
低氧耐力运动对营养性肥胖大鼠骨骼肌自噬蛋白表达的影响
2019-06-11 11:11:30赵芳芳文根吴菊花李志刚翁锡全林文弢
中国运动医学杂志 2019年4期
赵芳芳 文根 吴菊花 李志刚 翁锡全 林文弢
1 广州体育学院省重点生化实验室(广州510500)
2 吉林大学珠海学院
3 广西科技大学体育学院
4 百色学院体育学院
自噬(Autophagy)作为真核生物体内存在的自我保护机制,在机体能量代谢中发挥着重要的作用。研究表明,长期高脂饮食会抑制大鼠体内自噬相关因子的活性[1],而运动可激活机体的细胞自噬,并使细胞自噬相关因子的表达出现不同程度的提高[2,3]。除运动外,低氧亦可诱导自噬的发生,薄海等[4]研究发现,在低氧状态下进行中等负荷有氧运动训练,可提高骨骼肌线粒体ATP 输出,抑制活性氧(reactive oxygen species,ROS)生成,线粒体膜蛋白PTEN 诱导激酶1(PTEN induced putative kinase 1,PINK1)、Parkin和Bcl-2/腺病毒E1B 19 k Da 结合蛋白(Bcl2/adenovirus E1B 19 kDa interacting protein 3,BNIP3)表达量显著升高,提示低氧可显著促进线粒体自噬增加。同样,亦有研究表明,营养性肥胖大鼠在低氧条件下运动,其腓肠肌自噬相关因子P62、LC3、Beclin1的表达具有显著性提高[5]。而运动或低氧作为防治肥胖的有效干预手段是如何调控细胞自噬,发挥防控代谢性疾病的作用,值得进一步探讨。因此,本研究通过建立营养性肥胖大鼠模型,并对其进行不同浓度的低氧或结合耐力运动干预,探讨在持续性高脂饮食的条件下,低氧、运动、低氧结合运动对骨骼肌细胞自噬相关蛋白微管相关蛋白1 轻链3(Microtubule-associated protein 1 light chain 3,LC3)、酵母ATG6 同源物(Beclin1)、AMPKα2、Sestrin2 表达的影响,旨在为代谢性疾病的防治提供理论依据和实验支撑。
1 材料与方法
1.1 低氧耐力训练模型
选取建模成功的营养性肥胖大鼠80只,按不同浓度的低氧分为安静组和运动组(具体分组情况如表1),每组各10只,进行1周(第1~6天)常氧跑台适应性训练和1 周(第8~13 天)低氧结合耐力运动适应性训练(大鼠适应期训练方案见表2)。正式试验干预期,大鼠训练方案如下:运动组大鼠均进行中等强度耐力运动,其中时间为40 min,跑台坡度为0,跑速为20 m/min;运动频率为一周5次(每周一到周五),持续8周。其中,A组大鼠不进行任何干预,AE组大鼠进行常氧中等强度耐力训练,B 组、C 组、D 组分别给予氧浓度为16.3%、13.3%、11.3%的低氧暴露(在相应的低氧环境下,每天进行连续12 h 低氧干预,干预时间为早8:00 到晚8点),其中BE组和CE 组、DE组大鼠分别给予氧浓度为16.3%、13.3%、11.3%的低氧暴露并增加中等强度耐力运动训练(低氧暴露时间与低氧组相同,但在晚7点进行40 min的低氧耐力训练)。

表1 低氧运动训练大鼠分组情况(各组n=10)

表2 适应期运动方案
1.2 动物取材
末次低氧耐力训练结束后,禁食24 小时,称量大鼠体重并测体长,腹腔注射10%水合氯醛溶液(按0.3 ml/100 g体重剂量)进行大鼠麻醉,腹部解剖,取腓肠肌组织称重,并取一侧腓肠肌冰上匀浆抽提总蛋白,另一侧腓肠肌置冻存管于液氮中速冻,随后于-70℃超低温冰箱保存,待测。
1.3 Western blot法测定骨骼肌自噬相关蛋白
骨骼肌总蛋白提取:电子天平称取100 mg 腓肠肌并剪碎,放入1 ml RIPA 蛋白裂解液(碧云天)和10 μl PMSF 蛋白酶抑制剂(康为世纪)的混合液,于冰上研磨并裂解20 min;之后,低温离心机以-4℃、12000 rpm离心15 min,上清液于1.5 mL的EP管存放。BCA 法测定蛋白浓度后与上样缓冲液于沸水中煮5 min,使蛋白变性。15%、10%分离胶,5%浓缩胶电泳;采用湿转法将蛋白转移至PVDF 膜;5%脱脂牛奶封闭过夜;一抗(LC3、Beclin1、AMPKα2、Sestrin2 均购置美国ABCAM公司)、GAPDH(美国Proteintech Group公司)均用TBST缓冲液以1∶1000和1:8000稀释,于4℃冰箱孵育过夜;二抗(Sera care 公司)以1∶5000 稀释,室温摇床孵育2 h。Western blot 结果使用Tanon(天能)5200 Multi 仪器自带软件进行灰度值分析,以目的蛋白/内参值作为目的蛋白的半定量分析。
1.4 数据统计
采用SPSS22.0 统计软件进行统计学处理,实验所得数据均以平均值±标准(x±s)表示,GraphPad Prism5进行数理统计及图像生成,组间比较采用单因素方差分析(One-way ANOVA),以P<0.05为差异有统计学意义。
2 结果
2.1 各组大鼠腓肠肌细胞Sestrin2蛋白的表达情况
由图1可知,实施低氧暴露干预后,与A组相比,C组、D 组Sestrin2 蛋白表达水平显著性升高(P<0.05),B组有上升趋势;与B组相比,D组Sestrin2蛋白表达水平显著性升高(P<0.05)。实施耐力运动干预后,与安静组大鼠相比,运动组大鼠腓肠肌Sestrin2蛋白表达均具有升高趋势,其中AE 组与A 组、BE 组与B 组、DE 组与D组相比Sestrin2蛋白表达水平升高,差异有统计学意义(P<0.05)。低氧结合耐力运动双重干预后,与AE组相比,BE组、CE组Sestrin2蛋白表达有上升趋势,DE组Sestrin2蛋白表达显著性升高(P<0.05);与BE组、CE组相比,DE组Sestrin2蛋白表达显著性升高(P<0.05)。
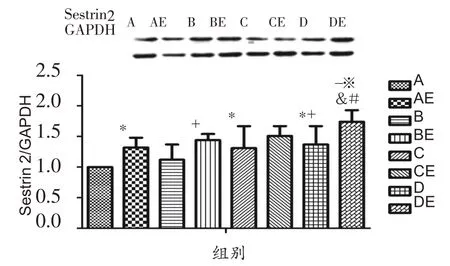
图1 低氧或结合耐力运动对肥胖大鼠腓肠肌细胞Sestrin2蛋白表达的影响
2.2 各组大鼠腓肠肌细胞LC3Ⅱ/LC3Ⅰ比值的变化情况
由图2可知,实施低氧暴露干预后,与A组相比,C组、D组LC3Ⅱ/LC3Ⅰ比值显著性升高(P<0.05),B组具有上升趋势;与B 组相比,D 组LC3Ⅱ/LC3Ⅰ比值显著性升高(P<0.05)。实施耐力运动干预后,与各安静组大鼠相比,运动组大鼠腓肠肌LC3Ⅱ/LC3Ⅰ比值均显著性升高(P<0.05)。低氧结合耐力运动双重干预后,与AE组相比,BE组LC3Ⅱ/LC3Ⅰ比值有上升趋势,CE组、DE组LC3Ⅱ/LC3Ⅰ比值显著性升高(P<0.05);与BE组相比,DE组LC3Ⅱ/LC3Ⅰ比值显著性升高(P<0.05)。
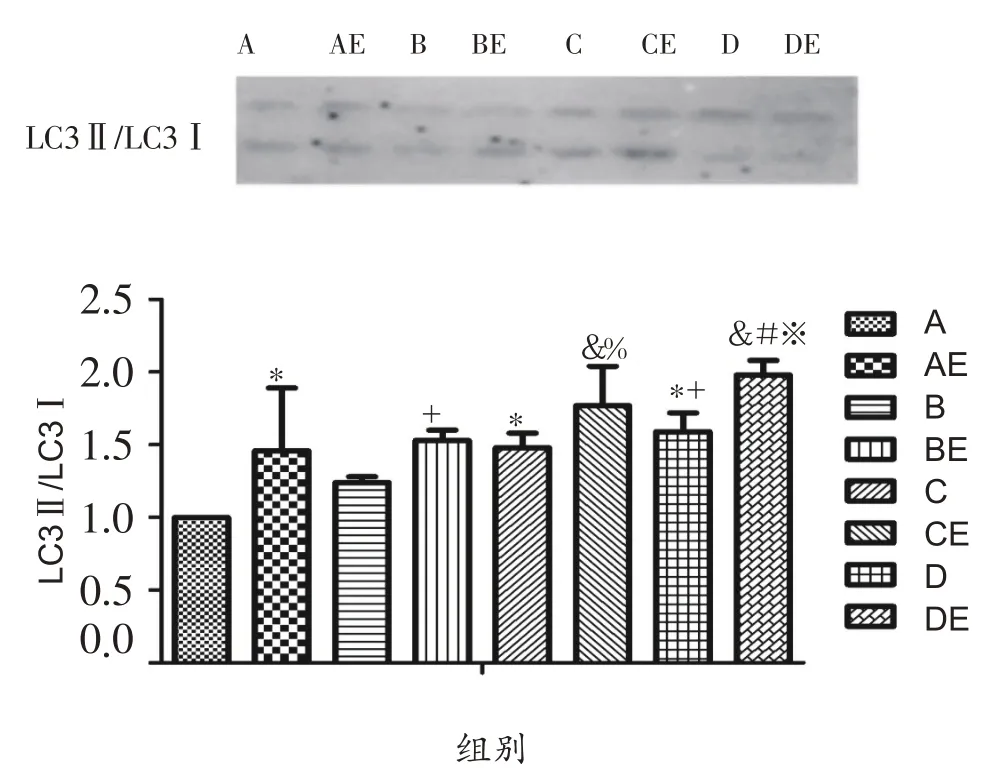
图2 低氧或结合耐力运动对肥胖大鼠腓肠肌细胞LC3Ⅱ/LC3Ⅰ比值的影响
2.3 各组大鼠腓肠肌细胞Beclin1蛋白表达情况
由图3可知,实施低氧暴露干预后,与A组相比,B组、C组、D组Beclin1蛋白表达显著升高(P<0.05),与B组、C 组相比,D 组Beclin1 蛋白表达显著升高(P<0.05)。实施耐力运动干预后,与安静组大鼠相比,运动组大鼠腓肠肌Beclin1 蛋白表达均显著升高(P<0.05)。低氧结合耐力运动双重干预后,与AE组相比,BE组、CE组Beclin1蛋白表达具有上升趋势,DE组Beclin1 蛋白表达显著升高(P<0.05);与BE、CE 组相比,DE组Beclin1蛋白表达显著升高(P<0.05)。
2.4 各组大鼠腓肠肌细胞AMPKα2蛋白表达情况
由图4可知,实施低氧暴露干预后,与A组相比,C组、D 组AMPKα2 蛋白表达显著性升高(P<0.05);与B组相比,C 组、D 组AMPKα2 蛋白表达显著升高(P<0.05)。实施耐力运动干预后,与安静组大鼠相比,运动组大鼠腓肠肌AMPKα2 蛋白表达均显著升高(P<0.05)。低氧结合耐力运动双重干预后,与AE组相比,BE 组AMPKα2 蛋白表达具有上升趋势,CE 组、DE 组AMPKα2 蛋白表达显著升高(P<0.05),与BE 组、CE 组相比,DE组AMPKα2蛋白表达显著性升高(P<0.05)。
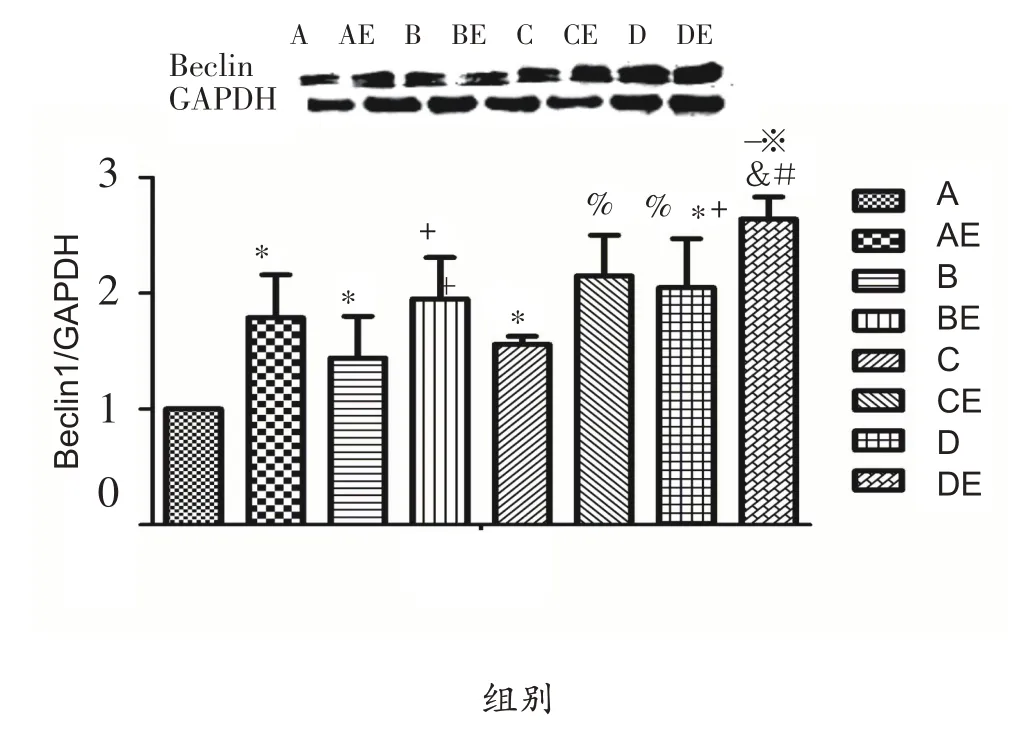
图3 低氧或结合耐力运动对肥胖大鼠腓肠肌细胞Beclin1蛋白表达的影响
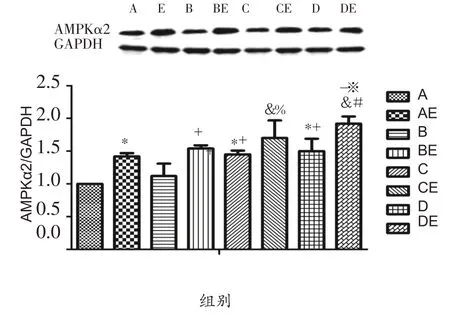
图4 低氧或结合耐力运动对肥胖大鼠腓肠肌细胞AMPKα2蛋白表达的影响
3 分析与讨论
3.1 低氧或结合耐力运动对肥胖大鼠腓肠肌细胞LC3Ⅱ/LC3Ⅰ比值的影响
LC3Ⅱ被认为是自噬活动的特异性标志物,通常用LC3Ⅱ/LC3Ⅰ比值来衡量自噬活性,其比值下降提示自噬活性减弱,反之则说明自噬活性增强[6,7]。崔迪等[8]研究发现,肥胖小鼠进行跑台耐力训练6周后,高脂对照组LC3Ⅱ/LC3Ⅰ比值下降,普通膳食组、高脂运动组小鼠骨骼肌p62蛋白水平下降,高脂运动组LC3Ⅱ/LC3Ⅰ比值上调。本实验结果与之一致,经过8 周的低氧或结合耐力运动后,与常氧安静组相比,常氧运动组、低氧组大鼠腓肠肌LC3Ⅱ/LC3Ⅰ比值具有升高趋势,并且常氧运动组、13.3%低氧安静组、11.3%低氧安静组大鼠骨骼肌LC3Ⅱ/LC3Ⅰ比值显著性升高,说明耐力运动及低氧暴露可激活自噬。与常氧运动组相比,13.3%低氧运动组、11.3%低氧运动组大鼠腓肠肌LC3Ⅱ/LC3Ⅰ比值显著性升高,说明在一定范围内,氧浓度越低,自噬激活的效果越明显。此外,就整体而言,11.3%低氧组激活自噬效果最明显,尤以11.3%低氧耐力运动组效果显著。
3.2 低氧或结合耐力运动对肥胖大鼠腓肠肌细胞Sestrin2、AMPKa2蛋白表达的影响
Sestrin 蛋白包含三个不同的亚基,并根据特定蛋白编码的基因分SESN1,SESN2 和SESN3,并且这三种亚基包含近50%的相同氨基酸序列[9,11]。作为应激诱导型代谢调节器,Sestrins 通过多种机制帮助细胞适应各种应激刺激,包括分解代谢反应的激活、合成代谢活动的停止以及启动细胞修复机制以维持细胞内稳态[9]。Sestrins(SESN)是具有多效生物功能的高度保守的蛋白质,并且在应激条件下上调,如DNA 损伤、缺氧、饥饿、生长因子耗尽、辐射和氧化应激等[10,11]。诱导后,Sestrins 保护细胞对抗基因毒性和氧化应激,因此被命名为应激诱导型代谢调节剂[9]。Sestrin 基因的失活会导致各种细胞和代谢疾病,如氧化损伤,线粒体功能障碍,脂肪堆积,肌肉退化,加速糖尿病并发症进展[12-14]。已知Sestrin2(SESN2)是一种高度保守的应激诱导型代谢蛋白,可抑制活性氧(ROS),并为各种有害刺激(遗传毒性和氧化应激、内质网应激和缺氧等)提供细胞保护。Lee等[15]研究发现,敲除Sestrin2/3基因的小鼠在正常的饮食条件下可出现胰岛素抵抗症状,这提示Sestrins 在机体代谢调控中可能发挥着重要作用。此外,当采用si RNAs 抑制细胞SESN2 的基因表达后发现细胞自噬活性出现显著性降低[16],这提示应激状态下细胞自噬活性的增强可能是Sestrins 发挥细胞保护作用的重要机制之一。研究发现,8 周的有氧运动可上调Sestrin2 和Sestrin3 的表达水平,并且单次急性运动可使AMPKα2 与Sestrin2 和Sestrin3 之 间 结 合 显著 增加[17]。本研究结果表明,运动、低氧均能上调Sestrin2蛋白的表达水平,经过8周低氧或结合耐力运动,运动组、低氧组、运动结合低氧组肥胖大鼠腓肠肌Sestrin2蛋白都有不同程度的提高,尤以11.3%低氧运动组大鼠腓肠肌的Sestrin2蛋白表达上调尤为显著,这可能与缺氧状态下HIF-1ɑ 和NO 的活性有关,但其具体机制仍需进一步研究。
Sestrin2 还通过激活关键能量感受器AMP 依赖性蛋白激酶(AMPK)和抑制哺乳动物雷帕霉素复合物1(mTORC1),在代谢调节中起关键作用。AMPK是一种在能量缺乏条件下激活的酶,可作为细胞中mTOR 的主要负调节因子。研究表明,Sestrin2 主要通过激活AMPK 和结节性硬化症2(TSC2)的磷酸化来抑制细胞中mTOR的活化[12,18]。在小鼠中,Sestrin2的基因切除增强了mTOR的激活,加重了肥胖相关特征,如葡萄糖耐受不良,胰岛素抵抗和肝脏脂肪变性等[12]。本研究结果表明,经过8周的低氧耐力运动,低氧或结合耐力运动均能够上调AMPKα2蛋白的表达水平,并且,在三种低氧浓度的干预下,11.3%低氧结合耐力运动的效果更为显著。本研究结果发现,运动组大鼠腓肠肌AMPKα 2蛋白表达水平要高于安静组大鼠,低氧结合耐力运动组大鼠腓肠肌AMPKα2蛋白表达水平要高于单纯低氧组。有研究发现,短暂低氧暴露(氧浓度为0.2%O2)即可增加心肌细胞AMPK 活性,且呈持续增加状态[19]。与此相似,Liu 等[20]研究发现慢性低氧暴露可增强机体大脑AMPK 的表达,进而抑制mTOR 信号通路,从而增强低氧暴露所引起的细胞自噬。本实验结果与吴菊花[21]的研究结果一致,由此可推测,低氧或结合耐力运动可正向激活AMPK 蛋白的水平。研究结果显示,氧浓度与运动的叠加效应要优先于单纯性低氧和单纯性运动,亦可推测,在一定范围内,氧浓度越低,对自噬因子AMPK的影响越显著。牛燕媚等[22]研究发现,长期进行有氧运动的小鼠,其骨骼肌中Sestrin2/3的表达水平会有显著性增加,并且其与AMPK 的结合也会显著增加。本研究结果与之相似,实验发现肥胖大鼠腓肠肌的AMPKα2 能够被运动和低氧诱导,并且其表达水平在11.3%低氧运动组尤为显著,Sestrin2蛋白表达趋势与AMPKα2 基本一致。由此,我们可以推测,Sestrins可能参与AMPK 的激活,参与长期低氧或结合有氧运动对骨骼肌自噬活性的激活,并在调节糖、脂代谢方面起到关键作用。虽然在哺乳动物中Sestrins 能够激活AMPK 已得到证实[23],但是Sestrins 是直接激活还是与其他因子结合之后再发挥作用,其具体机制仍不清楚。Maiuri等研究发现,在营养限制或者Rapamycin干预条件下,shRNA介导的Sestrin2表达抑制显著降低了自噬活性水平[24],综合刘效磊[17]和本实验的研究结果,可以推测Sestrins 能够正向调控AMPK 的表达,并在其上游发挥重要作用。
3.3 低氧或结合耐力运动对肥胖大鼠腓肠肌细胞Beclin1蛋白表达的影响
研究表明,低氧应激后细胞内ROS水平升高,激活了核因子κB(NF-κB)信号通路,使Beclin1 活性增高,可见Beclin1活性表现与低氧应激密切相关[25,27]。本研究结果发现,经过8周的低氧或结合耐力运动,与常氧安静组相比,其余各组肥胖大鼠腓肠肌Beclin1蛋白的表达都有上升的趋势。此外,本研究发现,与常氧安静组相比,常氧运动组大鼠腓肠肌Beclin1蛋白表达有显著性升高;与单纯性低氧暴露组相比,各低氧结合耐力运动组大鼠腓肠肌Beclin1 蛋白表达有显著性升高,11.3%低氧组Beclin1蛋白表达高于常氧组、16.3%低氧组及13.3%低氧组。由此,我们可以看出,单纯性耐力运动、低氧暴露、低氧结合耐力运动均能提高肥胖大鼠腓肠肌Beclin1蛋白表达水平。同样,Zhang等[26]报道慢性低氧可上调Bnip3和Beclin1蛋白的表达。由此可以说明,低氧与耐力运动两种刺激均可以提高肥胖大鼠骨骼肌Beclin1的表达水平,使骨骼肌细胞自噬活性增强,从而有利于维持细胞内环境的稳态,调节脂代谢紊乱。此外,低氧结合耐力运动组Beclin1表达水平明显高于单纯性耐力运动组和单纯性低氧暴露组,证明低氧和运动对大鼠骨骼肌自噬基因表达的诱导具有累积效应。
综上,低氧或结合耐力运动可激活细胞自噬,并诱导肥胖大鼠骨骼肌自噬相关蛋白表达增加,且低氧结合耐力的诱导效果要优于单纯性运动及低氧暴露。但自噬作为一种细胞保护机制,其活性过高或过低均不利于胞浆中蛋白质和细胞器生理功能的发挥,所以运动负荷、运动方式以及时间对自噬的调控还需进一步研究。
4 结论
本研究通过对营养性肥胖大鼠进行低氧耐力训练发现,运动和低氧都能显著激活骨骼肌细胞自噬,上调骨骼肌自噬相关蛋白LC3、Beclin1、AMPKα2、Sestrin2蛋白的表达水平。且与单纯运动和单纯低氧相比,低氧耐力训练对自噬的调控作用更加显著。纵观对自噬的研究,有关运动与自噬的研究尚处于起始阶段,低氧结合运动与自噬的研究更是在探索时期,因此,在之后的实验研究中,还需进一步探讨运动、低氧激发自噬的机制以及自噬适应性变化的原因。
猜你喜欢
分子诊断与治疗杂志(2023年8期)2023-09-07 12:00:34
天津医科大学学报(2021年3期)2021-07-21 09:03:42
中成药(2018年10期)2018-10-26 03:41:40
国外医药(抗生素分册)(2016年5期)2016-07-12 14:25:34
中国运动医学杂志(2016年3期)2016-07-10 12:07:23
中国运动医学杂志(2016年3期)2016-07-10 12:07:23
磁共振成像(2015年5期)2015-12-23 08:52:53
医学研究杂志(2015年5期)2015-06-10 06:43:26
实验动物与比较医学(2014年5期)2014-02-28 14:53:12
中华骨与关节外科杂志(2014年4期)2014-01-22 12:14:24
