
绵羊HADHB 基因生物信息学分析及营养应激状态下的表达变化
2019-05-27 03:00郭丽荣韩高链张枫惠李俊玲
中国畜牧杂志 2019年5期
郭丽荣,韩高链,张枫惠,李俊玲,秦 健,杜 荣*
(1.山西农业大学动物科技学院,山西太谷030801;2.山西农业大学实验教学中心,山西太谷030801;3.山西农业大学生命科学学院,山西太谷 030801)
应激是指机体对内外界各种异常刺激所产生的非特异性应答反应的总和[1]。营养应激是一种重要的应激,营养素浓度过高或过低均可诱发营养应激[2]。而禁食会使动物处于严重的营养应激状态,机体代谢紊乱后必须动员脂肪储备的能量以保证生命活动的进行[3]。脂肪酸从脂肪组织中释放出来被转移到骨骼肌、心脏以及肝脏的线粒体进行氧化分解,产生ATP 供能[4]。有研究通过基因芯片技术对脂肪组织的功能基因进行筛选后初步明确HADHB(Hydroxyacyl-CoA Dehydrogenase/3-ketoacyl-CoA Thiolase/enoyl-CoA Hydratase,beta subunit)基 因 是影响脂类代谢的关键基因之一[5]。HADHB 参与脂肪酸的β 氧化,催化其反应的第2 至第4 步,即2- 烯酰辅酶 A 的水合、脂肪酸-2-羟基脂酰辅酶 A 的脱氢以及 β-酮脂酰辅酶 A 的硫解[6-7]。关于不同动物HADHB 基因的研究有所报道,例如该基因缺陷的小鼠肝脏会发生脂肪变性[8];在被喂食高脂食物的成年雌性斑马鱼体内,HADHB 基因mRNA 表达量上升[9];HADHB 基因在鸡的胚胎时期即有表达,且在胚胎发育后期表达量最高[10]。但关于绵羊HADHB 基因的研究目前尚未见报道。本实验对绵羊HADHB 基因序列进行一系列生物信息分析,预测该基因的理化性质、序列同源性、空间结构以及亚细胞定位等特性,并通过禁食构建营养缺陷应激模型,利用qRT-PCR 技术初步研究HADHB 基因的表达变化,为进一步深入研究奠定基础。
1 材料与方法
1.1 实验动物及样品采集 随机选取6 只体重、月龄相近的雌性成年健康绵羊,分为正常组和禁食组,每组3只。正常组按照常规的饲养方式饲养,禁食组禁食3 d,禁食期间给予饮水,禁食结束后正常屠宰,并迅速采取6 份同一部位的骨骼肌组织样本,置于液氮中保存备用。
1.2 主要仪器 超净工作台由苏州安泰空气技术有限公司生产;全自动凝胶成像分析系统、PCR 仪、荧光定量PCR 仪均由美国BIO-RAD 公司生产;电子天平由奥豪斯国际贸易(上海)有限公司生产;核酸蛋白浓度测定仪由美国Thermo 公司生产;电泳仪电源及电泳槽由北京六一生物科技有限公司生产。
1.3 生物信息学分析 根据GenBank 预测的绵羊HADHB 基因序列(XM_004005732.3)进行生物信息学分析。
①利用ProtParam(http://expasy.org/tools/protparam.html)在线软件分析蛋白质理化性质;②利用ProtScale(http://expasy.org/tools/protscale.html)在线软件分析蛋白质的亲水性与疏水性;③利用TMpred(http://www.ch.embnet.org/software/TMPRED_form.html)在线软件分析蛋白质的跨膜区;④利用EditSeq 和MegAlign 软件进行同源性分析和系统进化树分析;⑤利用SOPMA(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html)在线软件进行蛋白质二级结构预测;⑥利用SWISS-MODEL(https://www.swissmodel.expasy.org/)在线软件进行蛋白质三级结构预测;⑦利用PDBsum Generate(http://www.ebi.ac.uk/thornton-srv/databases/pdbsum/Generate.html)在线软件评估预测的蛋白质三级结构;⑧利用PSORT II(https://www.genscript.com/psort.html?src=leftbar)在线软件进行亚细胞定位分析。
1.4 总RNA 提取及cDNA 合成 首先,将绵羊骨骼肌样本放入加有液氮的研钵中充分研磨后加入RNAios plus(TaKaRa)提取总RNA。取少量溶解的RNA 进行琼脂糖凝胶电泳,检测RNA 是否降解。其次,用核酸蛋白浓度测定仪测定RNA 浓度。最后,利用PrimeScripTMRT reagent Kit(TaKaRa)将RNA 反转录为cDNA,反应体系为10 μL: PrimerScriptRTMaster Mix(5×)2 μL,RNA 4 μL,RNase Free water 4 μL,混匀后于PCR 仪中反转录,反应条件为37℃ 15 min,85℃ 5 s,4℃ 5 min。终止反应后对反转录产物进行浓度测定。
1.5 引物设计及合成 选取常用的内参基因ACTB(HM067830.1)和GAPDH(HM043737.1),在NCBI设计引物并检测特异性,目的基因引物设计及合成方法同内参基因。引物序列由北京六合华大基因科技有限公司合成(表1)。

表1 基因PCR 引物序列信息
1.6 荧光定量PCR 实时荧光定量PCR 体系为20 μL:SYBR Premix Ex TaqTMII(2×)(TaKaRa)10 μL, Forward Primer (10 μmol/L)0.8 μL,Reverse Primer (10 μmol/L)0.8 μL,cDNA (100 ng/μL)2 μL,ddH2O 补足体系至20 μL。qRT-PCR 反应条件为95℃2 min,95℃30 s,60℃ 34 s。
1.7 统计分析 利用NormFinder 软件对2 个候选内参基因进行稳定性评估:分别确定这2 个基因6 个样本荧光定量PCR 得出的CT 值中的最小值,用其余5 个值分别减去最小值算出ΔCT,并进一步计算出2-ΔCT,再用NormFinder 计算两者的稳定值M。
目的基因的数据处理:根据2-ΔΔCT计算目的基因mRNA 的相对表达量,并利用SPSS 21.0 进行数据统计和差异显著性分析,P<0.001 表示差异极显著。用GraphPad PrismTM做柱状图。
2 结果与分析
2.1 绵羊HADHB 基因生物信息学分析
2.1.1 绵羊HADHB 蛋白理化性质分析 根据ProtParam在线软件分析结果得出绵羊HADHB 基因编码的氨基酸数目为475 个,其中含量最多的是Ala(A),占比12.2%,其次是Arg(R),占比4.8%,不含Pyl(O)和Sec(U)。负电荷残基(Asp+Glu)数是42,正电荷残基(Arg+Lys)数是55。理论等电点(pI)为9.38,表明HADHB 蛋白为碱性蛋白,其平均疏水性为-0.093,分子式是C2273H3648N630O673S24,分子量是51.3 ku,不稳定指数是34.16,较为稳定。在体外哺乳动物网状细胞中预测HADHB 蛋白半衰期为30 h,在酵母体内半衰期大于20 h,在大肠杆菌体内为大于10 h。
2.1.2 绵羊HADHB 蛋白亲水性与疏水性分析 ProtScale对绵羊HADHB 蛋白的亲水性和疏水性分析结果显示,在氨基酸第242 位的谷氨酰胺亲水性最强,数值是-2.478,第319 位的缬氨酸疏水性最强,数值是2.067(图1)。
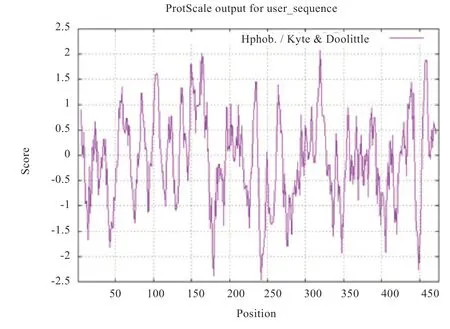
图1 绵羊HADHB 蛋白的亲水性
2.1.3 绵羊HADHB 蛋白跨膜区分析 利用TMpred 分析蛋白质的跨膜区结果显示,从内到外有4 个跨膜区,分别是138~164、301~322、425~442、450~470 氨基酸位点。从外到内有5 个跨膜区,分别是450~469、421~442、309~332、295~319、151~176 氨基酸位点(图2)。
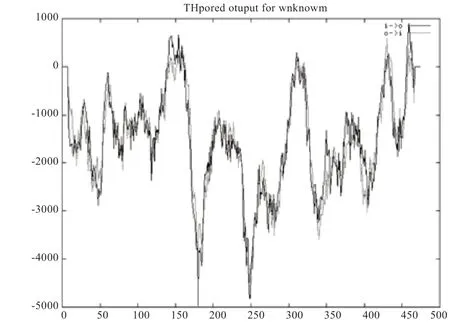
图2 绵羊HADHB 蛋白跨膜结构预测
2.1.4 绵羊HADHB 氨基酸序列同源性及系统进化树分析 利用EditSeq 和MegAlign 进行HADHB 氨基酸序列同源性分析和系统进化树分析,序列同源性分析结果显示(图3),绵羊HADHB 基因编码的氨基酸序列与牛(XM_005212929.3)、 山 羊(XM_013967822.2)、 人(XM_024452830.1)、卡氏小鼠(XM_021163089.1)、藏羚羊(XM_005978913.1)、东北狒狒(XM_021924673.1)、非洲鸵鸟南非亚种(XM_009669656.1)相应序列的同源性分别为96.4%、98.5%、90.7%、90.3%、99.6%、90.7%、82.7%,表明该基因在不同物种中保守性较高。系统进化树分析显示,绵羊和山羊、藏羚羊、牛的亲缘关系较近,与藏羚羊亲缘关系最近,与生物分类学保持一致(图4)。

图3 绵羊和其他物种HADHB 氨基酸序列同源性分析

图4 绵羊HADHB 氨基酸序列系统进化树分析
2.1.5 绵羊HADHB 蛋白二级结构预测 利用SOPMA在线软件预测HADHB 蛋白二级结构,结果显示二级结构中α-螺旋、β-转角、无规则卷曲和延伸链分别占43.58%、6.95%、34.32%、15.16%,α- 螺旋和无规则卷曲占大多数(图5)。

图5 绵羊HADHB 蛋白二级结构预测
2.1.6 绵羊HADHB 蛋白三级结构预测 通过SWISSMODEL 软件分别预测绵羊、山羊、藏羚羊、牛HADHB 蛋白的三级结构的结果显示(图6),该蛋白在4 个物种中均呈四聚体结构,且较为相似,与序列同源性分析及系统进化树分析中绵羊、山羊、藏羚羊、牛亲缘关系相近的结果保持一致。存在大量的α-螺旋和无规则卷曲,与绵羊HADHB 蛋白二级结构预测结果保持一致。
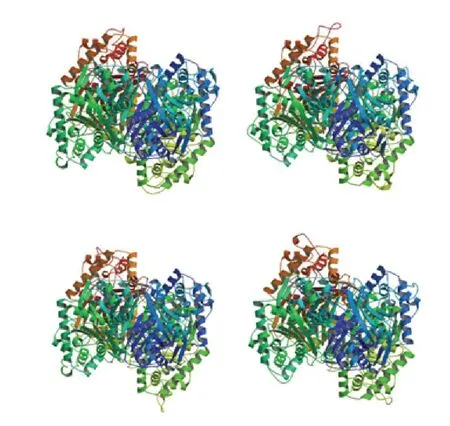
图6 绵羊、山羊、藏羚羊和牛的HADHB 蛋白三级结构预测
2.1.7 评估预测的绵羊HADHB 蛋白三级结构 利用PDBsum Generate 评估预测的绵羊HADHB 蛋白三级结构的结果显示,红色、棕色以及深黄色区域三者作为圆点允许出现的区域,所占比例分别是85.3%、12.1%、2.0%,表明该三级结构预测结果良好,可信度高(图7)。

图7 评估预测的绵羊HADHB 三级结构
2.1.8 绵羊HADHB 基因亚细胞定位分析 PSORT II 在线软件对绵羊HADHB 的亚细胞定位结果显示,有91.3%位于线粒体,有4.3%位于细胞核,还有4.3%位于细胞质。
2.2 荧光定量分析结果
2.2.1 内参基因的确定 候选内参基因ACTB 和GAPDH稳定性评估结果中,两者的稳定值M 分别是0.066 和0.302,M 值越小代表稳定性越好,所以选择ACTB 为内参基因。
在基因表达分析中内参基因的选择是十分必要的[11]。本实验通过对2 个内参基因稳定性的评估,证明ACTB基因在绵羊骨骼肌中的表达具有更高的稳定性,更适合作为该组织基因表达的内参基因。
2.2.2 营养应激状态下HADHB 基因表达的变化 qRTPCR 结果显示,ACTB 和HADHB 基因扩增动力学曲线和熔解曲线均较为平整,拐点清晰,结果良好(图8)。禁食组绵羊HADHB 基因mRNA 相对表达量与正常组相比极显著升高了8.9 倍(P<0.001),说明营养应激状态下HADHB 基因的表达上调(图9)。

图8 绵羊ACTB、HADHB 基因qRT-PCR扩增动力学曲线和熔解曲线
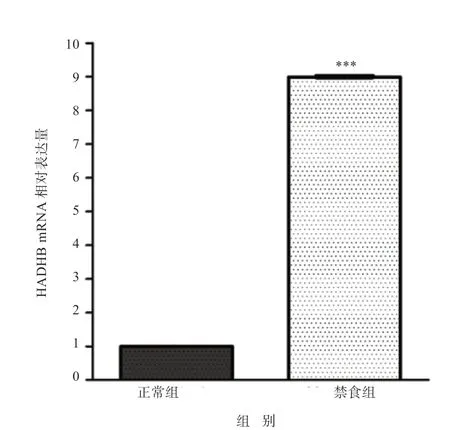
图9 正常组和禁食组绵羊HADHB 基因mRNA 的表达
3 讨 论
HADHB 是脂肪酸β 氧化的重要成员之一,其表达被靶向抑制后脂肪酸β 氧化也会受到抑制[12]。低乳脂率的奶牛乳腺上皮细胞中HADHB 基因高表达[13]。HADHB 基因编码4 个β 亚基,与HADHA 基因编码的4 个α 亚基共同构成多酶复合体- 线粒体三功能蛋白(Mitochondrial Trifunctional Protein,MTP)[14]。这2 种亚基的基因突变均有可能导致MTP 活性降低和正常功能的破坏[15]。HADHB 基因缺陷会导致机体线粒体脂肪酸氧化障碍(Fatty Acid Oxidation Disorder,FAOD),临床表现为肌张力低下,低血糖甚至发生脑病[16-17]。
本研究经生物信息学分析得出,绵羊HADHB 蛋白由475 个氨基酸构成,等电点为9.38,属于碱性蛋白,为日后蛋白质的分离、盐析和变性复性提供了理论依据。HADHB 蛋白不稳定指数是34.16,在哺乳动物、酵母、大肠杆菌中均具有较长的半衰期。有研究认为,蛋白质的半衰期越长其稳定性越高[18],HADHB 蛋白较长的半衰期也在一定程度上反映其较稳定的特性。据此可推断,HADHB 基因可在真核和原核细胞中重组,有利于过表达载体的构建和表达[19]。蛋白质的结构和功能与其跨膜区有着密切联系,根据跨膜区预测结果可推断配体特异性结合的位点,为HADHB 特异性受体及受体阻断剂的研究提供了依据。进化树结果表明,绵羊HADHB 氨基酸序列与牛、山羊、人、卡氏小鼠、藏羚羊、东北狒狒相应序列的同源性均超过90%,说明该基因编码区在生物长期进化中具有较高的保守性。其中绵羊和山羊、藏羚羊、牛亲缘关系较近,可为未来研究HADHB 在绵羊营养应激中的作用提供比较对象。蛋白质亚细胞定位显示该蛋白有91.3%定位于线粒体,与脂肪酸β 氧化的场所一致[20];同时实时荧光定量PCR 结果显示,禁食组绵羊HADHB 基因表达量极显著升高,因此推断机体在营养缺陷应激状态下,会刺激HADHB 基因表达增加,促进脂肪酸发生β 氧化,进而产生ATP 维持生命活动。根据绵羊三级结构预测结果得出该蛋白是一个四聚体,这种结构是否对脂肪酸的β 氧化有影响还有待进一步研究。
4 结 论
本研究首次通过生物信息学分析对绵羊HADHB蛋白质的理化性质和生物学特性进行了预测,通过实验比较确定ACTB 作为内参基因,检测到营养缺陷应激下HADHB 基因mRNA 表达量显著上升,为进一步深入研究绵羊HADHB 基因在营养应激中的作用及其机制奠定了基础。
猜你喜欢
传媒评论(2022年10期)2022-12-26
天津市教科院学报(2021年5期)2021-11-10
现代临床医学(2021年2期)2021-03-29
科学(2020年2期)2020-08-24
科学(2020年2期)2020-01-08
传媒评论(2019年12期)2019-08-24
传媒评论(2019年10期)2019-06-05
中国医学创新(2017年7期)2017-03-31
江苏农业科学(2016年8期)2017-02-15
烹调知识(2015年7期)2015-11-23
