
极度濒危树种羊角槭花部形态特征及开花动态
2019-06-06 02:40陈香波吕秀立赵明水崔心红张冬梅
植物研究 2019年3期
陈香波 吕秀立 刘 杨 赵明水 崔心红 张冬梅*
(1.上海市园林科学规划研究院,上海 200232; 2.上海城市困难立地绿化工程技术研究中心,上海 200232; 3.江西农业大学园林与艺术学院,南昌 330045; 4.浙江天目山国家级自然保护区管理局,杭州 311300)
物种濒危固然有其生态历史的原因,但自身的生物学特性和生存能力也不容忽视,因存在生殖系统障碍或者对外界环境不适应、不能够产生正常发育后代个体,也可能导致种群更新不良、数量锐减直至濒危[1]。通过对濒危植物生殖生物学特性及影响其生殖有效性的生理、生态因子进行研究,探讨其内在的濒危机制并找出提高其生殖有效性的技术措施,对于实现濒危物种的有效保护具有十分重要的意义[2]。
羊角槭为槭树科(Aceraceae)槭属(Acer)落叶乔木,为国家二级保护植物、中国为数不多的野外生存数量不足10株的极度濒危树种之一,系古老的残遗种,野外发现时仅残存4株[3~4]。国家林业局与国家发改委联合印发的《全国极小种群野生植物拯救保护工程规划(2011~2015年)》中,羊角槭被明确列入一期拯救保护的120种极小种群植物名录,被IUCN评估为易危种(VN)。近年来由于低温冻害、植株衰退等原因,野外残存的几株羊角槭相继死亡,生存形势异常严峻,亟待加强保护。
迄今,有关羊角槭的研究报道资料甚少,仅有个别关于光合荧光特性以及基因组DNA提取分析亲缘关系方面的报道[5~7]。研究发现羊角槭种子败育率高,饱满种胚率低至4.92%,远不及其它槭属植物,加之存在强烈的种子休眠萌发抑制[8],天然更新能力极弱,截至目前对于其种子不孕率高的生殖机理却知之甚少。
为探明造成羊角槭种子败育率高的生殖生物学原因,本研究以天目山自然保护区人工栽植的羊角槭植株为研究对象,于花期观察了两性花与雄花形态特征,并分别统计了两性花与雄花在枝条中不同位置的分布数量,记录单株与群体水平开花动态,分析羊角槭开花生物学特性与繁育机制、依此提出有针对性的保护措施。

表1 羊角槭花发育分期及对应花器官形态特征
1 研究方法
于2011~2013年观察天目山自然保护区人工栽植的羊角槭花部形态特征,随机摘取30朵正在开放的两性花与雄花,测定冠口直径、花丝长,计算平均数;并于2013年定点5株羊角槭对其开花动态进行了跟踪观测。为进行开花进程统计,首先对不同花发育时期两性花、雄花根据外在形态进行分期(表1),自4月12日~4月15日每日取材统计,其后自4月18日取材之后每隔5天取材一次至4月28日止。于上午10点剪取开花主枝一枝,对上部、中部以及下部开花小枝分别统计各分期两性花和雄花的数量、计算比例,参照典型枝条绘制羊角槭花序示意图;根据每一统计日的两性花、雄花的数量占比绘制各单株的开花进程图。
于10月底种子成熟时采集上述植株的种子剥出种仁,分别统计含饱满种胚、败育种胚以及空瘪的种子数量,计算不同发育状况种胚比率,对比揭示开花生物学与结实之间的关系。
2 研究结果
2.1 羊角槭两性花、雄花形态特征与发育
羊角槭花序顶生,伞房圆锥状(图1:A),复合花序分主轴、一级侧生轴与二级侧生轴三级分支,一、二级侧生轴分支进化为聚伞状花序(图2)。一般两性花多为主轴顶花、一级侧生轴顶花或最末端二级侧生轴顶端(图1:B),雄花位于侧生轴顶端或二级侧生轴顶端或下部(图1:C)。羊角槭花为杂性,雄花与两性花共生于一个花序(图1:A~C),具花萼和花瓣各5片,离生雄蕊8枚。两性花子房成扁球状突出于花盘之上,其上密披绒毛,柱头显著伸出约3~5 mm,先端二裂,弯曲向外翻卷,冠口直径8.34 mm,花丝较短、为4.83 mm(图1:B,D)。两性花具雌蕊,子房上位,2室,每室2枚倒生胚珠,成熟花药不开裂。观察到两种形态雄花,Ⅰ型雄花子房微型、柱头极度短缩呈一小黑点(图1:E),Ⅱ型雄花一团白色绒毛中稍有2裂柱头隐现(图1:F),两种形态雄花差别还体现在:Ⅰ型雄花较Ⅱ型雄花早开放大约3~5天,花盘不凹裂(图1:E),而Ⅱ型雄花花盘较厚、呈辐射状凹裂(图2-1,F),雄花冠口直径8.92 mm,花丝平均长8.85 mm(表2)。所调查的3年中,仅在2011年观察到两种形态的雄花,而2012与2013年只观察到Ⅱ型雄花,Ⅰ型雄花未出现。
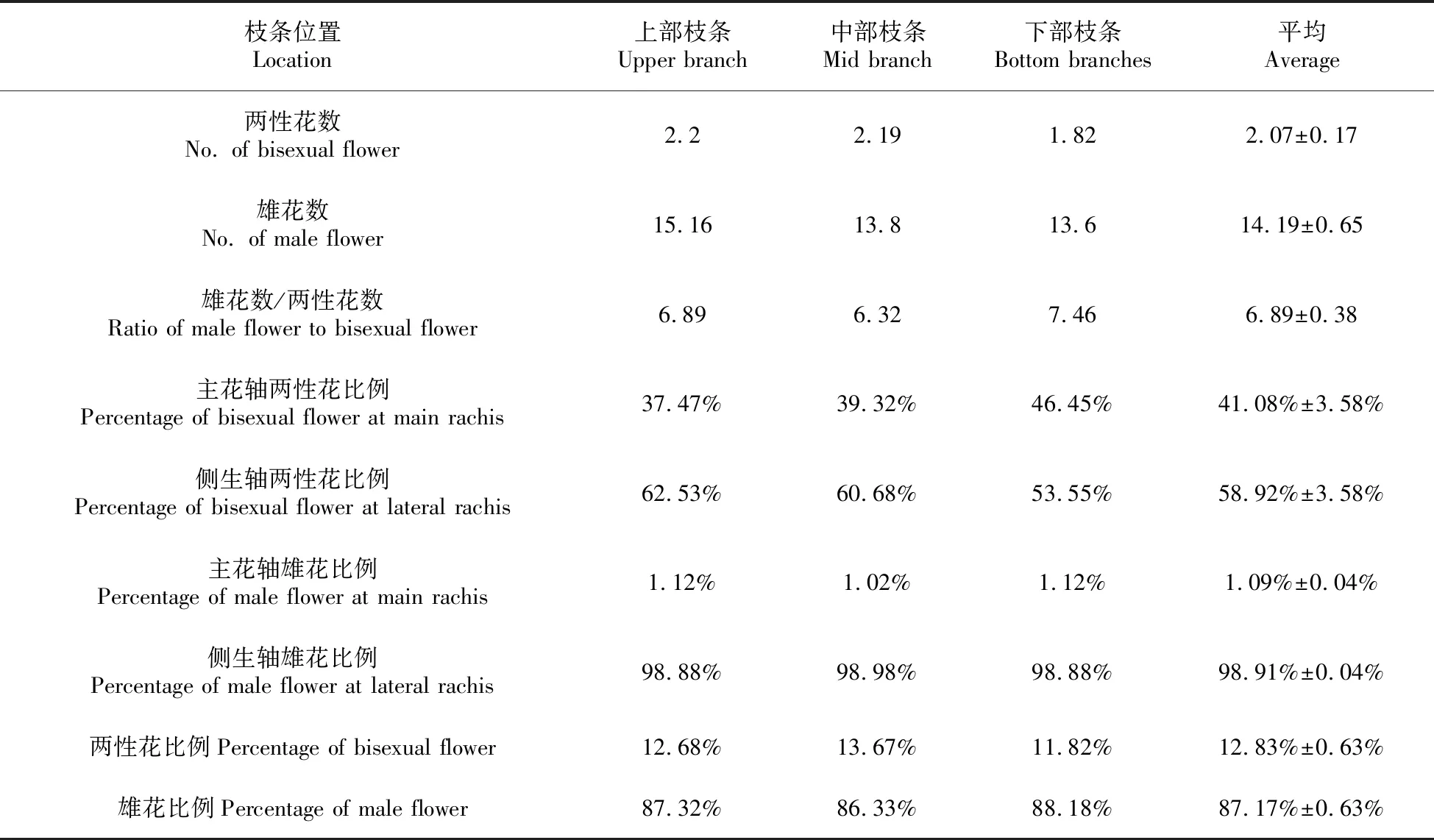
表2 羊角槭两性花、雄花在不同位置枝条中的分布数量及比例

图1 羊角槭花序与花形态特征 A.羊角槭花枝;B.羊角槭花序(箭头所指示两性花);C.羊角槭花序(箭头所指示雄花开放);D.两性花(示雄蕊与雌蕊);E.Ⅰ型雄花.;F.Ⅱ型雄花;G.两性花不同分期;H.雄花不同分期 St.雄蕊;Pi.雌蕊Fig.1 The floral morphology of Acer yangjuechi A.Flowering branch; B.Inflorescence(Arrow pointing to bisexual flower); C.Inflorescence(Arrow pointing to male flower); D.Bisexual flower with stamen and pistil; E.TypeⅠmaleflower; F.TypeⅡmaleflower; G.Different stage of bisexual flower; H.Different stage of male flower St.Stamen; Pi.Pistil
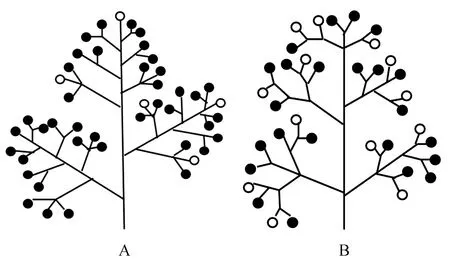
图2 雄花、两性花在花序上的位置分布 A.羊角槭;B.三角槭 实心圆代表雄花,空心圆代表两性花。Fig.2 The distribution of male flower and bisexual flower in inflorescence A.Acer yangjuechi; B.Acer buergerianum Solid circle means male flower,hollowcircle means bisexual flower
单个花序开花动态为:先是主轴顶花开放,其后一级侧生轴顶花、主轴侧生顶花、侧生轴侧生顶花至各轴下部花渐次开放。两性花开放时,花萼张开成近水平状,柱头显著伸出(图1,G:[0]期),此时雌蕊柱头成熟可接受花粉受精时期。开放第二天,花萼收紧包被子房(图1,G:[1]期);随后两性花子房日渐膨大,略带翅形(图1,G:[2]期);单花开放后4~6 d柱头开始萎缩,雄蕊萎蔫脱落,子房明显膨大呈翅形,边沿逐渐由浅绿色变为红色;4月下旬,果实两翅平均间距2 cm(图1,G:[5]期);5月以后,翅果渐伸长,至两翅间距达到4 cm时,花萼、花瓣均已凋谢脱落。雄花初形成时花萼紧裹,宽度仅5 mm以下(图1,H:[-2]期),至4月中旬后,雄花生长到7 mm左右宽度时,花萼逐渐微张(图1,H:[-1]期),直至雄花花萼完全张开与花盘成平面(图1,H:[0]期),此时为雄花花药成熟散粉时期,随后雄花花萼萎蔫、落花凋谢。
2.2 雄花、两性花在枝条上的数量比例与位置分布
观察记录定点枝条花序上的不同性别分化花的数量及在枝条分布的相对位置(上部、中部、下部),统计了位于枝条上部、中部以及下部共计414个花序,其中两性花总数877、雄花总数5 960,雄花比两性花数量比例6.89(表2)。每花序两性花数随花序位置自上而下略有减少(2.2→2.19→1.82),而主花轴两性花比例却是随着花序位置自上而下逐渐增加37.47%→39.32%→46.45%,相应的分支轴两性花比例则有所下降(62.53%→60.68%→53.55%),说明羊角槭枝条上部花序两性花着生比例高于中部和下部枝条,但下部枝条主轴两性花比例较上部和中部枝条居多,而分支轴两性花数量却正好相反,以上部枝条偏多。
与同是槭属植物的三角槭枝条花序上雄花、两性花数量比较发现,三角槭枝条上雄花与两性花数量比为2.63,枝条中两性花比例较羊角槭的6.89增加了2.6倍,表明三角槭枝条中两性花数量较多且更多分布在枝条的外围处易于接受花粉的位置(图2)。
2.3 羊角槭开花动态
2.3.1 单株羊角槭开花进程
以苗圃株羊角槭为例(图3),4月11日还未见有两性花开放,4月12日50%两性花开放,而至4月15日整株已完全看不到未开和正在开放的两性花,都已进入[1]期以后。若对4月12日~4月15日观察到的两性花进行分期,则两性花的发育次序基本为‘花苞(花萼紧密包裹)—花萼微张待开—花萼完全张开与花盘成平面(此时柱头成熟、可接受花粉受精)—花萼包裹、子房膨大—子房膨大、果两端略有翅形’,逐日增进、渐次发育,而后只是翅果形态大小的变化。由图3,苗圃株羊角槭两性花开放时期集中于4月12日~4月14日,而对应于此时的雄花发育却大多数只在花萼紧密包裹、还未成熟的[-2]期,只有极个别雄花(7%以下)花萼完开处于散粉阶段。
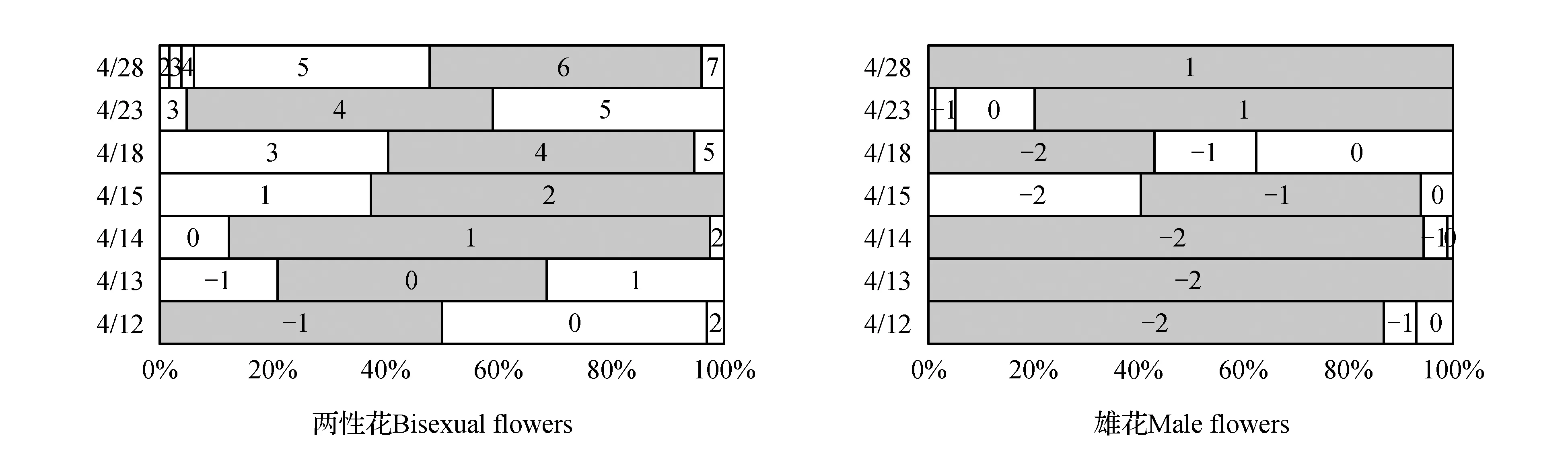
图3 苗圃株花发育进程 图中数字代表不同花发育分期;每一时期分期花数量占比最多者以灰底色标示,代表开花进程处于该分期阶段。Fig.3 Flowering Process of individual located in nursery The number means different developing stage of flowers.The grey base sign the maximum number of flowers which means flowering process at that developing stage.
羊角槭开花进程紧凑,同一植株不同花枝间花发育进程彼此接近。两性花开放时间短,单个两性花花萼自包裹[-1]期至完全展开仅1~2天,两性花柱头为湿型柱头,开放时分泌液体,此时具有可授性,随后花萼收紧、包被子房从而不再接受花粉。柱头成熟可授期也仅在这短暂的1~2天内,整株两性花开放时间3天以内,呈现出集中开放模式。雄花的开放期相对较长,单个雄花经历了‘花萼紧密包裹(花苞阶段)—包裹松动、花萼微张—花萼完全张开与花盘成平面—花萼脱落、雄花凋谢’的发育过程,单花花期约5~6天,而整株雄花花期在10~12天。
2.3.2 群体开花进程
浙江天目山羊角槭正常年份于4月中旬始花,以观测年2013年为计,4月12日只有苗圃株羊角槭有花萼紧密包裹未开花的两性花占多数(50%),处于[-1]分期阶段,此时画眉山庄两株与禅源寺羊角槭尚有少部分[0]期正在开放的两性花(比例分别是20%、12.96%、3.85%),但以[1]期两性花占多数,太子庵株羊角槭已进入[2]期,未见[0]期两性花开放。4月15日,苗圃株两性花开花进入[2]期,此时画眉山庄b株与禅源寺株仍属[2]期,太子庵株已进入[3]期(43.24%)。3天之后的4月18日,多数植株都在[4]期(苗圃、画眉山庄a、禅源寺),画眉山庄b株在[5]期(55.17%)、太子庵株在[6]期(50%)。后期两性花子房膨大生长,形态上只是看到形成的小翅果明显增大,两翅间最大距离加宽,至4月底,翅果已基本可以达到3~4 cm。从单株角度评判,所观察的5株羊角槭两性花开花时期不完全一致,存在先后次序,苗圃株明显落后于其它4株,处于群体两性花开放的末期,其次是画眉山庄两株稍早于苗圃株,处于群体两性花开放的中期,而禅源寺与太子庵处开花时期稍早,处于群体两性花开放的早期。从群体角度考虑,推算天目山栽植羊角槭群体两性花开花盛期应集中在4月11~4月13日的三日之内。
相对于两性花开放,雄花开放期跨越时间较长,自4月12日~4月23日各单株均有雄花开放,而占比多数的开放期在4月18日前后(图4)。所有株在4月18日之前,多数雄花仍处于花萼紧密包裹、花苞阶段的[-2]期或花萼微张、即将开放的[-1]期,处于开放期的雄花相对占少数(50%以下)。苗圃株在4月15日前仅有极个别雄花(6.29%)处于开放散粉期,其它株在4月15日处于正开放散粉的[0]期雄花占比在24.81%(画眉山庄a)~36.69%(太子庵)之间,多数的雄花处于未开放阶段([-2]期或[-1]期),直到4月18日,[0]期开放雄花比例仍不过32.06%(禅源寺)~44%(太子庵),未开放雄花略占多数,至5天后的下一观测日4月23日,大多数雄花已进入花后阶段,花萼脱落、雄花凋谢,仅剩极少数雄花开放。由此推测,4月20日前后1~2天,应是羊角槭雄花盛花期。雄花开花期拉得较长,特别是前期(与两性花成熟重合期)都只是少数雄花散粉。比较群体两性花盛花期(4月11~4月13日)与雄花盛花期(4月19~22日),两者间存在一定的时间间隔。就单个植株而言,以苗圃株为例,两性花大量开放期在4月13日,而雄花大量开花期则在4月20日左右,群体内单株间仅有个别交错的两性花期与雄花期。
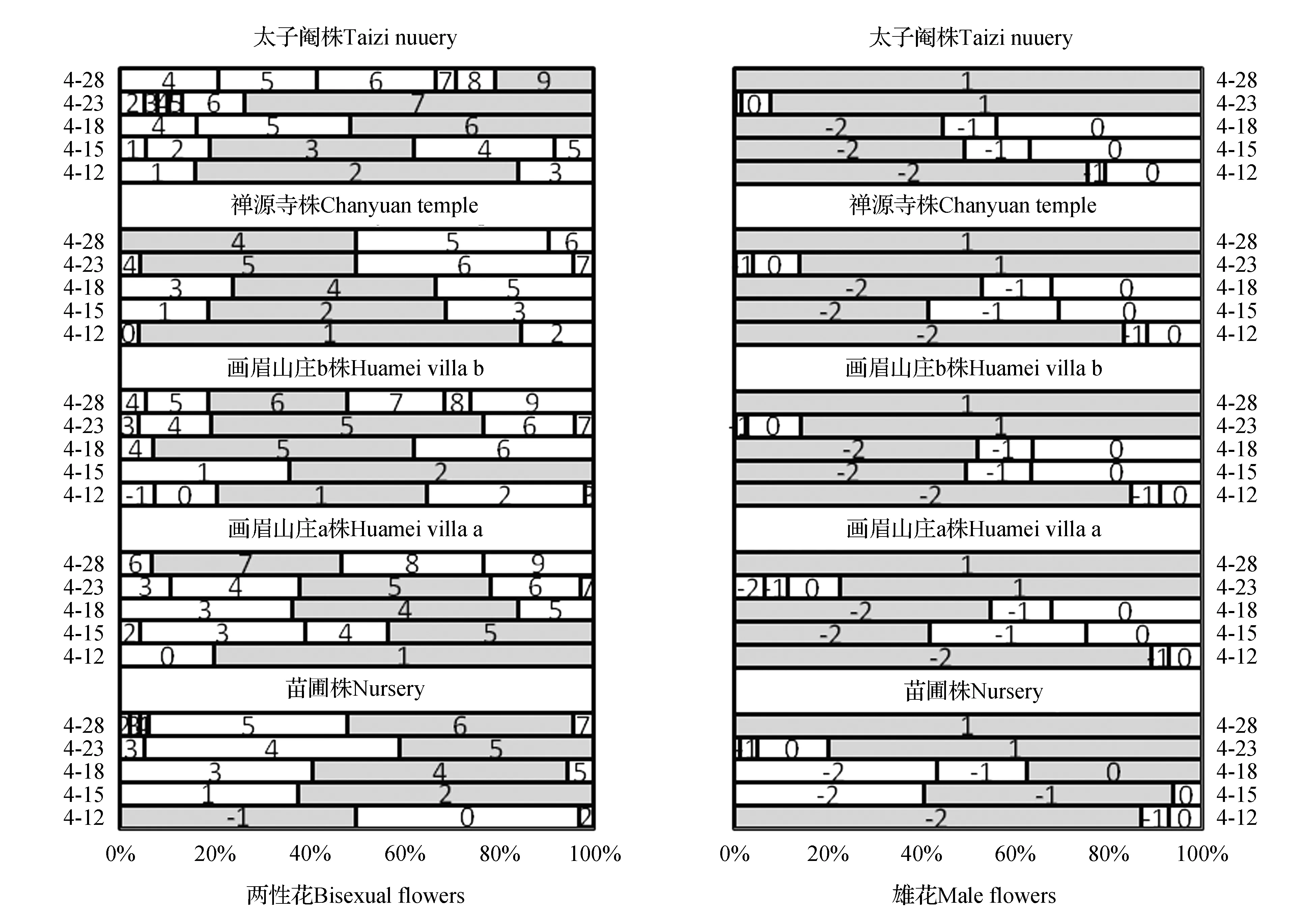
图4 群体内不同单株两性花与雄花发育进程Fig.4 The flowering process of bisexual flower and male flower in different individuals
2.4 羊角槭单株结实情况
对天目山人工栽植羊角槭群体结实情况统计结果发现(图5),画眉山庄两株羊角槭正常种胚率最高,分别为28.83%和26.21%,其次为太子庵与禅源寺羊角槭植株(23.94%和19.7%),而苗圃株正常种胚率仅10.53%,近90%的种子为败育种胚或空籽。
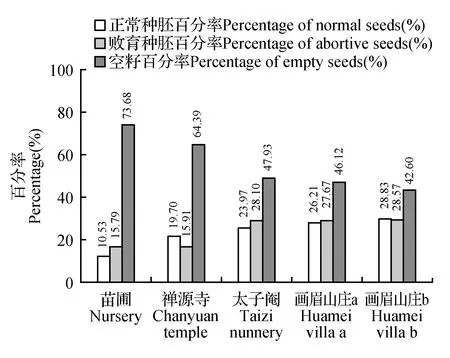
图5 不同单株羊角槭种胚发育情况Fig.5 The embryo development of different individuals
根据Dafni的杂交指数(OCI)标准[9],羊角槭植株上着生两性花与雄花,‘功能性’雌蕊与雄蕊空间上分离,记为1;单个植株存在相对分隔的两性花期(雌性成熟期)与雄花期(雄性成熟期),两性花期短且略先于雄花期,记为1;花朵直径大于6 mm,记为3,由此判定羊角槭的杂交指数为5,则其繁育系统应为异交为主、部分自交亲和,需要传粉者。画眉山庄两株羊角槭相邻生长,正常种胚率高于其它3株(禅源寺、太子庵、苗圃),距离相近促成异交结实利于繁殖成功,与其繁育系统异交为主、部分自交可育是相关联的。
3 讨论
3.1 羊角槭花发育形态特征及与其它槭属植物的比较
槭属植物性别分化复杂,多数种的花表现为杂性、雄花和两性花同株或异株,只有少数种花单性、雌雄异株[10]。二重雌雄异型异熟是槭属植物特化的一种繁育机制,具有两期雄花,两期雄花差别很大,雌花在形态上表现为两性花,植株雌雄功能按照雄—雌—雄顺序依次表达[11~14]。De Jong经过大量形态学和生理学研究之后,将二重雌雄异型异熟归结为单花形态水平上的两性花和单性花,但功能上仅为雄花和雌花[10]。迄今,已在色木槭(A.mono)、挪威槭(A.platanoides)、欧亚槭(A.pseudoplatanus)等多种槭属植物中观察到这一特殊的交配型式样[10,15]。
羊角槭花序顶生,伞房圆锥状花序中同时分布有雄花和两性花,两种花在形态方面有很大差别,两性花子房密披灰白色短绒毛,花柱较长、无毛,中段以上2裂,柱头反卷;而雄花观察到两型,Ⅰ型雄花子房微型、柱头极度短缩呈黑点状,Ⅱ型雄花白色绒毛中稍有2裂柱头隐现,花丝较两性花长。细胞学研究发现,两性花与雄花花药组织相比绒毡层残留多且解体明显,花药成熟时因不具有唇细胞不能裂开而散出花粉,成为结构性的雄性不育[16];而雄花胚珠出现珠心组织退化、双珠被细胞停止发育等退化现象,不能完成正常的胚囊发育,并且Ⅰ型雄花子房退化速度较Ⅱ型雄花更为迅速[17]。与张灵芝等对平基槭、色木槭花发育观察结果相似[15,17],羊角槭在花发育过程中通过成熟花药的不开裂和子房胚珠的败育而分别产生功能性雌花和雄花。除了花的形态和功能多样,就不同类型花在枝条上的分布位置及数量来看,羊角槭两性花数量仅占花序种花总数的12.83%,而雄花占据87.17%,两性花与雄花比例1∶6.89,远低于三角槭两性花在枝条上的分布数量,两性花数量明显偏少。
庙台槭与羊角槭亲缘关系非常近,新版《中国植物志》已将两者归并为1个种[18],最新分子生物学研究通过nrDNA ITS和cpDNA序列比对发现两者间差异较大,分子证据不支持将羊角槭归并为庙台槭[19]。曹小勇等观察发现庙台槭具有两性花、雄花和无性花3种类型的花,两性花花丝较雄花短,雄花成熟时其雌蕊败育、两心皮未合拢呈开裂状,而脱落时仍为花蕾状的花被定义为无性花[20]。就单个花序花的分布与开花动态,庙台槭与羊角槭非常相似,花序的顶花及Ⅰ级分枝轴的顶花由两性花或雄花组成,Ⅱ级分枝轴的顶花近乎全为雄花,个别为两性花,并且均先是顶花与各级分枝轴的顶花开放,由上而下依次进行[21]。对羊角槭连续3年花期观测中,仅在2011年出现两期雄花,而在2012与2013年两性花开放时期未观察到Ⅰ型雄花,庙台槭Ⅲ级分枝轴的顶花由无性花组成并及早脱落[20],是否该脱落无性花即为Ⅰ型雄花,还需做进一步观察比较,以揭示两者间异同,添加新的分类学证据。
3.2 羊角槭开花与授粉结实
单株水平上两性花与雄花成熟数量极不对等,大多数两性花柱头成熟可授粉期时该树体雄花则只有不及10%的雄花处于成熟散粉阶段,只能更多依靠群体内其它个体开放雄花提供花粉,而群体内单株间只存在有限的两性花期与雄花期交错。羊角槭花粉存在部分败育现象,花粉生活力45.97%[16],而两性花又呈集中开放模式,开放时间短。据观察羊角槭花期传粉昆虫以蝇类、娥类居多,大多在晴天的11:00~16:30左右访花,虫媒授粉是其主要的传粉方式[21]。距离较近、相邻生长便于单株间彼此方便提供花粉授粉,种胚发育饱满比例高,在本研究中得到了验证。当然还需进一步进行花粉量、胚珠比观察与计算以及人工控制授粉试验,测定不同授粉处理的结实率以进一步验证其繁育系统。
3.3 交配型变化及其影响因素
据对欧亚槭(Acerpseudoplatanus)的研究发现,不同年份间同一植株会发生交配型间的相互转换,Ⅰ型雄花并不是每一年份都会出现,可能受气候与环境因素影响[10],推测与植株体内的激素调节水平有关[15]。羊角槭在2011年出现两期雄花,而后两年未出现Ⅰ型雄花。单株交配型式样年份间变化情况,需要进一步多年跟踪观测,分析受影响的内、外因素,以更好地揭示交配型与羊角槭有效结实之间的关系。
天目山分布的羊角槭植株分布较为分散,花期有先后,群体中个体间相互授粉受距离障碍与花粉量限制影响,导致子房授粉受精不充分,未受精子房虽继续膨大而成为空粒或败育种子,这可能是导致羊角槭种子发育不良,败育、空瘪严重的最直接原因。因此从提高繁殖效率的角度考虑,应建立适生的羊角槭保护小区,缩短种植间距、增加单株间花粉供给,扩大异交结实成功率,以实现羊角槭这一极小种群植物可持续保育的目标。
猜你喜欢
金沙江文艺(2022年4期)2022-04-26
西北农林科技大学学报(自然科学版)(2021年5期)2021-05-31
江苏农业科学(2020年6期)2020-05-21
江苏农业科学(2020年21期)2020-03-08
中国瓜菜(2019年8期)2019-09-19
传奇·传记文学选刊(2018年11期)2018-11-26
草堂(2018年11期)2018-11-13
金山(2018年4期)2018-04-26
天津农业科学(2017年12期)2018-01-10
阅读与作文(小学低年级版)(2016年10期)2016-10-12
