
中国黄牛酪氨酸关联蛋白1(TYRP1)基因多态性的研究
2019-05-27 11:21姚一博党瑞华雷初朝
中国牛业科学 2019年2期
荣 誉, 姚一博, 党瑞华, 陈 宏, 雷初朝
(西北农林科技大学动物科技学院,陕西 杨凌 712100)
毛色不仅是牛一种明显的外貌特征,也是一种质量性状。毛色具有多样性,观察简单的特点,因此可以用毛色作为品种划分的依据,此方法具有简单、快捷、成本低的优点,这一点在其他动物中也有应用[1-3]。由于牛个体经济价值大,牛品种的划分与育种工作应该在早期进行,因此毛色便是一个理想的遗传标记;其次,牛的毛色性状还可能与生产性能有关[4],所以研究中国黄牛中与毛色有关的基因具有重要的意义。
牛的毛色与皮肤、毛发中黑色素与伪黑色素的相对数量和分布情况有关。黑色素广泛存在于各种动植物中,并且黑色素是脊椎动物表层结构的色素中含量最高的色素。黑色素可分为真黑色素和伪黑色素。其中,真黑色素为黑色或褐色。伪黑色素则大多为黄色和红色,颜色相对较浅[5]。黑色素是由黑色素细胞产生的黑色素体经过一系列酶学反应生成的。而黑色素体是在黑色素细胞内的高尔基体—内质网—溶酶体复合物(GERL complex)内形成,本身并无活性,无法产生黑色素[6]。黑色素是由酪氨酸激酶(tyrosinase,TYR)催化酪氨酸羟化而启动的一系列生化反应而合成的。
酪氨酸相关蛋白酶1(tyrosinase-related protein 1,TYRP1)基因对真黑色素的形成具有重要作用[7-8]。有研究表明,TYRP1基因可以影响黑色素的合成[9],且在控制黑色素细胞产生黑色素类型的关键步骤中起催化作用[10]。由于不同物种间以及同一物种内不同个体间均有相似的毛色类型,因此认为不同动物中都存在关于毛色变异的遗传基础[6]。目前TYRP1基因影响毛色的具体机制尚不清楚,但是研究TYRP1基因的多态性对培育特定颜色的品种仍具有重要意义。Berryere等[9]研究发现TYRP1基因的突变(g.1300C>T)与徳科特牛的棕色被毛性状有关,该突变导致组氨酸替换为酪氨酸,其中CC基因型被定义成BB,CT基因型被定义成Bb,TT基因型被定义成bb,bb基因型个体表现为棕色被毛性状。但该位点并没有在中国黄牛中进行过验证。本试验旨在利用测序等技术验证该突变是否与中国黄牛被毛颜色存在相关性,从而为中国黄牛的纯种鉴别、保种选育等提供理论依据。
1 材料与方法
1.1 试验动物
试验采集了1 143个牛耳组织样品,其中包括14个中国黄牛品种(991头)和48头安格斯牛、104头婆罗门牛。采用常规苯酚—氯仿法抽提基因组DNA,稀释浓度至10~20 ng/μL,保存备用。
1.2 引物合成和PCR扩增
选用Berryer等[9]报道的1个与徳克特牛的棕色毛色相关的SNP引物信息(表1),引物由上海生工生物工程股份有限公司合成。
25 μL PCR扩增体系:2×Mixture(上海宝生物公司)10.5 μL,上、下游引物(10 pmol/L)各0.5 μL,模板DNA 1 μL,超纯水12.5 μL。PCR反应程序:95 ℃预变性5 min,95 ℃变性30 s,64 ℃复性30 s,72 ℃延伸30 s,32个循环后,72 ℃终延伸10 min,放置于4 ℃条件下保存。利用1.0%琼脂糖凝胶电泳(含溴化乙啶染色)对PCR产物进行检测。
1.2.1 测序、基因型及基因频率的计算 将符合测序要求的PCR产物送至上海生工生物工程股份有限公司进行测序,测序结果用Chromas 2.3软件进行比对和校正。将所有黄牛的SNP位点进行统计,计算黄牛的基因型与等位基因频率。
1.2.2 对各群体进行Hardy-Weinberg平衡检验以及多态性分析 统计16个牛品种的基因型频率与基因频率,使用POPGENE软件检验种群是否达到Hardy-Weinberg平衡(Version 3.2),并计算多态性信息(PIC)含量[11]。

表1 引物信息
2 结果与分析
2.1 TYRP1基因的基因型频率和等位基因频率分析
本研究分析了TYRP1基因第7外显子上的突变(g.1300C>T)在991头中国黄牛(14个品种)、48头安格斯牛、104头婆罗门牛中的变异情况。从表2、表3可以看出,BB基因型仍然是优势基因型,在不同的群体中B等位基因频率远远高于b等位基因频率。同时,笔者还发现,红棕色和褐色群体的b等位基因频率高于其他群体,其中红棕色个体中,b等位基因频率为0.077,B等位的基因频率为0.923。褐色个体中,b等位基因频率为0.125,B等位基因频率为0.875。但是,本研究结果显示,此突变与不同毛色的牛并无相关性。
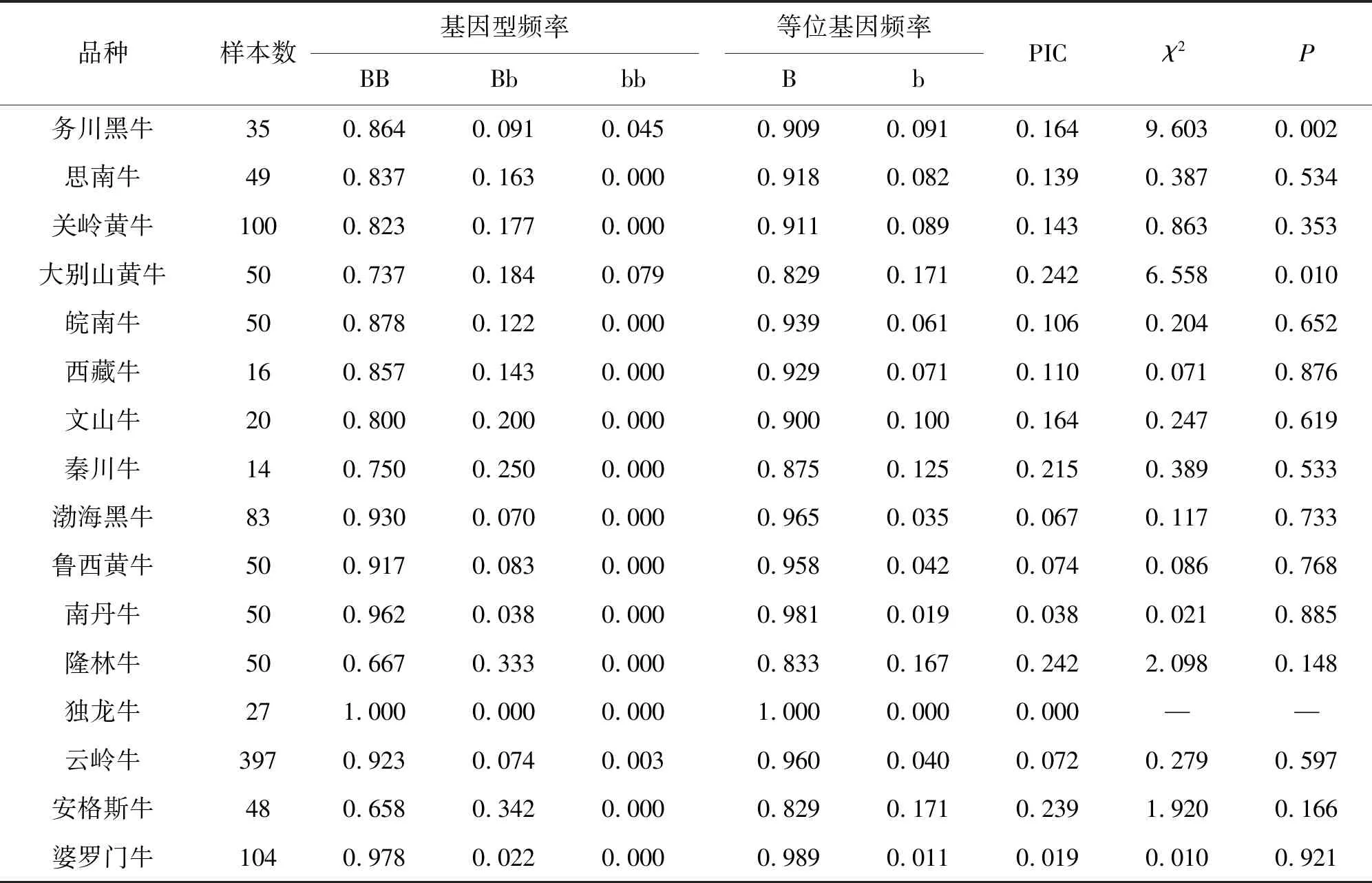
表2 黄牛TYRP1基因型频率、等位基因频率及多态性分析
2.2 TYRP1基因遗传多态性信息分析与Hardy-Weinberg平衡的检验
通过对16个品种进行多态性分析发现群体多态性均为低度多态(PIC<0.25)(表2)。Hardy-Weinberg平衡分析(表2)表明,该突变在思南牛、关岭黄牛、大别山牛、皖南牛、西藏牛、文山牛、安格斯牛、秦川牛、渤海黑牛、鲁西黄牛、南丹牛、隆林牛、独龙牛、云岭牛、婆罗门牛中达到Hardy-Weinberg平衡状态(P>0.05),而务川黑牛和大别山牛均未达到Hardy-Weinberg平衡状态(P<0.05)。
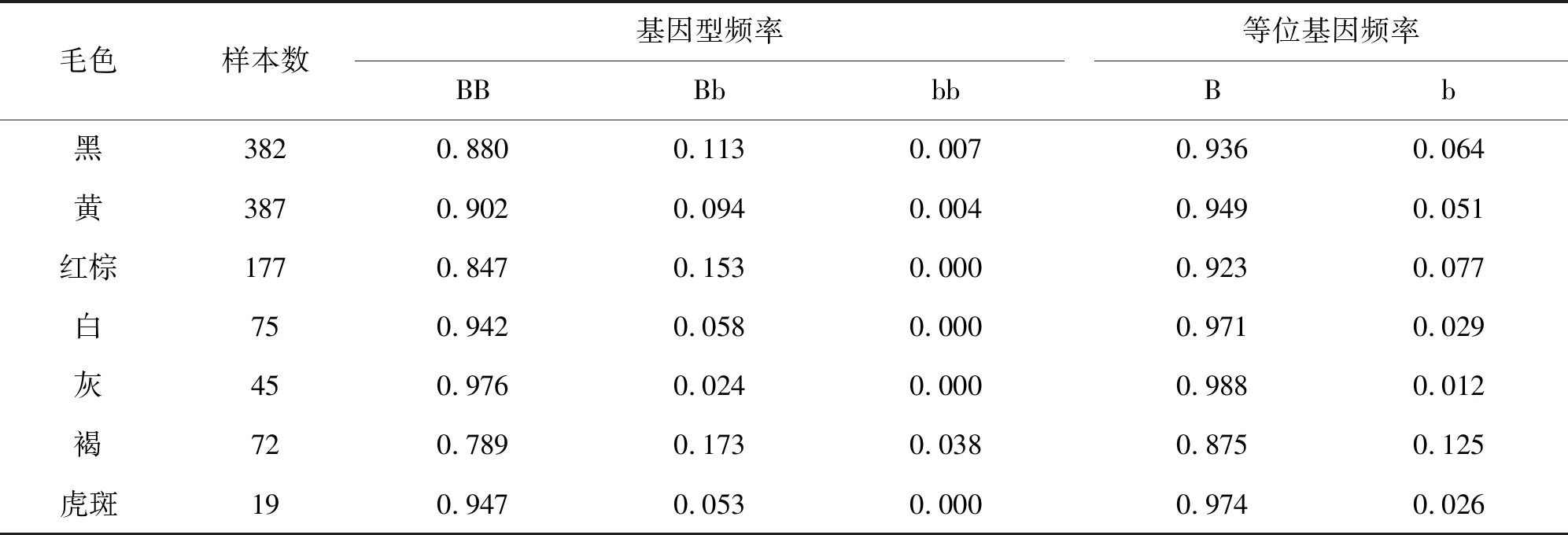
表3 不同毛色牛的基因型频率与等位基因频率
3 讨 论
在牛羊中等偶蹄动物中,对TYRP1基因多态性与毛色性状相关性的研究较多,大量研究表明,TYRP1基因与毛色性状存在相关性。Gratten等[12]发现索尔艾绵羊TYRP1基因外显子4上含有非同义突变(g.869A>G)与浅色表型呈现极显著的相关性,但在不同毛色个体表达量不同。在猪与鸟类中也有类似报道[13-14]。此外,在朗德鹅中通过定量PCR发现,TYRP1在不同羽色中呈现差异性表达,在颜色较深的个体中表达量更高[15]。Berryere等[9]发现,德克斯特牛TYRP1基因编码区的一个单核苷酸多态性(H424Y)与其棕色表型相关。
从表2可以看出,大别山牛、安格斯牛、秦川牛、隆林牛多态性较高,隆林牛的遗传多样性最高(0.242),这可能与当地开展的保种工作有关[16]。从群体遗传多态性角度分析,如果PIC低说明群体在该位点遗传变异低,有较低的选择余地,不可以利用该位点进行与毛色性状相关的标记辅助选择。群体PIC低还说明遗传变异小,可获得的遗传进展相对较少。因此,该位点不能作为中国黄牛毛色基因的辅助标记选择。
本研究结果显示,不同毛色的个体与TYRP1第7外显子上的突变(g.1300C>T)没有相关性。从表3中可以发现,b等位基因在不同毛色群体中均表现为低频,且与红棕色表型无相关性。同时,笔者发现b等位基因在深色表型群体中的频率高于浅毛色群体。Li等[17]研究发现,在不同颜色的蒙古马的皮肤组织细胞中TYRP1基因呈现差异性表达,板栗色蒙古马的TYRP1基因相对表达量最高。Makiko等[18]研究发现,在白化病人的皮肤组织中TYRP1基因的相对表达量显著低于正常个体。因此,笔者认为TYRP1基因第7外显子上的突变(g.1 300C>T)与红棕色毛色不相关,但可能对毛色具有一定的调控作用。
猜你喜欢
林业科技(2022年4期)2022-08-06
养猪(2021年4期)2021-08-26
阅读与作文(小学低年级版)(2019年12期)2019-12-26
奥秘(创新大赛)(2019年8期)2019-11-30
小天使·四年级语数英综合(2019年9期)2019-11-09
创新作文(3-4年级)(2019年3期)2019-09-03
小学阅读指南·低年级版(2017年10期)2017-11-20
创业家(2015年2期)2015-02-27
小学生作文·小学中高年级适用(2014年3期)2014-08-18
