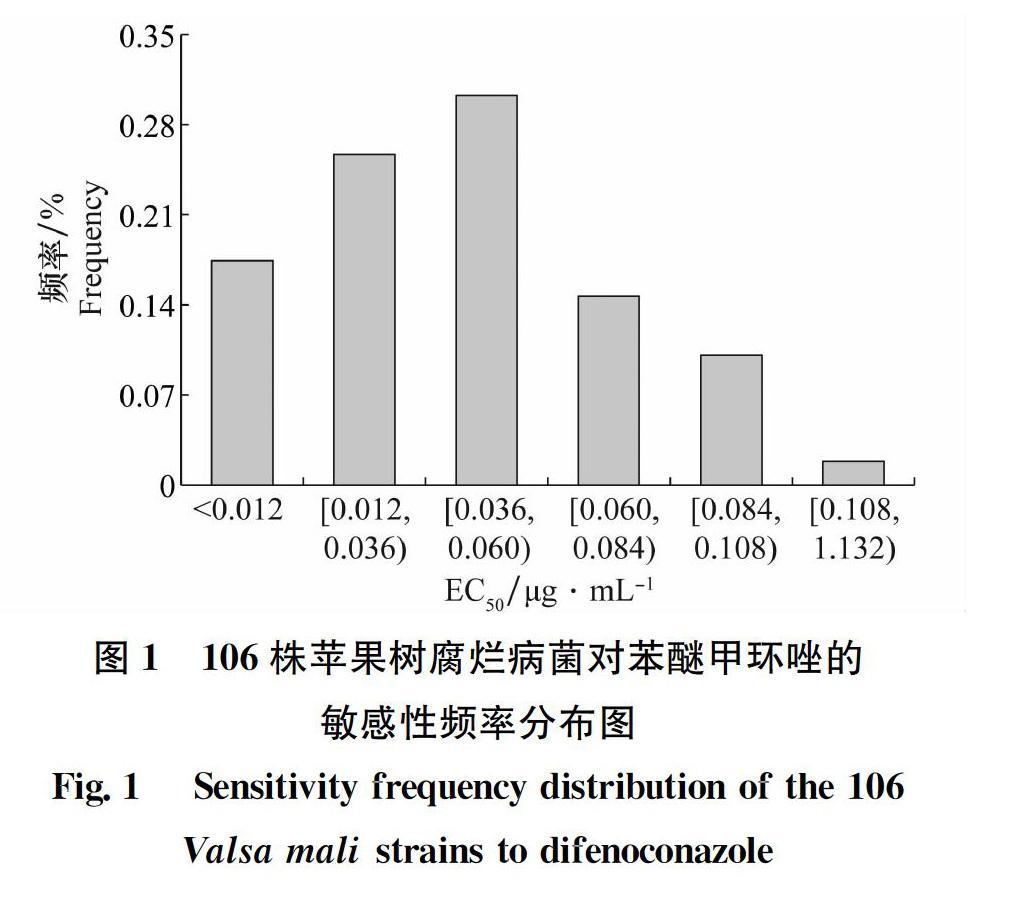
检测发现,苯醚甲环唑对来自7个省的106株苹果树腐烂菌的EC50分布在0.003~0.123 μg/mL,平均值为(0.044±0.029)μg/mL。所检测的菌株对该药剂具有较高的敏感性,未发现抗药性亚群体。但最大值为最小值的41倍,EC50跨度范围比较大,表明群体内个体间对苯醚甲环唑的敏感性差异较大。
将本研究得到的EC50分成6个区间,每个区间出现的菌株频率如图1所示,EC50敏感性频率呈现连续性单峰曲线分布,通过SPSS中的KS正态检验得到P=0.135>0.05,说明菌株对苯醚甲环唑的敏感性频率符合正态分布,按照野生病原群体对药剂的敏感性呈正态分布的原理,本研究中106株病菌的平均EC50(0.044 μg/mL±0.029 μg/mL)可作为苹果树腐烂病菌对苯醚甲环唑的敏感基线。
2.2不同地理来源的菌株對苯醚甲环唑的敏感性水平
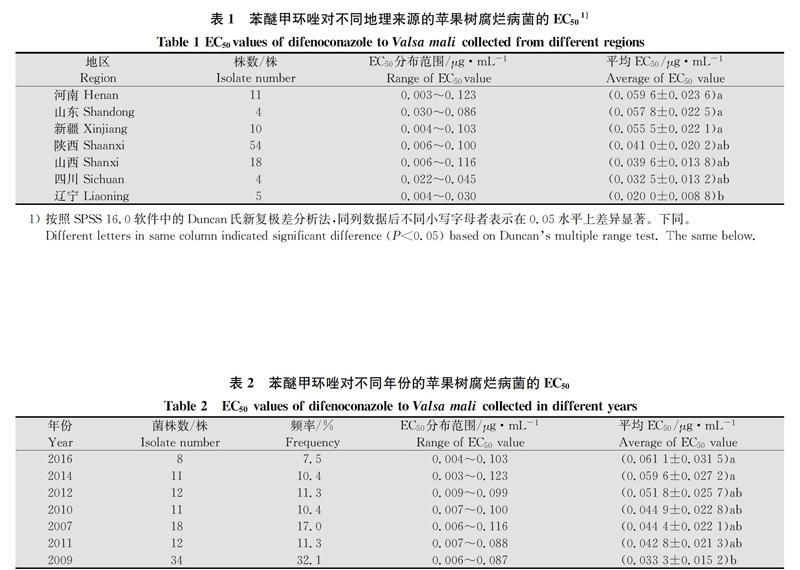
不同地理来源的苹果树腐烂病菌菌株对苯醚甲环唑的敏感性存在一定的差异(表1),最敏感的是辽宁菌株,其EC50平均值为(0.020 0±0.008 8)μg/mL,最不敏感的是河南菌株,其EC50平均值为(0.059 6±0.023 6)μg/mL。山西、新疆、河南的菌株群体内部个体对苯醚甲环唑的敏感性差异较大。辽宁地区的菌株同河南、山东和新疆3个地区的菌株在P=0.05水平上差异显著,其他地区的菌株对苯醚甲环唑的敏感性无显著差异。
2.3不同年份的苹果树腐烂病菌对苯醚甲环唑的敏感性水平
如表2所示,采自不同年份的苹果树腐烂病菌对苯醚甲环唑的敏感性存在一定差异。2009年采集的菌株对苯醚甲环唑的敏感性最高,2016年菌株的敏感性最低,随着年份推移病菌对苯醚甲环唑的敏感性逐渐减弱。
3讨论
杀菌剂用于防治某种植物病害之前,建立其敏感基线有助于优化该杀菌剂的使用策略[17]。虽然苯醚甲环唑已经在生产上应用多年,但苹果树腐烂病菌对其敏感性水平尚不明确。本研究共检测了采自中国7个省份的106株苹果树腐烂病菌对苯醚甲环唑的敏感性。研究结果表明中国苹果树腐烂病菌对苯醚甲环唑还处于野生敏感种群阶段,可将苯醚甲环唑对供试菌株的EC50平均值(0.044±0.029)μg/mL作为中国苹果树腐烂病菌对苯醚甲环唑的敏感基线。该敏感基线可用于监测田间苹果树腐烂病菌群体对苯醚甲环唑敏感性的变化,为评价其抗药性风险和未来制定抗药性治理策略提供参考。
本研究发现苹果树腐烂病菌对苯醚甲环唑的敏感性差异与地理位置有关,这可能与不同地区的苹果种植年限和面积、气候环境条件、腐烂病的发生历史,用药措施以及病菌本身的遗传差异及群体组成等因素有关[18]。此外,随着时间的推移,苹果树腐烂病菌对苯醚甲环唑的敏感性逐渐减弱,可能是由于生产中连续大量使用同类杀菌剂或者其他有交互抗性的杀菌剂,使得病菌对杀菌剂的敏感性发生了改变。为了延缓苹果树腐烂病菌对苯醚甲环唑抗药性的产生,建议在田间通过轮换使用不同作用机制的药剂,使苹果树腐烂病菌对苯醚甲环唑的敏感性处于低抗性频率阶段,以延长农药的使用寿命。
参考文献
[1]WANG Xuli, ZANG Rui, YIN Zhiyuan, et al. Delimiting cryptic pathogen species causing apple Valsa canker with multilocus data [J].Ecology and Evolution,2014,4(8):13691380.
[2]黄丽丽,张管曲,康振生,等.果树病害图鉴[M].西安:西安地图出版社,2001:1.
[3]曹克强,国立耘,李保华,等.中国苹果树腐烂病发生和防治情况调查[J].植物保护,2009,35(2):114117.
[4]焦浩,范艳云,高小宁,等.8种药剂对苹果树腐烂病的田间防效评价[J].河南农业科学,2015,44(10):9599.
[5]翟慧者,胡同乐,陈曲,等.10种化学杀菌剂对苹果树腐烂病的防效评价[J].植物保护,2012,38(3):151154.
[6]TAMURA O, SAITO I. Histopathological changes of apple bark infected by Valsa ceratosperma (Tode ex Fr.) Maire during dormant and growing periods [J]. Annals of the Phytopathological Society of Japan, 2009, 48: 490498.
[7]陈亮,刘君丽,司乃国,等.丁香菌酯对苹果树腐烂病的防治[J].农药,2009,48(6):402404.
[8]王磊,郜佐鹏,黄丽丽,等.防治苹果树腐烂病杀菌剂的室内筛选[J].植物病理学报,2009,39(5):549554.
[9]刘长令.世界农药大全[M].北京:化学工业出版社,2005:122126,155158,281282.
[10]祁之秋,周明国.戊唑醇对小麦纹枯病菌的抑菌作用[J].农药学学报,2003,5(3):8084.
[11]吴学宏,肖建华,张文华,等.2%三唑醇·戊唑醇种衣剂对小麦纹枯病菌麦角甾醇含量的影响[J].农药学学报,2005,7(4):372375.
[12]刘保友,张伟,栾炳辉,等.苹果轮纹病菌对苯醚甲环唑和氟硅唑的敏感性及其交互抗性[J].植物病理学报,2013,43(5):541548.
[13]HENRQUEZ S J L, SAEMIENTO V O, ALACN C P. Sensitivity of Venturia inaequalis Chilean isolates to difenoconazole, fenarimol, mancozeb and pyrimethanil [J]. Chilean Journal of Agricultural Research, 2011, 71(1): 3944.
[14]RCKANOVIE, MIHAJLOVIM, POTONIK I. In vitro sensitivity of Fusarium graminearum (Schwabe) to difenoconazole, prothioconazole and thiophanatemethyl[J]. Pesticide Phytomedicine, 2010, 25(4): 325333.
[15]YIN Y, LIU X, LI B, et al. Characterization of sterol demethylation inhibitorresistant isolates of Fusarium asiaticum and F. graminearum collected from wheat in China [J]. Phytopathology, 2009, 99(5): 487497.
[16]祁之秋,鞠雪娇,纪明山,等.辽宁省稻瘟病菌对咪鲜胺敏感基线的建立[J].农药学学报,2012,14(6):673676.
[17]CHEN Yu, YAO Jian, WANG Wenxiang, et al. Effect of epoxiconazole on rice blast and rice grain yield in China [J]. European Journal of Plant Pathology, 2013, 135(4): 675682.
[18]齐永志,李海燕,苏媛,等.小麦纹枯病菌对噻呋酰胺的敏感性及抗药性突变体的主要生物学性状[J].农药学学报,2014(3):271280.
(责任编辑:杨明丽)
猜你喜欢
农民致富之友(2019年14期)2019-05-24
江苏农业科学(2017年7期)2017-05-23
杂草学报(2016年2期)2017-04-14
农业与技术(2017年4期)2017-04-05
现代农业科技(2017年1期)2017-03-06
江苏农业科学(2016年6期)2016-07-25
江苏农业科学(2016年3期)2016-05-03
江苏农业科学(2016年3期)2016-05-03
江苏农业科学(2015年11期)2016-01-27
长江蔬菜·学术版(2014年5期)2014-07-28
