
诸葛菜种子抗真菌蛋白的提取及其抑菌活性
2019-05-30 08:10张玮玮袁素素王财成叶秀娟
福建农林大学学报(自然科学版) 2019年3期
张玮玮,袁素素,王财成,叶秀娟
(福建农林大学生物农药与化学生物学教育部重点实验室,福建福州350002)
抗菌蛋白是植物在长期进化过程中产生的具有抗微生物活性的一类防御性蛋白[1],是机体天然免疫系统的重要组成部分.由于抗菌蛋白具有抗菌谱广、不易产生耐药性、低毒性等特点,极有可能成为一类新型抗菌素或植物源杀菌剂[2].众多研究人员已经从多种植物中分离出抗菌蛋白,并对其结构、功能和作用机理等进行了研究.例如:Ye et al[3]从甘蓝(Brassica oleracea)种子中分离出一种分子质量约为30 ku的蛋白,其对落花生球腔菌(Mycosphaerella arachidicola)、白色念珠菌(Candida albicans)和铜绿假单胞菌(Pseudomonas aeruginosa)均具有抑制作用,且该蛋白可以抑制鼻咽癌细胞和肝癌细胞的增殖;Lin et al[4]从芥蓝(Brassica alboglabra)中纯化出的抗真菌蛋白(5 907 u)能够有效抑制尖孢镰刀菌(Fusarium oxysporum)、玉蜀黍长蠕孢(Helminthosporium maydis)、落花生球腔菌和苹果黑腐皮壳菌(Valsa mali)的生长,还可以抑制肝癌细胞HepG2和乳腺癌细胞MCF7的增殖;Lin et al[5]还从芸薹(Brassica campestris)中分离出抗真菌蛋白campesin(9.4 ku),该蛋白可以抑制尖孢镰刀菌和落花生球腔菌的生长,对HepG2和MCF癌细胞的增殖也有抑制作用.随着研究的深入,这些天然活性产物的应用将更加广泛.
诸葛菜(Orychophragmus violaceus)系十字花科诸葛菜属,因农历二月前后开蓝紫色花,又称二月蓝、翠紫花[6].诸葛菜营养价值较高,其根、茎和叶均可食用[7].诸葛菜种子富含维生素B1、维生素B2、维生素C和胡萝卜素以及铁、钙等矿质元素[8-11];其含油量高达35.8%,其中,棕榈酸、油酸和亚油酸含量较高,而亚麻酸和芥酸含量低,是非常优质的油料资源[12-14].目前,学者们已从十字花科多种植物如大白菜[15]、高菜[16]等中分离出抗真菌蛋白,但是有关诸葛菜种子中的抗真菌蛋白还未见报道.因此,本研究以诸葛菜种子为试验材料,分离纯化其抗真菌蛋白,并测定该蛋白的抑菌活性和稳定性,为促进诸葛菜抗真菌蛋白的应用提供参考.
1 材料与方法
1.1 材料与仪器
诸葛菜种子:购买于江苏省宿迁市四季青种业有限公司.
15种供试植物病原真菌:尖孢镰刀菌、稻瘟病菌(Magnaporthe oryzae)、灰葡萄孢菌(Botrytis cinerea)、芭乐炭疽病菌(Colletotrichum gloeosporioides)、禾谷镰刀菌(Fusarium graminearum)、长柄链格孢菌(Alternaria longipes)、烟草炭疽病菌(Colletotrichum micotianae)、玉蜀黍长蠕孢、小白菜炭疽(Colletotrichum higginsianum)、落花生球腔菌、突脐蠕孢菌(Exserohilum turcicum)、茄病镰刀菌(Fusarium solani)、苹果黑腐皮壳菌、辣椒疫霉(Phytophthora capsici)、小孢拟盘多毛孢(Pestalotiopsis microspora)均由福建农林大学生物农药与化学生物学教育部重点实验室提供.
仪器:Centrifuge 5860R高速冷冻离心机(Eppendorf公司),LGJ-18S冷冻干燥机(上海松源华兴科技发展有限公司),AKTA pure快速蛋白纯化系统(瑞典GE公司),SPX-250智能恒温培养箱(宁波赛福实验仪器厂).
1.2 抗真菌蛋白的分离、纯化
取150 g诸葛菜种子,加入20 mmol·L-1醋酸铵缓冲液(pH 4.6)800 mL,经豆浆机粉碎后置于4℃匀浆浸提12 h,离心(8 000 r·min-1,30 min)后取上清液备用.将上清液上样于SP-Sepharose阳离子交换层析柱(5.0 cm×20 cm),依次用含0.2、0.5和1.0 mol·L-1NaCl的20 mmol·L-1醋酸铵缓冲液进行梯度洗脱,在280 nm波长下检测洗出液的光密度.将各组分收集,以尖孢镰刀菌为指示菌检测各组分抑菌活性.
将有活性的组分透析,冷冻干燥后进一步用Mono STM5/50 GL强阳离子交换柱分离,该柱预先用20 mmol·L-1醋酸铵缓冲液平衡,用0~1.0 mol·L-1NaCl梯度洗脱,收集各洗脱峰,以尖孢镰刀菌为指示菌检测各洗脱峰抑菌活性.
将有活性的组分透析并冷冻干燥,用20 mmol·L-1醋酸铵缓冲液溶解后上样于SuperdexTMPeptide 10/300 GL凝胶层析柱,收集各洗脱峰,以尖孢镰刀菌为指示菌检测各洗脱峰抑菌活性,并收集活性组分,透析并冷冻干燥.
1.3 蛋白分子质量测定
Tricine-SDS-PAGE电泳方法参考文献[3],分离胶浓度16.5%,夹层胶浓度10%,浓缩胶浓度4%.初始电压为30 V,电泳1 h后调至80 V,再继续电泳2.5 h.电泳结束后,用考马斯亮蓝R-250染色2 h,再用脱色液(10%甲醇和10%乙酸)进行脱色.
1.4 蛋白质谱分析
将电泳检测为单一条带的样品经胰蛋白酶酶解后,用LCMSMS(nanoLC-QE)进行质谱分析,通过MASCOT等质谱匹配软件分析LCMSMS数据,通过UniProtKB数据库检索、比对、鉴定分离得到的蛋白质.质谱分析由上海中科新生命生物科技有限公司完成.
1.5 蛋白的抗真菌活性检测
用纸片扩散法[17]进行测定.将待测的植物病原真菌接种于PDA培养基上,28℃恒温培养,待菌落生长至圆面直径为3.0~3.5 cm时,将灭菌的圆滤纸片放在距离菌丝边缘0.5 cm处,各取15 μL蛋白样品和对照(PBS缓冲液,pH 7.4)加在滤纸片上,28℃恒温培养至菌落生长到滤纸片边缘时,观察有无抑菌活性.
1.6 蛋白对尖孢镰刀菌生长半抑制浓度(IC50)的测定
采用琼脂平板稀释法[17]进行测定.将蛋白样品用PBS缓冲液配制成一系列浓度,并经0.22 μm滤头过滤除菌.加热融化0.7%PDA培养基,待冷却至40~50℃时,取0.5 mL蛋白溶液(对照组为0.5 mL PBS缓冲液)与1.5 mL培养基于培养皿(30 mm×15 mm)中混匀,用打孔器截取相同大小真菌菌块,置于培养皿中心,封口并转移至28℃恒温培养,直至对照组菌落生长至培养皿边缘.用十字交叉法测量真菌菌落的生长直径,并计算该蛋白对尖孢镰刀菌生长的抑菌率,每组3个重复.运用SPSS软件对数据进行Probit回归分析,计算IC50.

1.7 蛋白对尖孢镰刀菌孢子萌发IC50的测定
1.7.1 孢子制备 将尖孢镰刀菌接种于PDA培养基上,28℃黑暗条件下恒温培养7~15 d,取5 mL无菌水加入上述培养皿中,用涂布器在真菌表面来回刮动,滤除菌丝得到孢子悬液.稀释孢子悬液至浓度为1×106~1×107CFU·mL-1,取等体积孢子悬液与0.05%葡萄糖溶液混合备用.
1.7.2 孢子萌发抑制率的测定 用凹玻片法[18-19]进行测定.将蛋白样品用PBS缓冲液配制成一系列浓度,并经0.22 μm滤头过滤除菌.取10 μL孢子悬浮液(对照组为PBS缓冲液)与10 μL蛋白溶液混匀,加在凹玻片中央,置于含有两层浸湿的无菌滤纸片的培养皿(90 mm×15 mm)中,28℃恒温保湿培养至对照组孢子萌发率达到90%以上时,将各培养皿放入4℃冰箱中.6 h后用光学显微镜观察并记录孢子萌发数,每组至少观察100个孢子,每组3个重复,计算孢子萌发抑制率.运用SPSS软件对数据进行Probit回归分析,计算IC50.

1.8 蛋白的稳定性检测
1.8.1 热稳定性 取5个1.5 mL离心管,分别加入抗真菌蛋白(用PBS缓冲液溶解,pH 7.4)0.1 mL,其中4管分别在40、60、80、100℃条件下水浴15 min(待冷却至室温时进行试验),1管未处理作为阳性对照,另取1个离心管加入PBS缓冲液0.1 mL作为阴性对照.以尖孢镰刀菌作为指示菌,采用纸片扩散法进行抑菌试验,每组设3个重复.
1.8.2 pH稳定性 取7个1.5 mL离心管,分别加入抗真菌蛋白(用PBS缓冲液溶解,pH 7.4)0.1 mL,其中6管分别加入调配好的pH值为1、3、5、10、12、14的PBS缓冲液0.1 mL,1管未处理作为阳性对照,另取1个离心管加入PBS缓冲液0.1 mL作为阴性对照,室温放置1 h.抑菌试验方法同1.8.1.
1.8.3 有机溶剂稳定性 取6个1.5 mL离心管,分别加入抗真菌蛋白(用PBS缓冲液溶解,pH 7.4)0.1 mL,其中5管分别加入用PBS缓冲液配制的15%(v/v)甲醇、乙醇、异丙醇、乙醚、丙酮溶液0.1 mL,1管未处理作为阳性对照,另取5个离心管分别加入15%甲醇、乙醇、异丙醇、乙醚、丙酮0.1 mL作为阴性对照,室温放置1 h.抑菌试验方法同1.8.1.
1.8.4 金属离子稳定性 用 PBS 缓冲液分别配制 50、100、150、200、250 mmol·L-1的 Mg2+、K+、Mn2+、Ca2+溶液.取6个1.5 mL离心管,分别加入抗真菌蛋白(用PBS缓冲液溶解,pH 7.4)0.1 mL,其中5管分别加入50、100、150、200、250 mmol·L-1Mg2+溶液 0.1 mL,1 管未处理作为阳性对照,以 250 mmol·L-1Mg2+溶液作为阴性对照,室温放置1 h.K+、Mn2+、Ca2+溶液按照同样的方法进行处理.抑菌试验方法同1.8.1.
1.9 数据处理
用Excel和SPSS 17.0软件对数据进行分析处理.
2 结果与分析
2.1 抗真菌蛋白的分离、纯化
诸葛菜种子蛋白上清液经SP-Sepharose阳离子交换柱分离后,得到4个洗脱组分a、b、c、d(图1A);以尖孢镰刀菌为指示菌,用滤纸片法测得b组分具有抗真菌活性(图1B),所以将b组分进一步纯化.b组分经Mono STM5/50 GL强阳离子交换柱分离后,得到5个洗脱峰e、f、g、h、i(图1C);抑菌活性检测结果显示g洗脱峰具有抗真菌活性(图1D),所以选择g组分进一步纯化.将g组分上样于SuperdexTMPeptide 10/300 GL凝胶层析柱进一步分离,得到j、k、l、m共4个洗脱峰(图1E);抑菌活性检测结果表明l洗脱峰具有抗真菌活性(图1F).
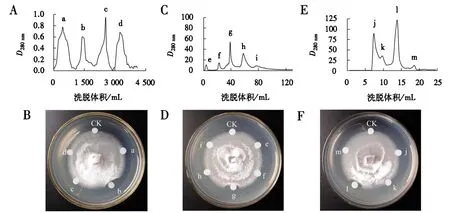
图1 诸葛菜种子抗真菌蛋白的分离层析及各组分活性检测结果Fig.1 Column chromatography and activity detection of the antifungal protein from O.violaceus seeds
Tricine-SDS-PAGE电泳结果(图2)表明,诸葛菜种子抗真菌蛋白经过3步分离纯化后,Ⅰ组分蛋白呈单一条带,说明已达到较高纯度,其相对分子质量约为10 ku,将该蛋白命名为Zp.
2.2 质谱分析结果
将Tricine-SDS-PAGE电泳得到的单一条带进行质谱分析,在UniProtKB数据库中检索,结果如表1所示.经蛋白相似度和序列比对发现,Zp蛋白可能是诸葛菜种子中的一种防御素蛋白(defensin),在UniProtKB数据库中的序列号为C9WGT1,相对分子质量为8 737.17 u,这与电泳得出的Zp蛋白分子质量较为接近.
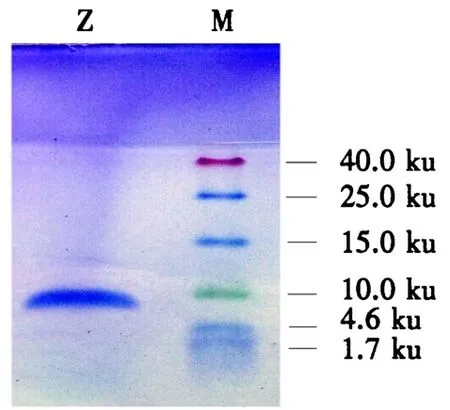
图2 诸葛菜种子抗真菌蛋白的Tricine-SDS-PAGE电泳结果Fig.2 Tricine-SDS-PAGE electrophoresis of the antifungal protein from O.violaceus seeds
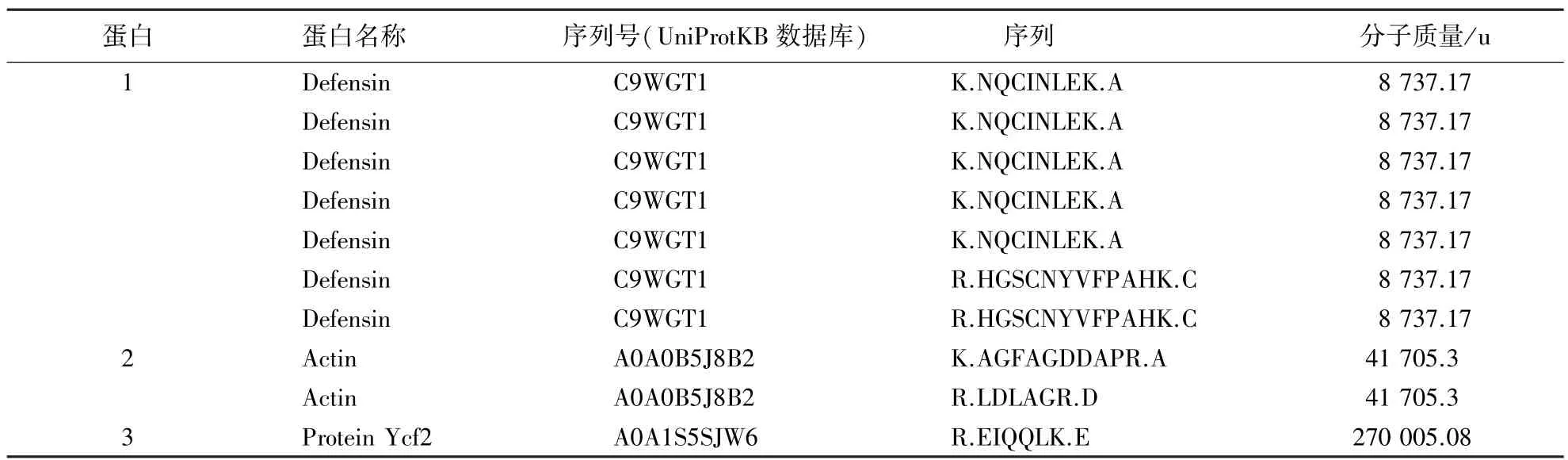
表1 Zp蛋白的质谱鉴定与UniProtKB数据库检索结果Table 1 Mass spectrometry and database retrieva(UniProtKB)of Zp protein
2.3 Zp蛋白的抗真菌活性
Zp蛋白的抗真菌效果如图3所示,其对稻瘟病菌、灰葡萄孢菌、突脐蠕孢菌、烟草炭疽病菌、苹果黑腐皮壳菌、芭乐炭疽病菌、小白菜炭疽、玉蜀黍长蠕孢、落花生球腔菌、小孢拟盘多毛孢和尖孢镰刀菌共11种植物病原真菌均具有抑制作用,而对茄病镰刀菌、辣椒疫霉、长柄链格孢菌和禾谷镰刀菌等4种真菌没有明显的抑制作用.由此可知,诸葛菜种子抗真菌蛋白具有一定的广谱性.
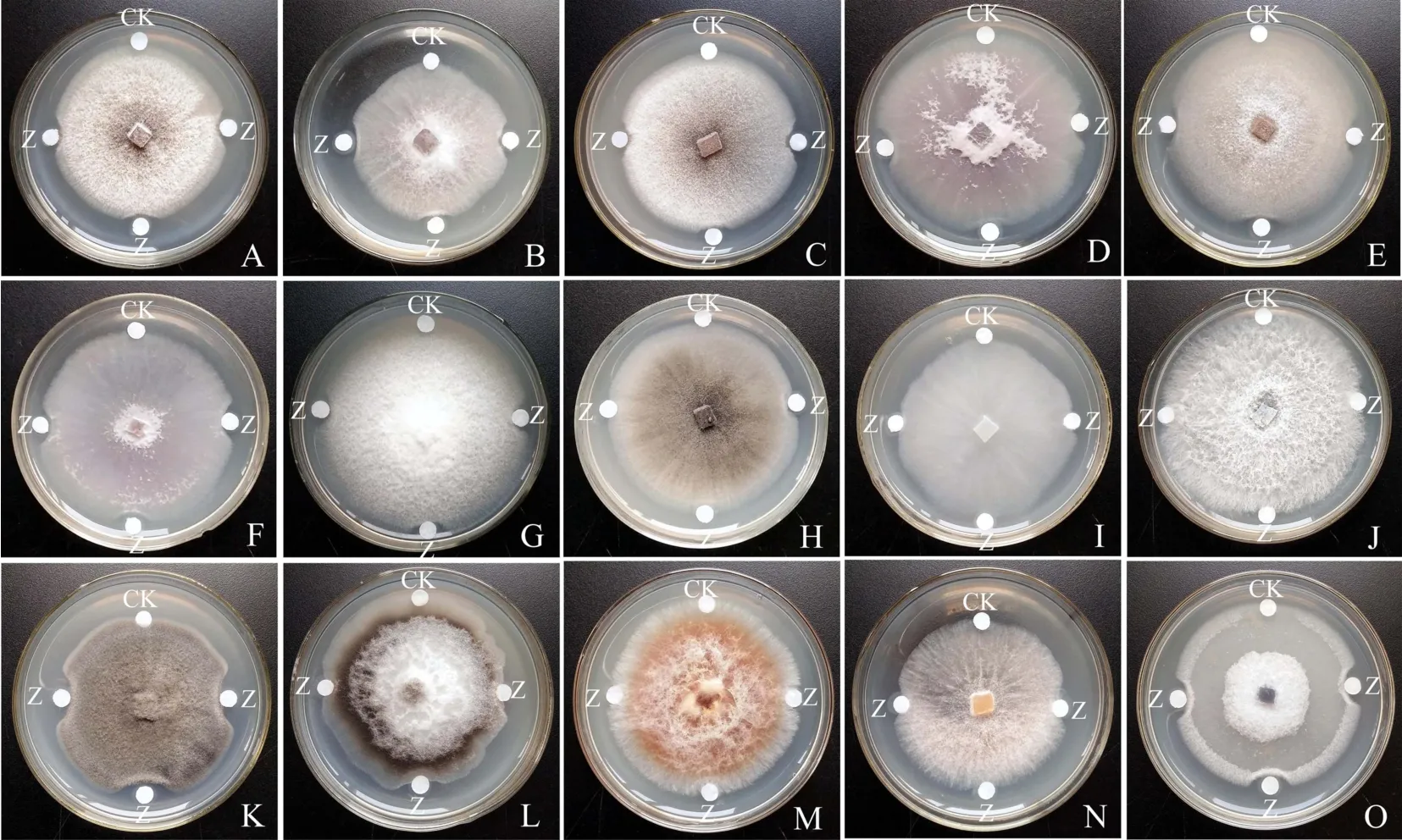
图3 Zp蛋白的抗真菌效果Fig.3 Antifungal effects of the Zp protein from O.violaceus seeds
2.4 Zp蛋白对尖孢镰刀菌生长的IC50
Zp蛋白对尖孢镰刀菌生长的抑制效果如图4A所示.随着Zp蛋白浓度的增大,对尖孢镰刀菌生长的抑制率逐渐升高,当Zp蛋白浓度为142.5 μg·mL-1时,对尖孢镰刀菌的抑制率超过60%(图4B).Zp蛋白对尖孢镰刀菌生长抑制的Probit回归方程:Probit(p)=-1.530+0.882x.由此得出Zp蛋白对尖孢镰刀菌生长的 IC50为 54.36 μg·mL-1.
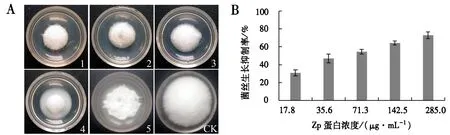
图4 Zp蛋白对尖孢镰刀菌生长的抑制效果Fig.4 Inhibtory effects of Zp protein on the growth of F.oxysporum
2.5 Zp蛋白对尖孢镰刀菌孢子萌发的IC50
Zp蛋白可以抑制尖孢镰刀菌孢子的萌发:与对照组相比,处理组孢子萌发数显著减少,且萌发的牙管较短,有的孢子形态膨大加粗,甚至出现畸形(图5A).随着Zp蛋白浓度增大,对尖孢镰刀菌孢子萌发的抑制率逐渐升高,当蛋白浓度为18.85 μg·mL-1时,孢子萌发抑制率达到60%(图5B).Zp蛋白对尖孢镰刀菌孢子萌发抑制的Probit回归方程:Probit(p)=-1.449+1.204x.由此得出Zp蛋白对尖孢镰刀菌孢子萌发的 IC50为 15.97 μg·mL-1.
2.6 Zp蛋白的稳定性
从图6A可看出,Zp蛋白在40、60、80、100℃水浴处理15 min后,对尖孢镰刀菌的抑制活性基本保持不变,表明该蛋白的热稳定性良好.由图6B可知,Zp蛋白在经不同pH值的溶液处理后,对尖孢镰刀菌仍有较强的抑制作用,说明该蛋白的耐酸碱性较强.
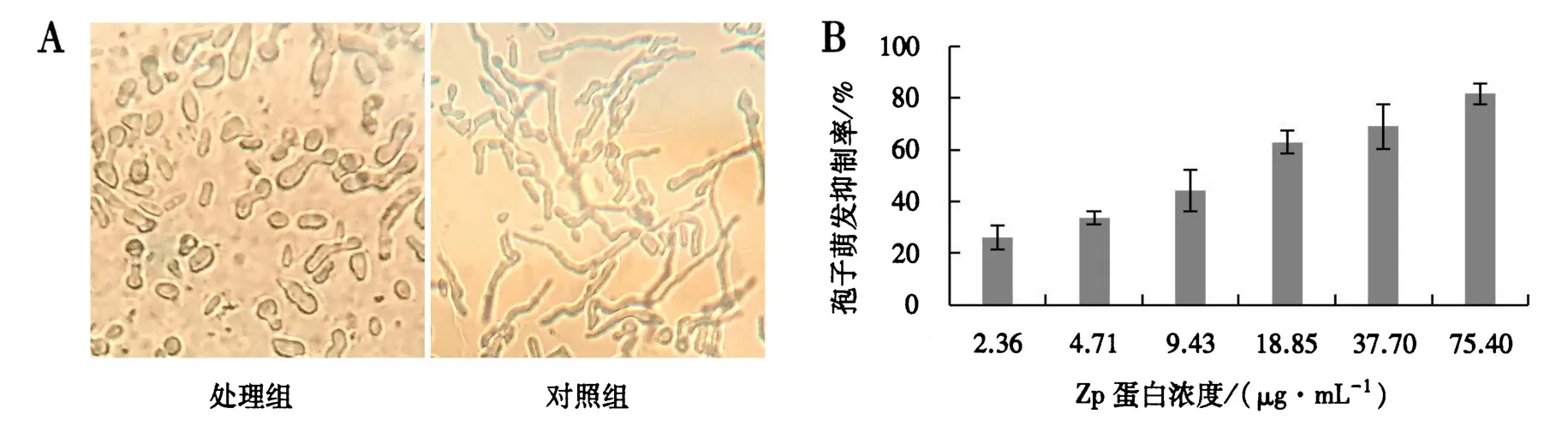
图5 Zp蛋白对尖孢镰刀菌孢子萌发的抑制效果Fig.5 Inhibtory effects of Zp protein on the germination of F.oxysporum spores
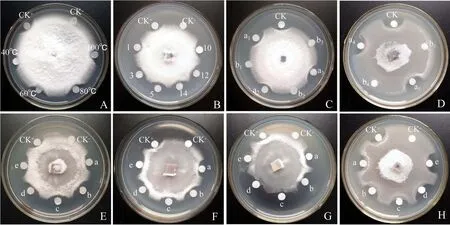
图6 Zp蛋白的稳定性Fig.6 Stability of Zp protein
由图6C、6D可知,经15%甲醇、乙醇、异丙醇、乙醚、丙酮溶液处理过的Zp蛋白能够明显抑制尖孢镰刀菌的生长,而单独的相应有机溶剂对尖孢镰刀菌生长并没有抑制作用,由此说明该蛋白对有机溶剂具有一定的耐受性.由图 6E、6F、6G、6H 可知,不含 Zp 蛋白的 250 mmol·L-1Ca2+、Mg2+、Mn2+、K+溶液对尖孢镰刀菌生长没有抑制作用,而含有该蛋白的不同浓度(50、100、150、200、250 mmol·L-1)金属离子溶液对尖孢镰刀菌生长具有明显的抑制作用,说明Zp蛋白在强阳离子溶液中活性稳定.
3 讨论与结论
本研究从诸葛菜种子中分离出一种分子质量约为10 ku的抗真菌蛋白Zp.Zp蛋白对多种植物病原真菌均具有抑制作用,其中对尖孢镰刀菌生长的IC50为54.36 μg·mL-1,对其孢子萌发的IC50为15.97 μg·mL-1.抗真菌蛋白存在于十字花科的多种植物种子中,且这些蛋白对多种植物病原真菌具有抑制活性.例如:油菜(Brassica napus)种子中分离出的抗真菌蛋白(9 412 u)对尖孢镰刀菌和落花生球腔菌的IC50分别为78.12和42.35 μg·mL-1[20];甘蓝种子中纯化出分子质量为30 ku的抗真菌蛋白,对落花生球腔菌的IC50为150 μg·mL-1[3];芥蓝种子中纯化出分子质量为5 907 u的抗真菌蛋白,对尖孢镰刀菌、玉蜀黍长蠕孢、落花生球腔菌和苹果黑腐皮壳菌的IC50分别为25.40、12.40、14.18和0.89 μg·mL-1[4].由此可知,Zp蛋白的抗真菌活性较强.经质谱鉴定,Zp蛋白可能是诸葛菜种子中的一种防御素蛋白.Chan et al[21]已经从褐色芸豆(Phaseolus vulgaris)的种子中分离出一种防御素类蛋白,分子质量为5.4 ku,该蛋白也具有抗真菌活性.防御素类蛋白是植物抗菌蛋白中数目最多的一个种类且具有广谱的抗菌活性[22].通过对Zp蛋白的抑菌活性进行检测可知,其对尖孢镰刀菌等11种真菌都有较强的抑制活性,表明该蛋白具有抑菌广谱性.防御素类蛋白还具有典型的三维稳定结构[23].检测可知,Zp蛋白具有良好的耐热、耐酸碱性;已有研究发现,油菜种子抗菌蛋白和芥蓝种子抗菌蛋白也具有热稳定性和强酸强碱稳定性[4,24-25].Zp蛋白对有机溶剂和金属离子也具有一定的耐受能力;Terras et al[26]研究表明,萝卜(Raphanus sativus)种子Rs-AFP1和Rs-AFP2抗菌蛋白在K+溶液(50 mmol·L-1)中具有抑菌活性,且抑菌活性稳定.油菜、芥蓝、萝卜以及诸葛菜都属十字花科植物,说明十字花科多种植物种子中分离出的抗真菌蛋白具有一定的共性.
综上所述,Zp蛋白抑菌谱广,抑菌活性稳定,具有潜在的应用价值.但有关诸葛菜种子抗真菌蛋白抑制植物病原真菌生长的具体作用机理还有待研究.
猜你喜欢
河北北方学院学报(自然科学版)(2022年11期)2022-02-03
小读者(2021年4期)2021-06-11
中国科技纵横(2021年24期)2021-03-02
小哥白尼(军事科学)(2019年3期)2019-06-26
中成药(2018年6期)2018-07-11
小溪流(画刊)(2017年9期)2017-10-12
鸭绿江(2013年12期)2013-03-11
中国火炬(2011年7期)2011-07-25
