
古尔班通古特沙漠中4种荒漠草本植物的生物量与化学计量特征
2019-06-06 02:40:16庄伟伟
植物研究 2019年3期
郭 浩 庄伟伟 李 进
(新疆师范大学生命科学学院,新疆特殊环境物种保护与调控生物学实验室,干旱区植物逆境生物学实验室,乌鲁木齐 830054)
生物量反映了生态系统生产力的大小,是研究生态系统物质循环和能量流动的基础[1]。生物量在植物中的分配策略是植物生活史理论研究的主要内容,反映了植物光合产物在地上和地下各器官中的不同投资分配[2]。就荒漠草本植物而言,在荒漠生态系统物种多样性中占据着主体,也更是陆地生态系统的重要组成部分。相比于森林生态系统,荒漠生态系统具有较低的生物多样性和生态系统稳定性[2~3]。在古尔班通古特荒漠植物区系中,草本植物占重要驱动因子。生物量在叶、枝和根系之间的分配是植物对环境长期适应的结果,并在植物的生长、发育和繁殖等方面有巨大的作用[4]。同时,植物生物量及其分配策略的研究也是生态系统碳循环的基础,是理解生态系统中碳的分配及存储的关键,在全球气候变化的背景下,这有很小的生物量比例,但贡献了绝大部分的物种丰富度和多样性[5~6],并且对荒漠生态系统净初级生产力、碳动态、能量流动及养分循环方面都起着重要作用[7~8]。短命植物和一年生草本植物是荒漠草本植物区系的主要构成。一年生荒漠草本植物是一类在4~8月保持较长生长期的耐旱植物,在维持夏末和初秋时期荒漠生态系统稳定性方面也起一定的作用[9]。生物量的分配主要是受遗传特性、环境条件和个体发育的影响,且分配模式在不同生活型植物间也存在着较大差异[8]。
化学计量学是研究多重化学元素平衡的科学[10]。化学计量学的驱动因素包括生物因素与非生物因素,植物化学计量学特征可能受多种因素协同影响从而改变了从单因素限制角度分析获得的植物化学计量学变化规律[11]。相较于其它常量或微量元素,C、N、P三种元素的耦合作用更强[12],因此目前化学计量学多集中于对C、N、P的研究。相关研究表明,植物N、P元素的含量不仅对植物自身起到调控作用而且随着植物的生长会最终影响生物量的分配[13~14],对古尔班通古特沙漠来说土壤有机质含量较少,降雨量匮乏,植物所需的营养元素也不足以支撑植物整个周期的生长发育。正是由于在不同生长期,植物体内各个元素间的非平衡分配使得植物利用有限的雪水迅速完成生活史[15]。一般情况下,植物随生长周期的变化,细胞内的贮藏性物质比例会逐渐增多[16~17],但是由于不同生长季节,植物体内N、P计量特征也可能会出现明显差异[10]。那么一年生荒漠草本植物在不同的环境与生长压力下生物量分配比例也是否会随着时间和物种的不同而发生改变,以及生物量的分配模式与N、P计量比之间具有如何相关生长关系?
为此,在古尔班通古特沙漠开展了野外原位实验。为使实验样地异质性最小化并达到良好的可操作性,我们选择了分布广泛、小尺度分布较均匀、植株较大的4种一年生草本植物沙蓬(Agriophyllumsqarrosum(L.)Moq.)、雾冰藜(Bassssiadasyphylla(Fisch.et Mey.)O.Kuntze)、角果藜(CeratocarpusarenariusL.)和碱蓬(Suaedaglauca(Bge.)Bge.)对比分析这4种同一生活型植物生物量分配特征与不同生长期化学计量比随植物生长的变化。该研究有助于认识荒漠草本植物生物量分配随化学计量特征的变化模式,及其地上生物量与地下生物量间的异速生长关系。旨在阐明荒漠植被对极端环境的适应机制,并为判断化学计量学是否影响植物生物量分配提供参考。同时对荒漠草本植物的生存策略和功能特征做出进一步了解,并为荒漠植被的恢复与保护提供科学依据。
1 材料与方法
1.1 研究区概况
研究区位于古尔班通古特沙漠(44.16°~46.33°N,84.52°~90.00°E),该沙漠是我国最大的固定和半固定沙漠,年蒸发量大于2000mm,而年降水量普遍不超过150mm,沙漠腹地的年降水量仅有70~100mm,且主要集中于冬春季;空气相对湿度50%~60%,5~8月空气相对湿度通常小于45%[18~19]。沙漠草本植物种类十分丰富,广泛分布于沙漠的短命植物和一年生植物在荒漠生态系统中具有不可替代的生态功能[9],在时空分布上的独特性对荒漠生态系统土地生产力恢复和防止荒漠化均具有重要作用[20~21]。春季仅荒漠一年生草本植物种类就占该沙漠全部植物种类的47%,且一年生短营养期植物的春季绿色产量能占到群落总产量的60%以上[22]。
1.2 数据采集
2018年4月初在古尔班通古特沙漠南部(44.37°N,86.12°E)选择一处(30m×30m)大型样方,将其划分为4块10m×10m的样方。因考虑到沙丘空间异质性较大以及各物种生长期的差异,所以在每个10m×10m的样方内又划分15个2m×2m的小样方,共计60个。该区域土壤类型为风沙土。经前期的野外调查发现,4种一年生草本植物在各个部位均有分布。故对以上4种植物分别在沙丘腹地进行取样。在第二对真叶出现时开始初次取样,取样次数定为4次,每次间隔15天左右,视为各植物的不同生长期。4种长营养期植物的初次取样时间为5月22日,末次取样时间为7月10日。每次调查时,在各个小样方中随机选择3~4株完整的植株,每个物种每次共获取30株。采用全株挖掘法获取样地内供试植物,根系尽可能深挖,以保证其完整。在获取包含根系的土块后用喷壶将植株冲洗干净。随后晾干并将获得的植株装入封口袋带回实验室,进行两个小时120℃杀青后置于70℃条件下烘干至恒重。用0.0001g天平称重并使用球磨仪将地上地下部分分别研磨成粉末。
1.3 N、P元素测定
总生物量(TB)=地上生物量(AGB)+地下生物量(BGB)
(1)
地上生物量(AGB)=茎干物质重+叶干物质重
(2)
地下生物量(BGB)=根干物质重
(3)
根冠比(R/S)=地下生物量/地上生物量
(4)
植物N含量凯氏定氮仪测定;植物P的含量用钼锑抗比色法进行测定。测定之前需将植物粉末放入烘箱进行烘干,防止空气水分与元素的污染。
1.4 数据分析
对4种草本植物不同生长期的地上生物量(AGB)、地下生物量(BGB)、总生物量(TB)、根冠比(R/S)、N、P含量及N∶P首先进行正态性检验,不符合正态分布的数据通过对数转换使之服从正态分布,并利用Levene’s test检验方差齐性。并对以上指标进行单因素方差分析(差异显著性水平为0.05,极显著差异为0.01),在具有显著差异和方差不齐时,用Duncan和T2Tamhane’s test进行多重比较[21]。之后采用变异系数比较整个生长季植物N、P含量及N∶P的变异性(弱变异:0≤CV<0.15;中等变异0.15≤CV<0.35;强变异CV≥0.35),并对生物量与化学计量特征进行Pearson相关性分析[23]。常规数据的整理及分析分别在Excel2016和SPSS22.0软件中完成。
利用相关生长关系模型Y=β·Xα对4种植物地上生物量与地下生物量进行相关生长分析(Y是地上生物量,X是地下生物量,β为标准化常数,α为相关生长指数)。相关生长指数、以及等速生长检验均使用SMATR软件包在R软件中完成[1,3,21]。
2 结果与分析
2.1 生物量及其比例
据表1可知4种一年生荒漠草本植物的地上生物量(AGB)、地下生物量(BGB)和总生物量(TB)随生长期的变化呈持续增长趋势。在5月22日首次取样时,角果藜、沙蓬、碱蓬和雾冰藜的单株TB分别达到0.10、0.31、0.33和1.46g,经过整个生长季的演变在末次取样时分别达到取样初期的4.10、13.78、5.58和2.25倍,沙蓬的生物量积累位于4种荒漠植物之首。此外,从跨越整个生长季的宏观角度来看,4种植物从上一个生长期到当前生长期的生物量累积速率均不相同。
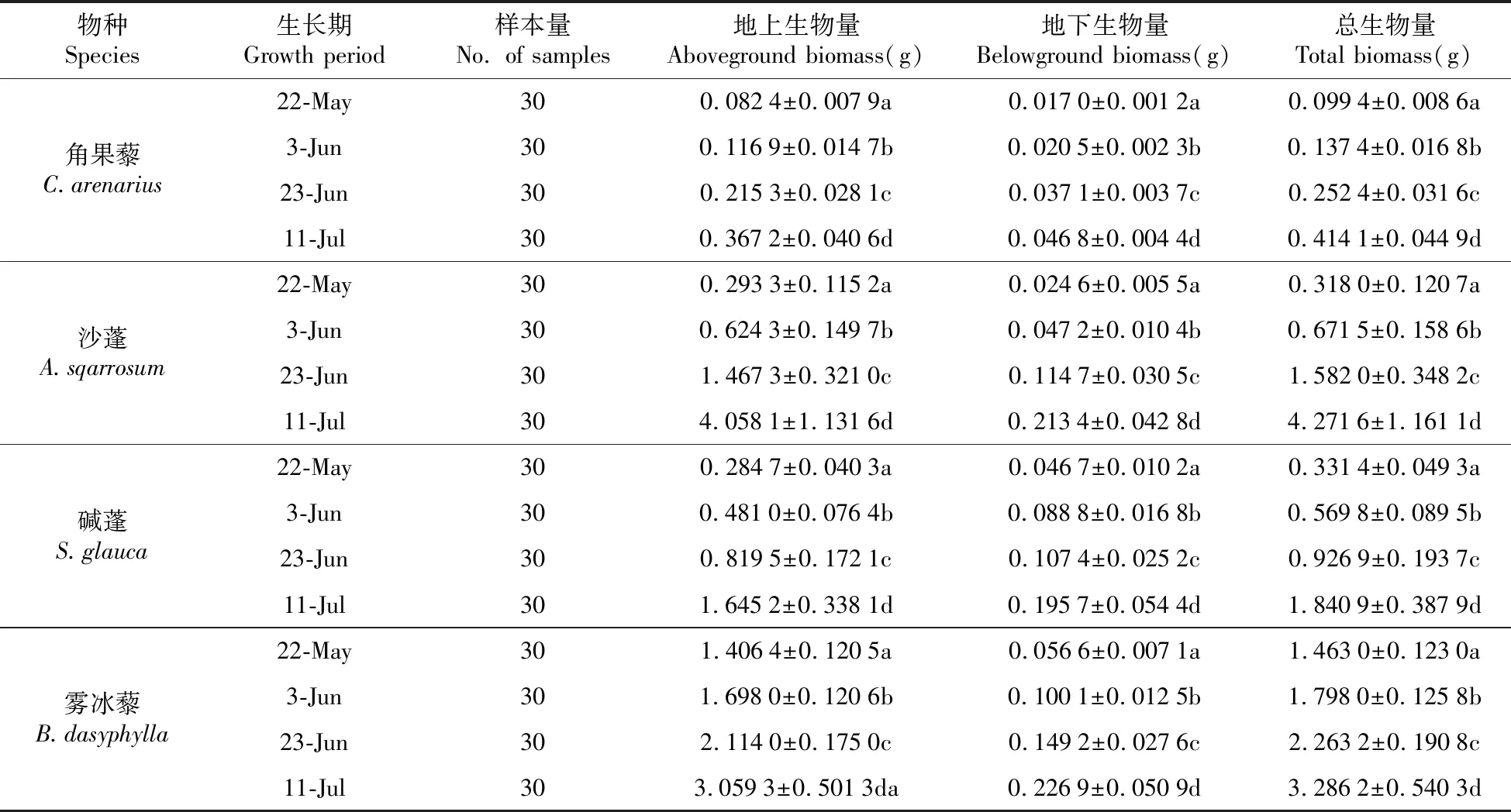
表1 4种草本植物不同生长期的生物量
注:不同字母表示在0.05水平上差异显著 下同。
Note:Different letters in the same column indicate significant differences at the level of 0.05 The same as below.
进一步分析4种同一生活型草本植物的生物量分配和根冠比(R/S)发现与生物量呈显著递增的趋势不同,R/S全部呈现递减的趋势(图1),表现为生长初期最大(0.11~0.74),生长末期最小(均在0.1左右)。说明了在植物的生长的同时将更多的生物量积累到地上部分,以便获取足够多的光照伴随自身的营养分配进而快速完成生活史。植物生长初期由于积雪融化和降雨浇灌呈现出的高R/S,可能是由于植物需要扩延根系以从土壤中获取足够的水分和养分来保证自身存活和生长。随着4种荒漠植物生活史的完成,末期R/S间均无显著差异(P>0.05),说明植物在初期对于获取地下资源的物质分配能力和阈值可能不同,但最终的生物量分配比例趋同。
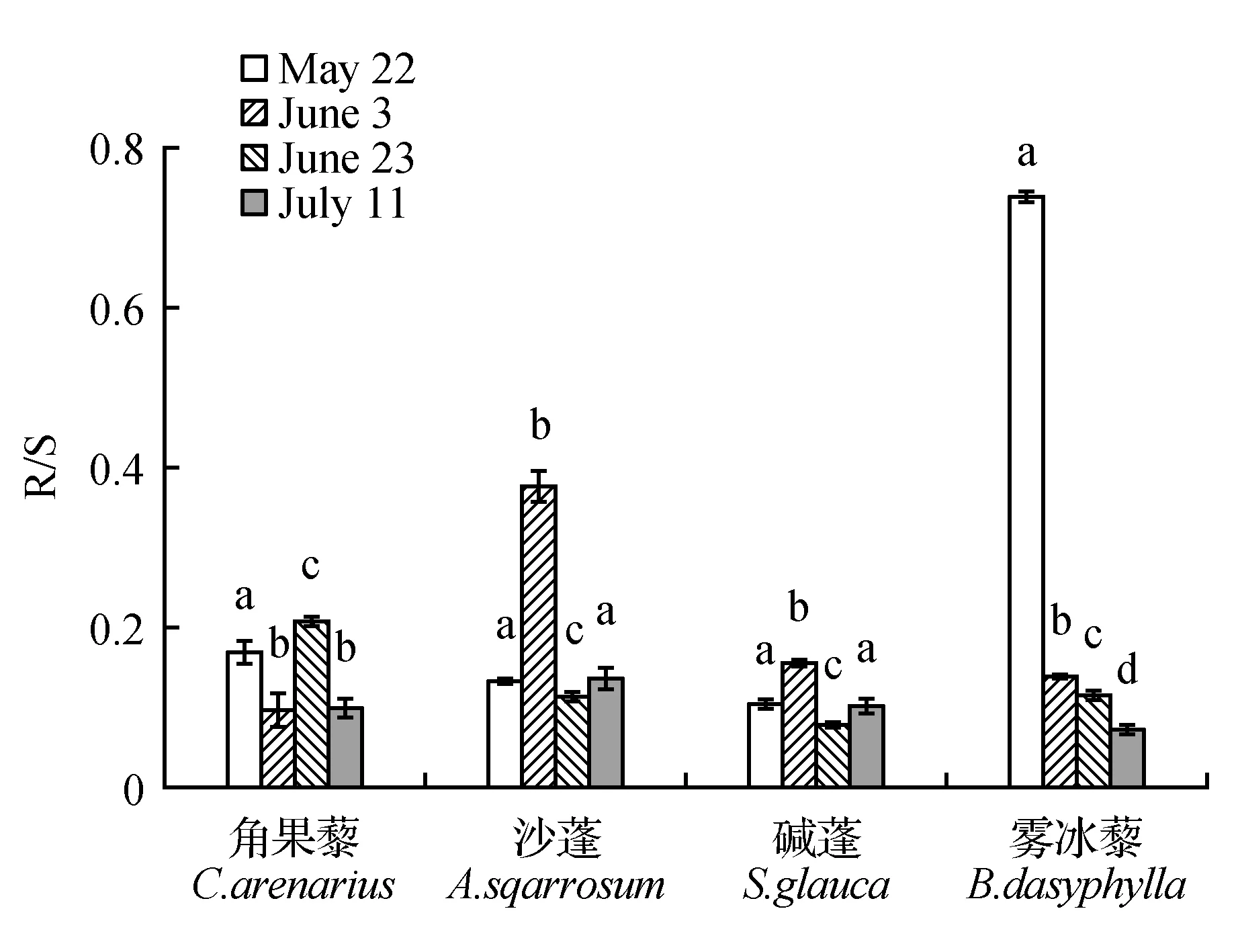
图1 4种荒漠草本植物不同生长期的根冠比(R/S)(平均值±标准误)Fig.1 Root-shoot ratio(R/S) of 4 kinds of desert herbaceous plants in different growth stages(mean±SE)
2.2 AGB、BGB间的相关生长关系
方差分析表明,4种荒漠草本植物地下—地上间仅5月22日的雾冰藜(0.064)无相关生长关系,其中5月22日的角果藜(0.004)、6月3日和6月23日的雾冰藜(0.029、0.005)具有显著(P<0.05)差异,剩余指标仅具有极显著差异(P<0.001)(图2,表2)。相关生长指数显示,整个生长周期相关生长指数动态变化的格局也有所异同,最大的为6月3日的雾冰藜(2.246),最小为5月22日的沙蓬(0.534),当P<0.01时,α≠1。由此可知:角果藜异速生长关系表现为等速→异速的变化过程;沙蓬从采样初期到末期的变化为异速→等速生长;而沙蓬和碱蓬的相关生长指数相反,分别由异速→等速→异速、等速→异速→等速的生长过程。故4种一年生荒漠草本植物不同生长期生物量的相关生长关系在同一生活型之间也存在较大差异。
2.3 4种植物不同生长期叶片化学计量特征的变化
随着植物的生长,4种不同荒漠植物的N、P及其化学计量比在植物体内存在一定的差异。由图2可知碱蓬、角果藜、雾冰藜和沙蓬4种植物N、P含量随生长期的延长逐步降低。在整个生长期中,4种植物的N与P含量的变化趋势均为相似,体现了二者间较强的耦合作用和协同变化的特点。碱蓬、雾冰藜、角果藜和沙蓬在各生长期植物N∶P范围分别为12.84~17.76、12.13~19.73、7.69~13.2和5.68~6.95 mg·g-1,碱蓬、角果藜和沙蓬呈现出先增加后缓慢降低的趋势,而雾冰藜在整个生长期N∶P先增加后降低,在7月10日又展现出缓慢增长的状态;4种植物除雾冰藜C∶P先增加后降低外C∶N、C∶P的变化都表现增长趋势。并随着生长期的延长呈现不同的变化规律(图2)。同时对4种草本植物N、P含量及其计量比的各指标在整个生长季节综合变异分析(表3),可知植物N含量的变异系数最大为30.00%。其次为N∶P的变异(20.12%),比值变化范围为14.90%~25.91%。对于4种荒漠草本植物的N、P及化学计量比在整个生长季所表现出不同的变异性可能是由于同种生活型植物不同生长期对养分吸收和利用具有不同的响应特性。
2.4 化学计量特征与生物量指标间的相关性
对于表4的分析可知4种荒漠植物生物量各指标间与化学计量特征的相关性也没有一致规律,大部分指标间相关性较弱。每种植物的N含量与P含量间均达到了显著水平且都为正相关,除沙蓬和雾冰藜的N∶P之外其余N、P含量及N∶P分别与植物的地上、地下生物量和总生物量间显著相关(负相关),表明过高或过低的N、P含量可能不利于其生物量的分配。
4种植物的R/S、α与N-P化学计量特征间的相关性均未达到显著水平,表明化学计量特征与生物量各指标间较弱的相关性影响着植物整个生长周期。相关系数显示,角果藜和沙蓬N含量的作用要强于P,相反P对碱蓬和雾冰藜生物量的分配作用稍强于N。由此可知,N、P化学计量特征可能对于同一生活型一年生荒漠草本的生长发育以及生物量的分配并不是主要因素。
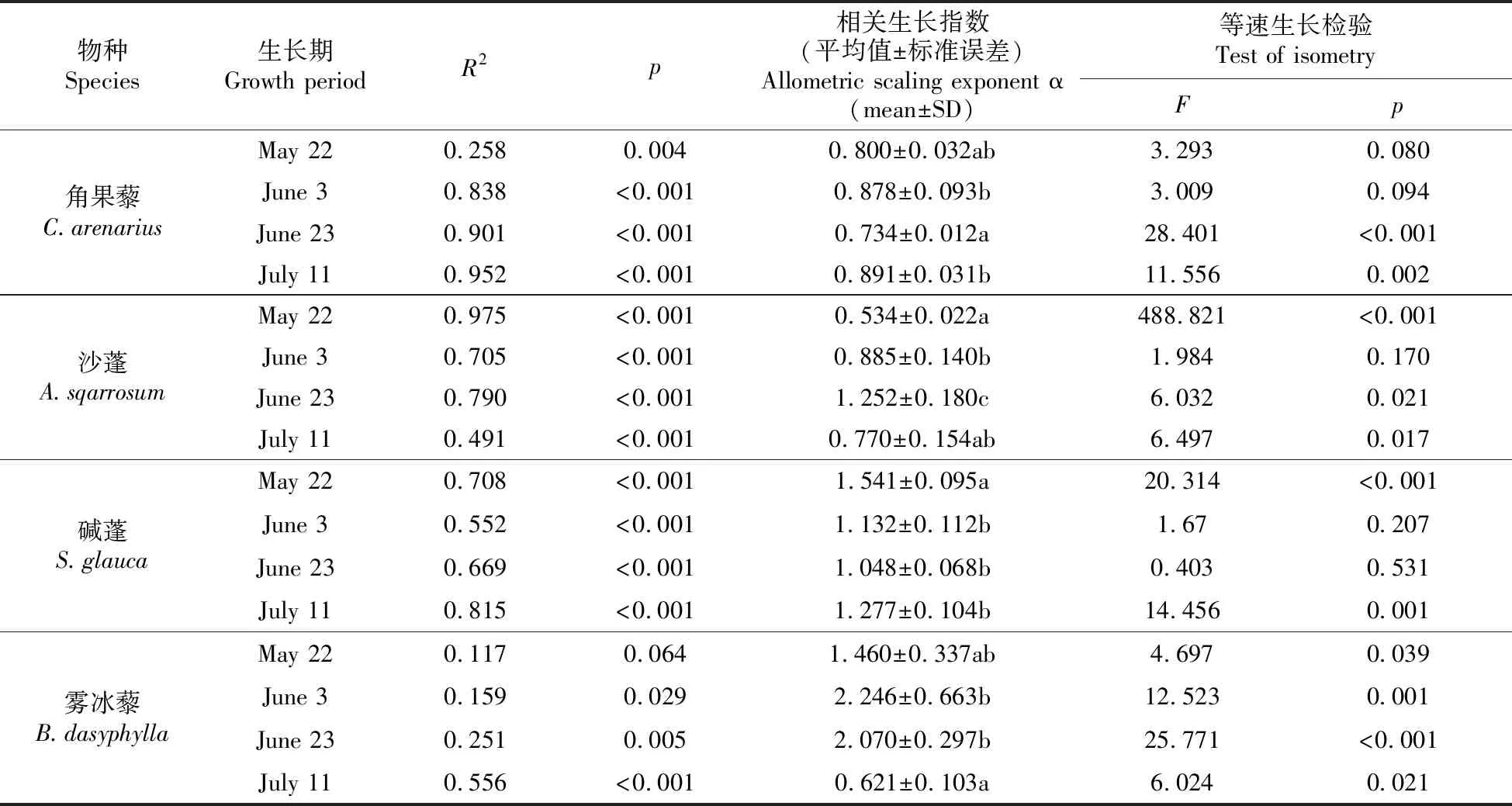
表2 4种草本植物不同生长期相关生长指数统计分析
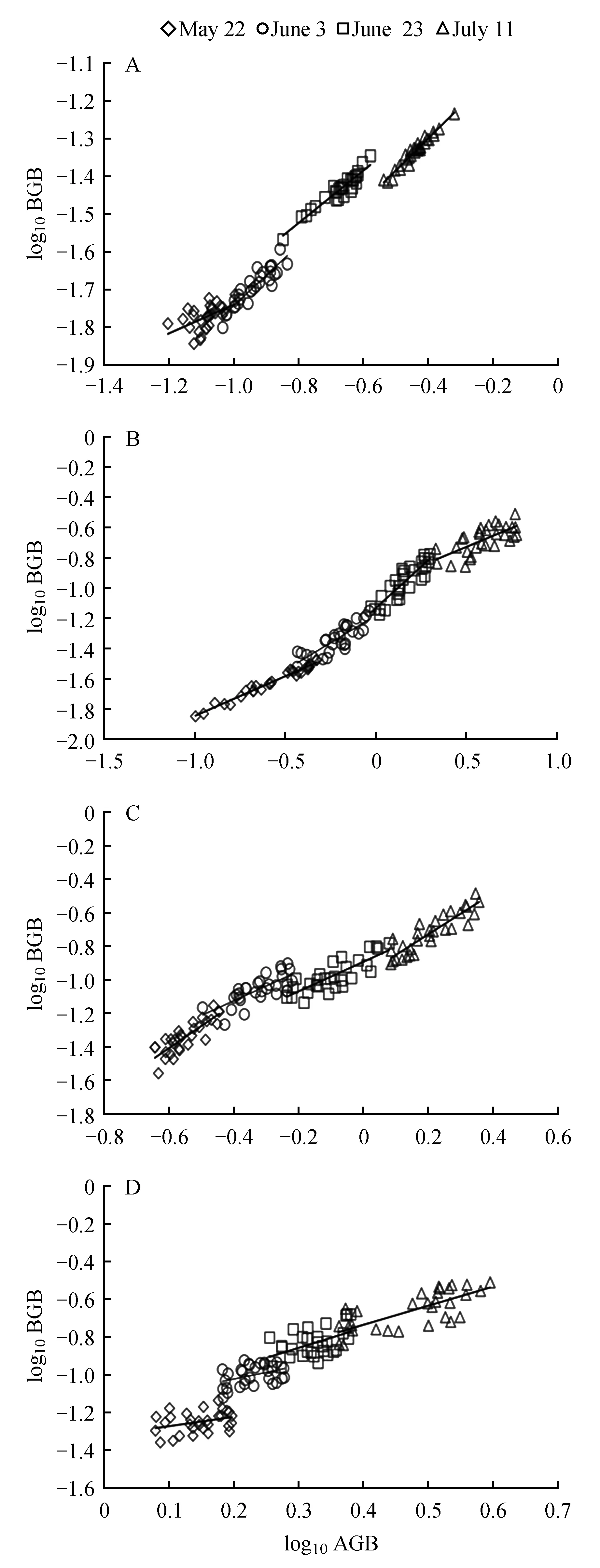
图2 4种荒漠草本植物不同生长期地上生物量(AGB)与地下生物量(BGB)相关生长关系(平均值±标准误) A.角果藜;B.沙蓬;C.碱蓬;D.雾冰藜Fig.2 Correlation between aboveground biomass(AGB) and underground biomass(BGB) in different growth stages of 4 kinds of desert herbaceous plants(mean±SE) A.C.arenarius;B.A.sqarrosum;C.S.glauca;D.B.dasyphylla
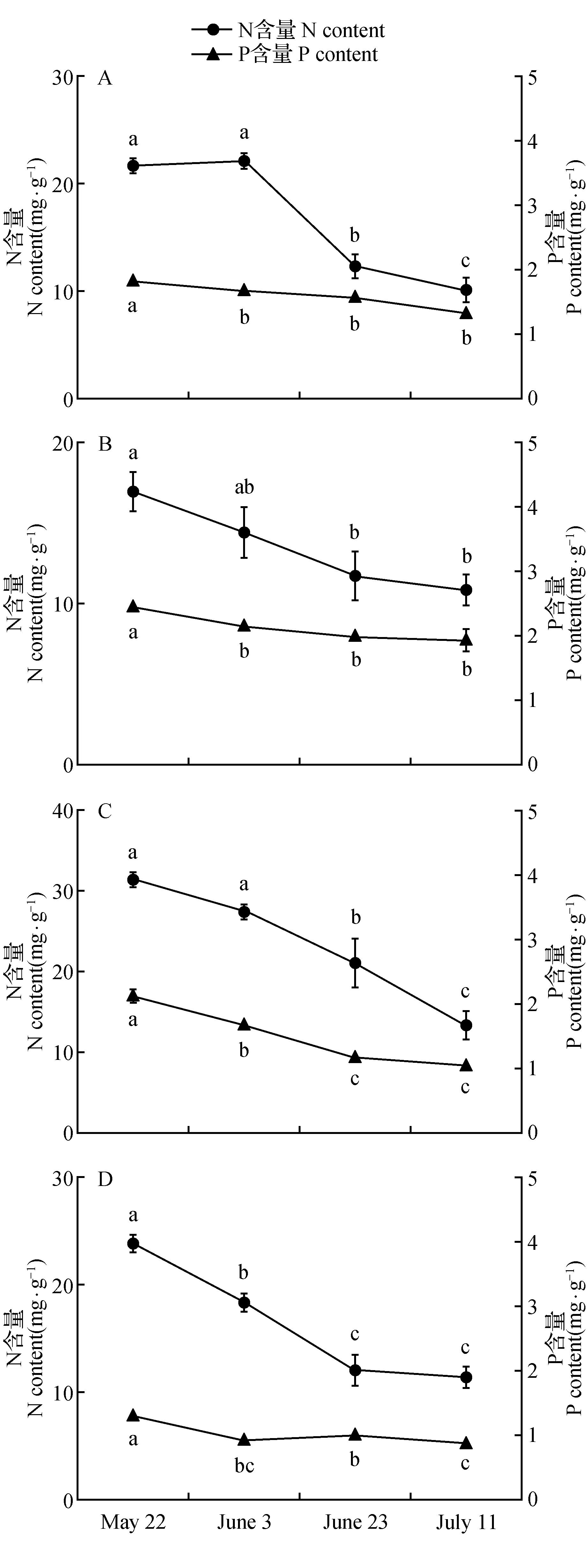
图3 4种荒漠草本植物不同生长期N、P含量(平均值±标准误) A.角果藜;B.沙蓬;C.碱蓬;D.雾冰藜Fig.3 Contents of N and P in four kinds of desert herbaceous plants in different growth periods(mean±SE) A.C.arenarius;B.A.sqarrosum;C.S.glauca;D.B.dasyphylla
Table 3 Variation coefficients of N,P and stoichiometric ratios of four herbs
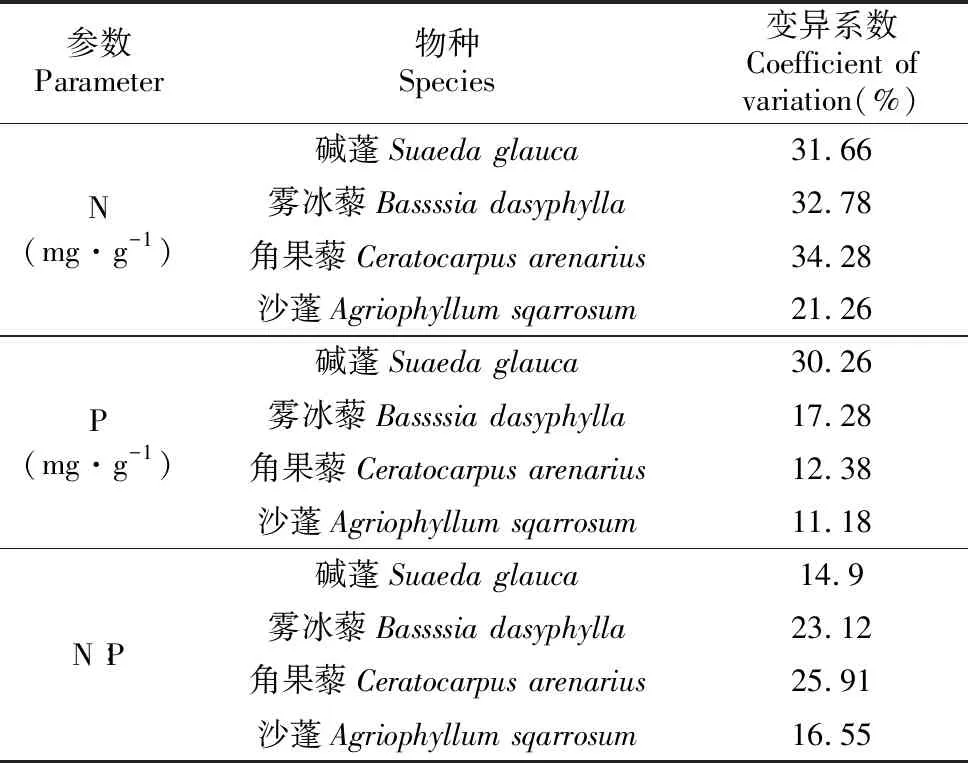
参数Parameter物种Species变异系数Coefficient of variation(%)N(mg·g-1)碱蓬Suaeda glauca31.66雾冰藜Bassssia dasyphylla32.78角果藜Ceratocarpus arenarius34.28沙蓬Agriophyllum sqarrosum21.26P(mg·g-1)碱蓬Suaeda glauca30.26雾冰藜Bassssia dasyphylla17.28角果藜Ceratocarpus arenarius12.38沙蓬Agriophyllum sqarrosum11.18N∶P碱蓬Suaeda glauca14.9雾冰藜Bassssia dasyphylla23.12角果藜Ceratocarpus arenarius25.91沙蓬Agriophyllum sqarrosum16.55
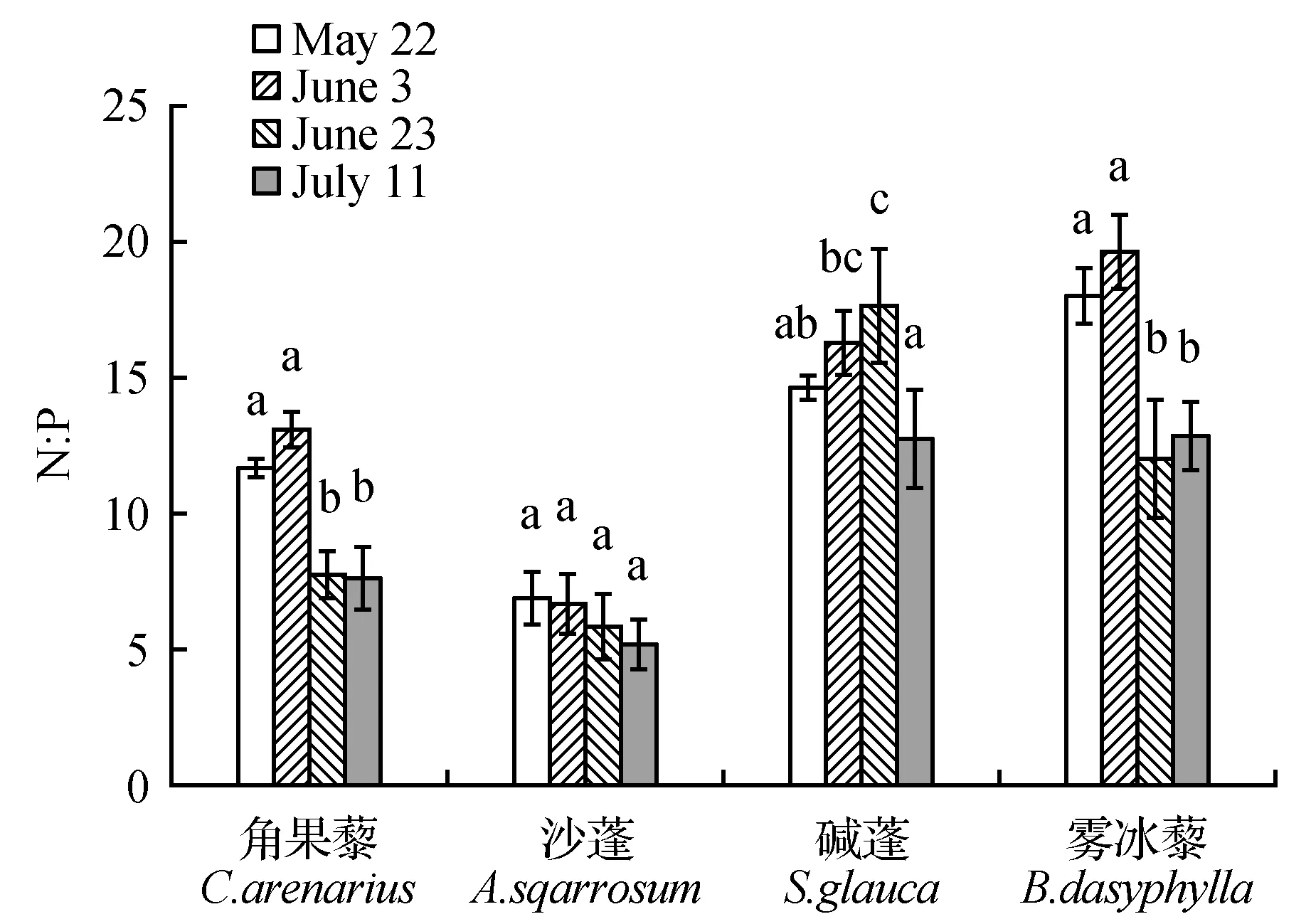
图4 4种荒漠草本植物不同生长期N∶P(平均值±标准误)Fig.4 Four kinds of desert herbs have different growth periods N∶P(mean±SE)
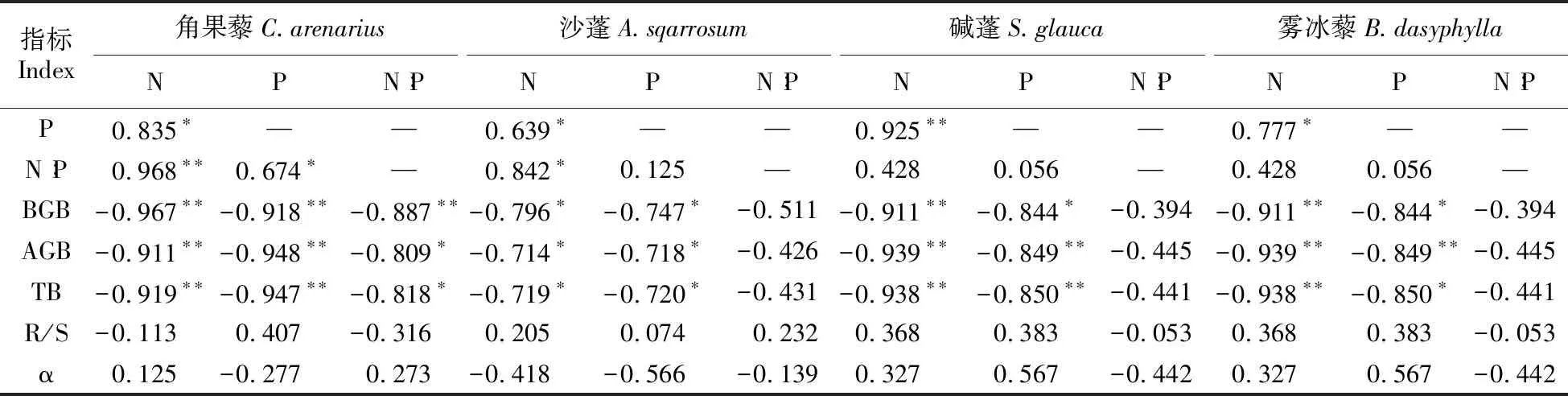
表4 4种草本植物生物量指标与N、P及化学计量比的相关性分析
注:AGB、BGB、R/S、TB和α分别表示地上生物量、地下生物量、根冠比、总生物量和异速生长指数。 *P<0.05;**P<0.01
Note:AGB,BGB,R/S,TB and α represent aboveground biomass,belowground biomass,root to shoot ratio,total biomass and allometric scaling exponent,respectively. *P<0.05;**P<0.01
3 讨论
3.1 生物量分配与相关生长关系
生物量是植物积累能量的主要体现,其在各器官中的分配方式不仅受外界环境的影响,同时也受大小的限制[24]。本研究中,不同植物之间生物量分配比例不同,表明在同一荒漠地区,不同植物积累能量的能力不同,而随着植物的生长,能量在生物体内各器官的积累越来越多[25]。最优分配理论认为,植物可以通过调节生物量在各器官中的分配来适应外界的环境条件,从而最大限度地获取光照、水分和养分等受限的资源,以维持其生长速率的最大化。荒漠植物在不同的生长阶段,如何调整生物量在各器官间的分配比例以适应不利环境,是研究荒漠植物生存策略的重要内容。在极端干旱的荒漠生态系统中,植物常以发达的根系来适应干旱胁迫,一般具有较大的R/S,且R/S随着干旱的加剧而增加[3,21]。在古尔班通古特荒漠地区,降雨小于200 mm,土壤多为风沙土,因此水分和养分成为植物生长的主要限制因子[26]。本研究中,4种荒漠草本植物的根冠比均随个体增大呈减小趋势,且这种趋势在植物生长初期较明显,表明随个体增大植株的地上部分生物量分配比例增加而根系生物量分配比例下降,以满足自身生长与后代繁衍。随着4种植物生长周期的延长,地上生物量在总生物量中占的比例逐渐增大直至生长末期,而R/S也并无显著差异,这也充分说明同一生活型荒漠草本植物在维持自身生长及资源利用方面基本相同。
相关生长关系能够揭示不同物种间生物量分配的基本速率问题。相关生长指数则反映了资源的分配速率,等速生长关系体现了植物构件生物量间具有恒定的分配速率,而异速生长关系则表明生物量分配的不均匀性[21]。本研究中角果藜异速生长关系表现为等速→异速的变化过程;沙蓬从采样初期到末期的变化为异速→等速生长;而沙蓬和碱蓬的生长关系相反,分别由异速→等速→异速、等速→异速→等速的生长过程。4种植物相关生长关系在生长期的变化是异同的。沙蓬与雾冰藜表现近似,相关生长指数增大到一定程度后又逐渐减小并趋于稳定,说明植物将生物量分配到地上和地下的比例是均一的,但两者生物量分配速率并不相同。与之不同的另外两种植物角果藜的相关生长指数增大到一定程度后减小又逐渐增大并趋于稳定,而碱蓬是先减小到一定程度后又逐渐增大并趋于稳定,说明同一种间不同植物生存策略有所差异。一般认为,植物AGB与BGB相关生长关系与植物种类、生活型无关[21,27]。但本研究的结果与这一结论并不完全相同。这可能是因为植物在适应温带荒漠气候环境过程中,形成了不同的资源分配策略,而且这也与植物自身某些遗传特性有关。另外,生物量变化范围狭窄也可能是影响分配的不确定因素,因此还有待进做出一步深入分析。
3.2 化学计量特征在不同时期的变化
本研究4种荒漠植物的生物量分配不仅受水分和温度等多种自然因素的影响,而且体内N、P的含量及化学计量也在整个生长季呈现较大差异(表1)。碱蓬N的含量居4种植物之首,在6月23日前后碱蓬体内的N∶P突然增长,这可能是由于干旱无雨所导致。干旱会导致相应的保护系统得到加强[28],从而促进叶片N含量升高;而高N也会使植物体内细胞液浓度增大[29],提高叶片水势,促进水分吸收,缓解植物干旱胁迫[30]。而P含量在沙蓬中较高且相对稳定,推测6月底至7月处于花果期的沙蓬需要更多的P元素来完成繁殖生长的过程。研究发现4种荒漠植物N∶P含量均随着植物的生长而降低,这主要可能是生物量增大从而使元素含量相对降低的结果。这与牛得草[30]和吴统贵[12]等研究规律相一致。相关实验证实N∶P<14为N限制,若N∶P>16则为P限制。雾冰藜在生长初期受P限制,生长中后期变为受N限制,沙蓬和角果藜整个生长季内均为N限制。而碱蓬在初次取样是受到N、P共同限制,6月的两次取样和末次分别为P、N的限制。在本研究中4种植物N∶P(6.32~15.73)与荒漠区域的非禾草类植物(14.2)[31]及所有旱生草本植物的平均N∶P(13.5)[32]相当,但植物元素的限制类型还是难以判断[33]。综合分析4种荒漠草本植物N、P含量及其计量比的变异,变异系数由大到小依次为N>N∶P>P>C。对N、P含量及N∶P变异系数的大小分析与牛得草[25]和李玉霖[31]所得出的结论略有不同。这可能是由于没有进行大尺度的样品采集与数据分析。另外,4种植物N含量(变化范围为10.17~23.73 mg·g-1)和N∶P(变化范围为5.68~19.73 mg·g-1)都和李玉霖[31]等研究报道相似,但是P含量变化范围0.88~2.44 mg·g-1,却低于其报道结果,这可能是由于土壤养分的不同所造成的差异。因此,少次采样分析并不能真实地反映植物的生态化学计量学特征。所以跨越整个生长期的采样并结合土壤有效成分对生物量的分配模式所做出分析才更为合理。
3.3 化学计量特征对生物量分配的可能影响
生物量分配是研究植物对环境适应策略的一个重要方向[27]。在植物生理生长过程中,也必然会伴随着体内N、P元素的形成,那么元素与生物量积累之间又有如何关系?部分研究表明,化学计量特征能够影响植物生物量分配:高N可以提高叶片光合作用,增加同化产物积累,使R/S显著减小,也即分配到地上部分的生物量比例显著增大;相反低N含量使植物R/S增大[21]。本研究中,4种植物的N、P含量间达到了正相关显著水平,部分N、P含量及N∶P分别与植物的AGB、BGB和TB间呈负相关,且4种植物的R/S、α与N-P化学计量特征间的相关性均未达到显著水平。因此一年生荒漠草本植物生长过程中化学计量比并未对生物量分配起决定作用,一方面可能是干旱环境下植物不同生存策略所引起的结果,另一方面也可能是物种样本量较少,准确性存在偏差。推测植物生长发育过程中自身遗传特性对生物量分配占主体因素。所以对于后续生物量和化学计量比的研究可以选择更多的物种并结合不同阶段土壤营养状况进行更加深入的探讨分析。
猜你喜欢
小哥白尼(趣味科学)(2022年5期)2022-08-15 08:34:46
中国石化(2021年8期)2021-11-05 07:00:14
农业工程学报(2021年23期)2021-03-11 01:11:30
今日农业(2020年19期)2020-12-14 14:16:52
绿色中国(2019年14期)2019-11-26 07:11:34
安徽科技学院学报(2018年5期)2019-01-18 00:36:42
种子(2017年12期)2018-01-17 10:55:45
文史春秋(2016年3期)2016-12-01 05:42:20
中国科技信息(2015年2期)2015-11-16 08:18:32
植物营养与肥料学报(2014年1期)2014-03-11 20:24:38
