
围栏封育高寒草地植物地上生物量和物种多样性对关键调控因子的响应
2019-05-28 06:35陈智勇谢迎新
草业科学 2019年4期
陈智勇,谢迎新,刘 苗
(1.河南农业大学农学院,河南 郑州 450002;2.中国科学院地理科学与资源研究所,北京 100101)
过去几十年来,全球变化对地球生态系统产生强烈的影响作用[1]。其中以草地退化最为突出[2-3]。草地退化已成为危害人类社会、经济发展和居住环境重要的问题之一[4-9]。草地退化已经严重威胁到了区域经济和环境的可持续发展[10],并引起了许多国家政府及一些国际组织的关注[11-13]。中国是受退化影响最深的国家之一,特别是在我国的北方[14-15]。目前,青藏高原退化严重,据不完全统计,青藏高原大约有0.45亿hm2的退化草地,占青藏高原草地面积的1/3[16],而且退化面积还在持续的增加[17]。如何抑制草地退化,已经成为我国亟待解决的问题。
围栏封育作为当前草地退化主要的植被恢复和重建的措施之一[18-20],是指使用围栏设施将草地围封一定的时间停止利用,使得草地植物休养生息,得以更新,提高草地植物的生产力[18, 21]。毋庸置疑,退化草地在排除人畜干扰后必定会进行自我恢复,但是在草地自我恢复过程中,草地的群落结构和功能及土壤性质如何变化,是否围栏封育越长,草地植被群落和土壤养分恢复的越好,还是围栏封育也有一定的最佳期限,长期围栏会导致草地植被群落和土壤养分退化降低等,问题尚未探明。
为了回答上述问题,国内外学者就围栏对草原植物的产生的影响做了大量的研究。如Osem等[22]研究了围栏对以色列地中海干旱草地生物量的影响,Altesor等[23]分析围栏对乌拉圭草地植物群落结构和草地生产力影响,认为随着围栏年限的增加植物群落趋于稳定,生产力会显著提高。我国学者对围栏在植被群落动态、多样性和土壤理化性质等方面的影响进行了大量的研究[24-25],围栏显著提高了高寒草地植物的高度、盖度和地上生物量以及可食牧草的比例[24];而羊草(Leymus chinensis)草原的生物量和多样性随着围栏年限的增加表现出先增加后减少的趋势[25],导致这种现象的主要原因是在围栏前期牲畜对植被的破坏被有效抑制,促进了退化地植被恢复,从而有利于土壤水分和养分增加[26-27];而长期封围导致优势种的频度增加,主要表现在资源利用和侵占上,其他物种的生长发育受到限制[28]。但是,由于草原类型复杂以及围栏封育时间和当地载畜量和放牧强度不同,研究结果有所差异[28-29]。特别是以往的研究侧重于土壤或植被的单个因素的变化,地上生物量和物种多样性变化机制以及围栏封育对植物群落结构和土壤理化性质的影响仍需进一步探究,尤其是不同围栏年限下退化草地恢复过程这些要素的变化特征尚不明确。
为此,本研究选取西藏申扎地区,选取不同围栏年限(4、5、7、8和9年)进行植被群落特征土壤有关特性取样分析研究,旨在分析群落结构和土壤性质的关系,进而明确影响植被动态的关键环境要素,以期为今后退化草地保护恢复提供参考。
1 材料与方法
1.1 研究区概况
研究区位于青藏高原那曲地区申扎县的中国科学院生态试验站 (图 1),30°45′-30°57′ N,88°42′-89°06′ E。申扎县位于西藏中部,位于那曲地区的西南部,海拔 4 750 m。年均温 < 0 ℃,7-8 月最高温10 ℃,最低温-40 ℃。气候以高原亚寒带干旱季风为主,降水集中在5-9月,年降水量约298.6 mm。区域环境表现为无霜期较短,干燥度1.5~6.0,年均风速 3.8 m·s-1,8 级以上大风高达 104.3 d,年均太阳辐射为2 915.5 h。该地区草原辽阔,是西藏传统牧区之一,主要以高寒草地为主,申扎站所在西藏中部区域高寒草地约占土地面积40%,湖泊河流约占23%。优势种为紫花针茅(Stipa purpurea)和苔草(Carex moorcrofii),伴生种为火绒草(Stellera chamaejasme),瑞香狼毒(Stellera chamaejasme)和小叶棘豆 (Oxytropis microphylla)等[30-31]。
图1 研究区位置Figure 1 Location of the study area
1.2 研究方法
1.2.1 样品采集
于2017年7-8月(植物高峰期),经实地调查分析,选取了围栏年限为4、5、7、8和9年的围栏梯度样地,每个围栏梯度样地设置3次重复,随机选择 3 个 50 cm × 50 cm 的样方区域,记录每个样方内物种的盖度、高度和植株数,采用刈割法将植被的地上部分从贴近地表处剪除来采集地上部分测定地上生物量,将不同物种分类装入信封编号带回实验室,在烘干箱中先通过105 ℃杀青1 h 处理,然后温度调至 80 ℃ 烘干 48 h,恒重后称量每个物种的重量并记录群落的地上生物量。此外,在每个样方区域内使用土钻在0-10、10-20和20-30 cm土层深度进行土壤样品采集,取样装入自封袋并编号,带回实验室进行土壤理化性质分析处理。同时使用土壤紧实度仪器、土壤温度仪和容重环刀对3层土壤的温度、紧实度和容重进行测定,每个样方内土壤进行3次重复。
1.2.2 土壤养分含量测定
将土壤样品的植物根系挑除,磨碎土样通过0.15 mm的筛,使用C/N元素分析仪测量样品的全氮 (soil total nitrogen, STN)、全碳 (soil total carbon, STC)含量,采用钼蓝比色法测定土样全磷(soil total phosphorus, STP)含量,使用Olsen法测定土样速效磷 (soil available phosphorus, SAP)含量[32],利用碱解扩散法测定土壤速效氮 (soil available nitrogen, SAN)含量,采用重铬酸钾容量法测定土壤有机质 (soil organic matter, SOM)含量。
1.3 数据分析方法
使用物种丰富度指数(Margalef指数,D)来计算植物群落的物种多样性。

式中:N为个体总数;S为生态系统中物种数目。
采用的Duncan方法检验均值差异的显著性(P<0.05),用最小显著差异法(LSD)来确定围栏年限梯度不同处理间的物种盖度、植株高度和地上生物量等指标的差异 (P< 0.05)。以上分析在 SPSS 24.0(SPSS, Chicago, USA)中计算。采 用 Microsoft Excel 2013对数据来进数据汇总计算,用Sigmaplot 10.0进行绘图。
2 结果与分析
2.1 围栏对植被群落及多样性的影响
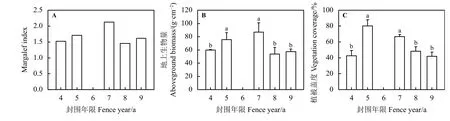
图2 不同围栏年限对群落多样性和植被群落的影响Figure 2 Effects of fencing years on the community biodiversity and vegetation community
随着围栏年限的增加,植物群落的多样性和地上生物量表现出相似的变化。随着围栏年限的增加,草地 Margalef指数 (图 2A)、地上生物量 (图 2B) 和植被盖度(图2C)均表现出先上升后逐渐下降的趋势;草地Margalef指数和地上生物量在围栏7年时达到最大值,分别为2.13和145 g·m-2。地上生物量在围栏5和7年之间以及围栏4、8和9年之间无显著差异(P> 0.05),5、7年的显著高于4、8和9年的(P< 0.05)。植被盖度在围栏4、8和9年之间无显著差异,但均显著低于围封5和7年。
2.2 不同围栏年限处理对土壤理化性质的影响
SAN含量随围栏年限增加表现出先上升后逐渐降低的趋势(图3A),在围栏4、8和9年之间无显著差异 (P> 0.05),但显著低于 5 和 7 年的 (P<0.05),且 7 年也显著高于 5 年 (P< 0.05)。STN (图 3B)和土壤有机质(SOM) (图3C)含量呈先缓慢上升后下降的趋势,且在不同围栏年限之间均表现出显著差异(P< 0.05)。STC含量随围栏年限增长逐渐下降(图3D),围栏4和7年之间以及围栏5、8和9年之间无显著差异(P> 0.05),但围封4和7年显著高于围封5、8和9年的(P< 0.05)。STP含量呈现缓慢变化,趋势不明显(图3E),围栏年限之间无显著差异(P> 0.05)。SAP含量随围栏年限增加而降低(图3F),围栏4和5年之间以及7、8和9年之间无显著差异(P> 0.05),但围封4年显著高于7、8和9年、围封5年显著高于围封8年的(P> 0.05)。
随着围封年限的增加,土壤含水量(soil water content, SWC)呈先增加后减小的趋势(图4A),围栏7年显著高于围封4年(P< 0.05),围封5、8和9年之间无显著差异 (P> 0.05);土壤紧实度 (soil density,SD)随围栏年限增加先升高后逐渐下降(图4B),围封4年和7年差异不显著(P> 0.05),围栏5、8和9年之间差异显著 (P> 0.05)。土壤温度 (soil temperature,ST)、土壤容重 (soil bulk density, SBD)和 pH 随围栏年限增加先下降后逐渐上升(图4C、D、E),ST围栏7年和8年之间无显著差异(P> 0.05),但与4、5和9年之间差异显著(P< 0.05)。SBD在围封7年时显著低于其余围封年限(P< 0.05)。土壤pH在围栏4年时显著高于围封 7 年 (P< 0.05),围封 5、8 和 9年之间无显著差异 (P> 0.05)。
2.3 围栏退化草地生物量对土壤理化性质的响应
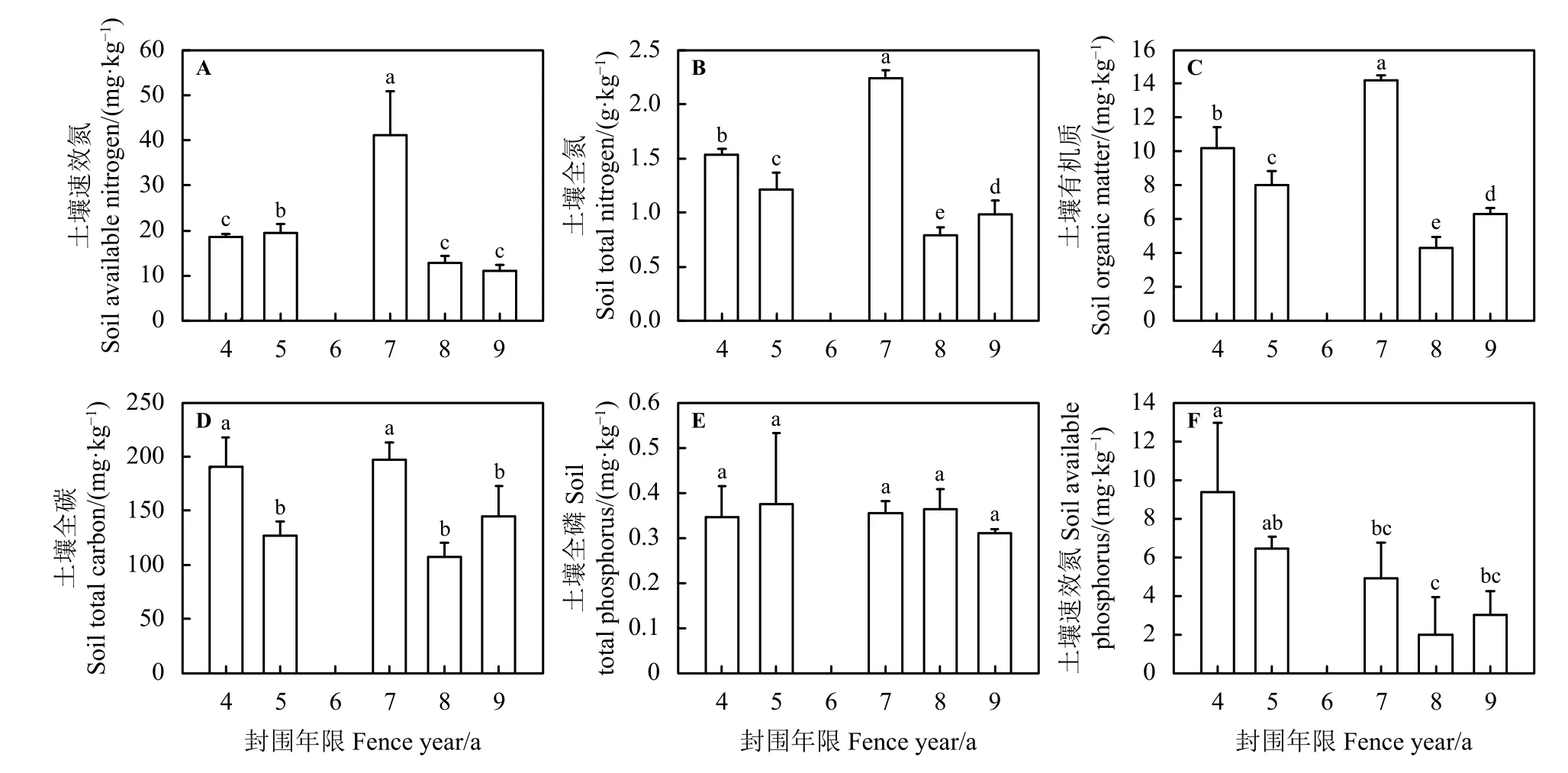
图3 不同围栏年限处理下土壤化学性质变化特征Figure 3 Characteristics of the soil chemical properties under different years of fencing treatment
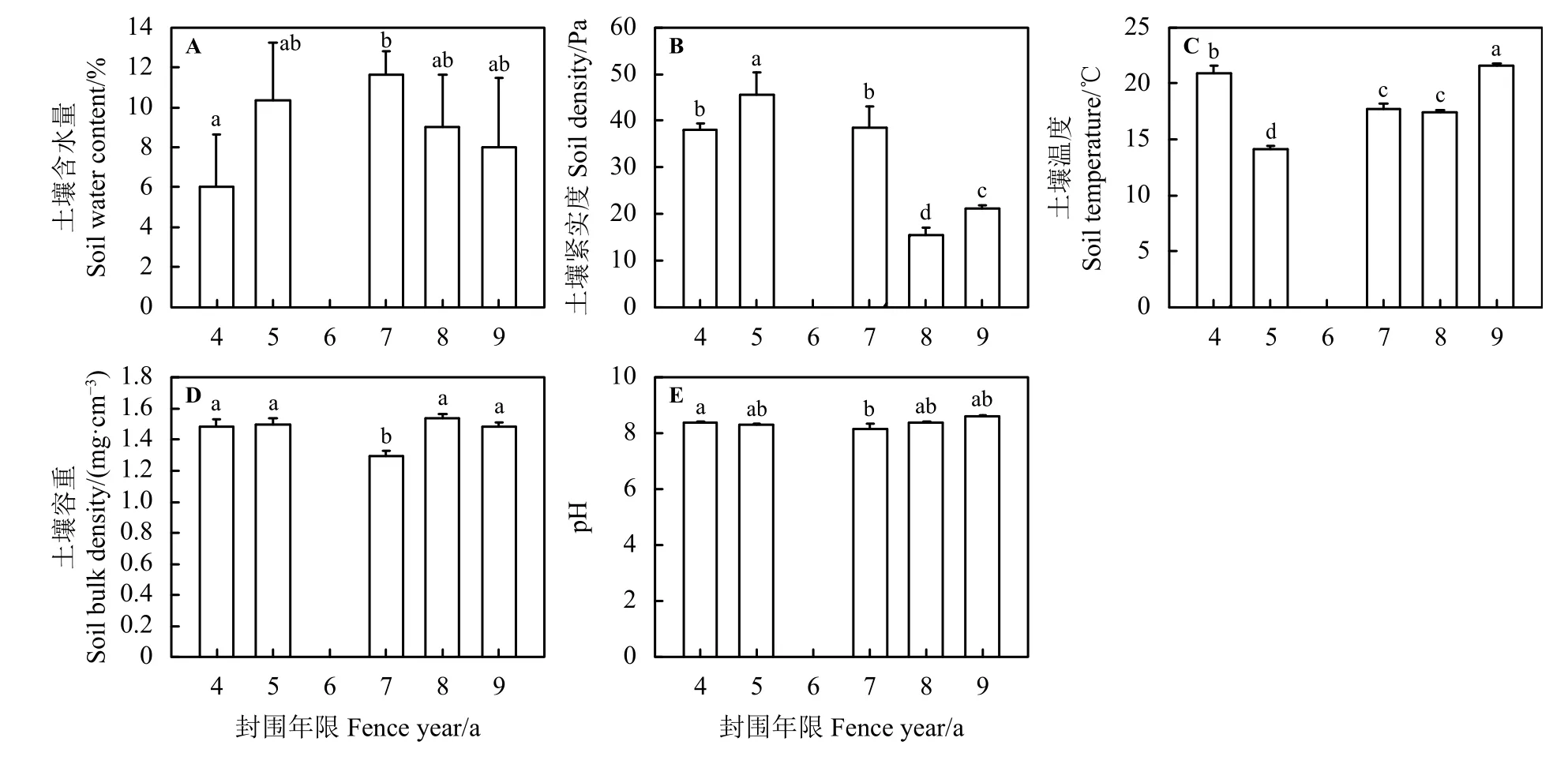
图4 不同围栏年限处理下土壤物理性质变化特征Figure 4 Characteristics of the soil physical properties under different years of fencing treatment
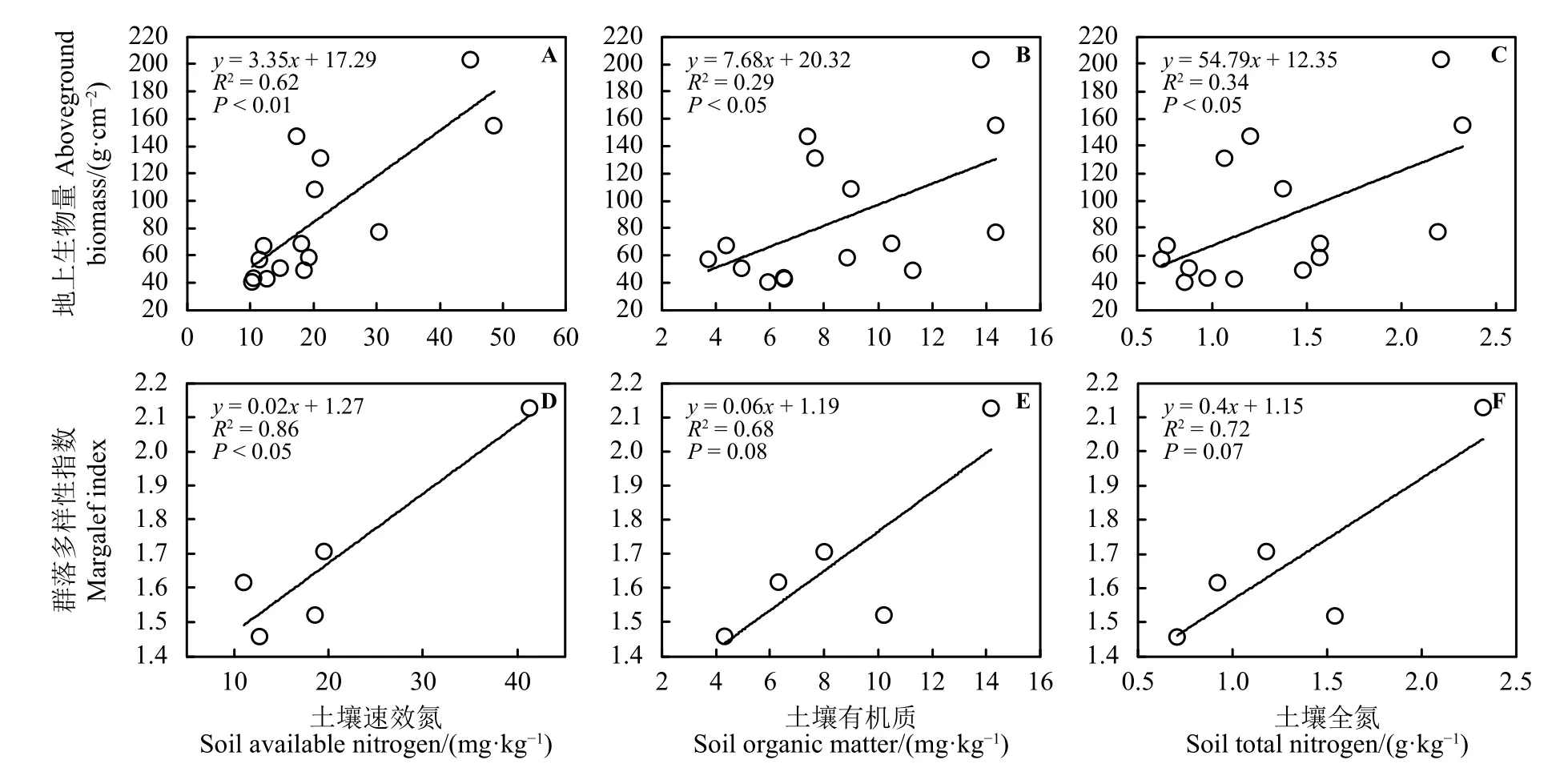
图5 围栏处理下地上生物量和群落多样性指数对土壤化学性质变化的响应Figure 5 Responses of the aboveground biomass and community biodiversity to changes in the soil chemical properties under enclosure treatment
通过土壤化学和物理性质的测定筛选出随着围栏年限土壤理化性质与植物生物多样性、地上生物量变化相似或相异的因子,并以线性相关性分析探究草地地上生物量和生物多样性的调控因子。结果表明,SAN、STN和SOM含量与围栏草地地上生物量存在显著正相关关系,草地地上生物量随SAN、STN和SOM的增加呈逐渐增加的趋势;其中SAN对围栏草地地上生物量的影响极显著(R2= 0.62,P< 0.01) (图 5A);STN (R2= 0.29,P<0.05) (图 5B)和 SOM (R2= 0.34,P< 0.05) (图 5C)对围栏草地地上生物量的影响显著。SAN、STN和SOM含量与围栏草地生物多样性存在正相关关系,生物多样性随SAN、STN和SOM的增加呈逐渐增加的趋势;其中SAN对围栏草地群落多样性的影响显著 (R2= 0.86,P< 0.05) (图 5D);而 STN (R2= 0.72,P= 0.07) (图 5B)和 SOM (R2= 0.68,P= 0.08) (图 5C)对草地生物多样性无显著影响。
同时SWC与地上生物量呈显著正相关关系 (R2=0.32,P< 0.05) (图 6B)。SWC 与物种多样性呈正相关关系 (R2= 0.55,P= 0.13) (图 6F),而 pH、STP 和SBD与地上生物量和物种多样性都呈负相关关系。
3 讨论
物种的多样性在时间动态变化上反映着植物演替的进程特点,目前围栏是使用最广泛的草地退化恢复的措施,并且能反映多样性动态[33]。草地生态系统的草地多样性越高,其系统越稳定,表示草地也就越健康[34]。本研究表明,围栏年限增加的同时,Margalef指数以及植物的群落盖度和地上生物量随围栏年限的增加呈先增长后降低的趋势(图2),以7年最高,与前人研究结果基本一致[25,35-36]。围栏封育可以解除放牧的压力,消除了人类对草地的干扰,为草地提供了恢复的机会,为植物生长发育创造了良好的条件,进而提高植被多样性、盖度及地上生物量[31, 37-38]。具体发生机制是围栏导致草地地上植物的枯落物增多,枯落物经过自然物理过程粉碎和微生物生化过程分解后再一次的进入土壤中,进而提升了土壤的有机质和养分的输入,进一步促进植株根际微生物活性的提高,促进植物生长[26],在围栏7年时生物量和物种多样性达到最大值,分别为 86.86 g·m-2(图 2B)和2.12(图2A)。另外植被对降尘和风蚀细粒物质具有截存效应,也有利于有机质和全氮的积累。从群落稳定性角度探讨,种间竞争是调节植物多样性的一个关键因素[39-40],随着围栏年限的增长群落中的优势种对资源的占用抑制了生命力弱的植物的生长发育,因此由于缺乏光照和营养物质,竞争能力较弱的植株密度降低,甚至从植物群落中消失,不利于草地的繁殖更新[41-43]。相关研究也表明,适度的围栏时间能够提升植被盖度和群落多样性,而长时间的围栏会导致生物多样性减少[44]。因此,在藏北退化地,围栏对植被恢复起到了先促进后抑制的作用,研究认为最佳封育期为7年。
土壤水分是干旱和半干旱区域植物生长发育的主要限制因子[45],本研究表明,水分和生物量呈显著正相关关系(图6B)。且相对围栏8和9年,围栏4、5和7年的草地土壤含水量较高,并随着围栏年限的增加先升高后降低,在7年达到最大值11.67%(图4A),这和一些研究结果相似[40]。分析其原因,在围栏7年之前是因为围栏前期牲畜的踩踏减少了,使紧实度降低(图4B),围封后植物根系得以生长,土壤结构得到恢复,从而增强了雨水渗透能力,使得土壤水分增加[46]。在围栏达7年之后,群落的地表破坏相对降低,土壤或者生物出现结皮,限制了雨水的渗入,导致降低了土壤含水量[47-48]。通常来说,水分的变化使得生物量改变,进而也会引起土壤温度、容重及pH的变化,本研究中,土壤温度、容重和pH随围栏年限的增加呈先升后下降的趋势(图4C、 D、E)。这与一些研究结果相似[49],分析其原因,短期禁牧后,群落盖度增加,使其地表蒸散降低,土壤温度降低。围栏后期植被盖度减少,土壤温度增加,反之温度增加导致地上生物量及植物多样性的减少(图6C、G);同时禁牧消除了牲畜的踩踏的影响,土壤孔隙度增加,加上根系的穿插,使得容重降低[50]。土壤pH对土壤的其他性质有着广泛的影响,但其在各个围栏年限之间变化较为缓慢,变化不显著,在8.13~8.58波动。这与一些研究结果相似[51]。
以往研究表明,放牧强度对土壤养分有很大影响,SAN、STN和SOM等养分含量随着放牧强度的加大而降低[52-53],而短期的围栏封育能够使SAN、STN和SOM含量增加[36]。本研究中,围栏使得全氮、速效氮和有机质呈先升高后降低的趋势,并在围栏 7年达到最大值,分别为22.40 g·kg-1、41.25 mg·kg-1和 14.18 mg·kg-1,但变化不显著 (P> 0.05)(图 3A、B、C),这与鱼小军等[36]的研究结果基本一致。原因是草地短期封育后,植物地上生物量增加,土壤能够储存大量地上部分凋落物,使得SOM含量增加,SAN和STN也会有所提高。有机质是土壤的重要组成部分,是植物所需的各种养分的基础,且含有有机态的氮,经过化学转化为植物所需的养分[54],使养分含量增加。退化草地随围栏封育年限增加植物群落初期快速发育,后期群落逐渐稳定,植被对土壤的消耗大于供给,导致土壤养分有所降低[55]。该研究结果与其他围栏封育研究结果相似[56-57]。而土壤全碳、全磷和速效磷含量无明显变化。这可能因为在高寒区,低温限制了营养的周转,导致土壤营养对围栏封育的响应缓慢相关[58]。
4 结论
围栏前期增加了多样性指数、地上生物量和盖度,但这些指标都表现出单驼峰状模式,阈值为7年,围栏年限大于7年会导致植物多样性和密度降低。因此,在围栏7年后,在植物休眠期进行割草或适度放牧,不会给草地造成损害,反而能改善草地质量,使得牧草得以更新。植物生物量和土壤是相互作用的关系,围栏处理不仅使地上生物量得以恢复,而且使得土壤养分增加,土壤养分又会反作用到植物,为植物提供生长所需营养。此外,过高的土壤pH、土壤温度和土壤容重是地上生物量下降的关键决定因素,而土壤含水量、土壤全氮、土壤速效氮和土壤有机质的增加是地上生物量增加的关键因素。
猜你喜欢
天津农林科技(2022年2期)2022-04-19
河南畜牧兽医(2022年3期)2022-04-13
今日农业(2021年2期)2021-11-27
绿色科技(2021年10期)2021-06-23
黑龙江工程学院学报(2020年5期)2020-10-21
红蜻蜓·低年级(2020年12期)2020-06-20
水土保持研究(2019年6期)2019-10-19
好孩子画报(2019年8期)2019-09-19
草业学报(2019年2期)2019-02-25
发明与创新·大科技(2018年2期)2018-03-17
