
林木根际细菌JYZ-SD5的促生抗逆性能及种类鉴定
2019-05-17 09:36徐秀倩吴小芹吴天宇曾梦嫚
生物技术通报 2019年3期
徐秀倩 吴小芹 吴天宇 曾梦嫚
(1. 南京林业大学南方现代林业协同创新中心,南京林业大学林学院,南京 210037;2. 江苏省有害生物入侵预防与控制重点实验室,南京 210007)
土壤微生物群落丰富,各具有不同的生物学功能。植物根际促生菌(Plant growth promoting rhizobacteria,PGPR)生存于植物根际和根表[1],具有促进植物生长,拮抗病原微生物[2],提高植物抗性,修复土壤,维持土壤质量健康等功能[3-4]。如PGPR可在植物根际快速繁殖,固定大气中的氮素[5]、使植物充分利用土壤的养分,促进植物更好生长[6]。Liu等[7]从杨树根际筛选出一株荧光假单胞菌(Pseudomonads fluorescent)JW-JS1,可显著促进NL-895杨和美洲黑杨苗木的生长和营养元素含量的提高。宋芳旭等[8]发现根际细菌水拉恩氏菌(Rahnella aquatilis)JZ-GX1对杨树溃疡病菌金黄壳囊孢(Cytospora chrysosperma)的防效可达84%。部分芽孢杆菌属(BacillusCohn)还可提高植物对重金属的耐受性[9],Kuramshina 等[10]发现 用枯草芽孢杆菌(Bacillus subtilis)内生菌株处理Sinapis alba种子可提高植物对镉和镍毒性作用的抵抗力,并减少地上部分的伤害表现。
在自然条件下PGPR对植物的有益功能是多方面相互作用的结果,然而目前关于PGPR的研究大多侧重于某个或某些生物学功能,对PGPR综合性能的研究相对较少。本实验室前期从杨树根际分离出一株具有解有机磷和产ACC脱氨酶特性的细菌菌株JYZ-SD5,初步研究表明其对杨树具有一定的促生作用。但关于该菌株更多的生物学功能并不十分清楚,其分类地位也不明确。为深入探讨和弄清该菌的潜在特性和应用价值,本研究进一步对该林木根际细菌JYZ-SD5的促生抗逆性能进行检测,同时展开林木温室促生试验,并确定其分类地位,以期为评估该林木根际细菌JYZ-SD5在野外试验中的应用价值提供参考依据。
1 材料与方法
1.1 材料
1.1.1 供试菌株 供试根际细菌:实验室前期从山东杨树根际分离出的细菌JYZ-SD5。供试林木病原菌:水杉赤枯病菌:细极链格孢(Alternaria tenuissima)、异角状拟盘多毛孢(Pestalotiopsis heterocornis);茶树纹叶枯病菌:山茶球座菌(Guignardia camelliae);茶树轮斑病菌:异色状拟盘多毛孢(Pestalotiopsis vesicolor);松枯梢病菌:松杉球壳孢(Sphaeropsis sapinea);杨树溃疡病菌:七叶树壳梭孢(Fusicoccus aesculi);银杏叶枯病菌:链格孢(Alternariasp.)。所有菌株保存于南京林业大学森林病理学实验室。
1.1.2 植物材料 2年生水杉盆栽实生苗,基质选用南京林业大学北大山林木根际表层土。
1.1.3 供试培养基 牛肉膏蛋白胨(NA)培养基,蒙金娜培养基用于测定菌株溶解有机磷的能力[11],无氮培养基用于固氮能力定性测定[12],解钾能力检测培养基[13],纤维素刚果红培养基用于分解纤维素能力的测定[14]。蛋白质酶检测培养基[15],金氏培基B(King’S medium B,KB)用于菌株产IAA的测定[11]。
1.2 方法
1.2.1 根际细菌JYZ-SD5固氮、解磷、解钾性能测定 (1)固氮特性检测:将供试菌株采用平板划线的方法接种在无氮培养基中,每个处理3次重复,置于28℃低温恒温培养箱(MIR-553,Japan),连续观察细菌的生长状况。(2)解有机磷特性检测:将供试菌株采用三点接菌的方法接种在解有机磷培养基中,每个处理重复3次,置于28℃培养箱中,连续观察菌落周围透明圈的有无。(3)解钾能力测定:将供试菌株采用液体培养的方法,接种到解钾检测液体培养基中,每个处理重复3次,置于28℃摇床培养,转速为200 r/min,培养数天后,用火焰分光光度计(FP6410,上海)检测细菌的解钾能力(解P1为空白对照,P2为接菌处理)。
1.2.2 根际细菌JYZ-SD5产IAA能力检测 (1)定性检测:挑取单菌落接入金氏培基B中,恒温摇床(Multitron Standard,Switzerland)37℃ 培养 24 h,用无菌枪头吸取1 mL菌液于无菌离心管中,加入4 mL Sackowcki’s 显色剂之后迅速充分混合,室温下避光显色40 min,观察颜色变化。(2)定量检测:精确称取IAA10 mg,先用少量乙醇溶解,再用蒸馏水定容至100 mL(浓度为100 μg/mL),然后分别稀释成 0、4、8、12、16、20、24 μg/mL,取各浓度1 mL,分别加入4 mL显色剂,40℃暗保温40 min,与OD535下测定OD值,绘制标准曲线。将培养的菌悬浮液和空白对照,离心10 min(10 000 r/min)后取上清液4 mL加入等量比色液,在黑暗中静置40 min,取出立即用HexIOS分光光度计(Lamnda365,Korea)测定OD535值,每个样品重复测3次,以加入比色液的空白对照调零,对比标准曲线计算该菌株分泌IAA量,连续测定7 d。
1.2.3 根际细菌JYZ-SD5拮抗林木病原菌活性测定 (1)平板拮抗:供试病原真菌接种于PDA培25℃养基中,每皿接种一块,重复3次,用接种环沾取液体摇培24 h后的菌液,在菌落两侧对称划线,放入培养箱培养,连续观测记录菌株的抑菌效果。(2)胞外代谢产物拮抗活性:获取供试菌发酵液,发酵液移入灭菌50 mL离心管中,10 000 r/mim,4℃离心10 min,取上清液,过细菌滤膜,得无菌胞外代谢产物。将无菌胞外代谢产物按0.5%的比例加入PDA培养基中,摇匀倒平板。将林木病原菌接种于平板中,重复3次,置于25℃培养箱中培养,观察该菌抑菌效果(抑菌率=(对照组菌落直径-处理组菌落直径)/对照组菌落直径×100%)。
1.2.4 根际细菌JYZ-SD5产纤维素酶、蛋白酶测定 (1)纤维素酶检测:将供试菌株采用三点接菌的方法接种在纤维素酶检测培养基中,每个处理重复3次,置于28℃培养箱中,连续观察菌落周围透明圈的有无。(2)蛋白酶检测:将供试菌株采用三点接菌的方法接种在蛋白酶检测培养基中,每个处理重复3次,置于28℃培养箱中,连续观察菌落周围透明圈的有无。
1.2.5 根际细菌JYZ-SD5的重金属耐受性测定 (1)配制NA培养基,分别添加KMnO4、NiCl2.6H2O、K2Cr2O7、CuSO4.5H2O。 使 Mn7+、Ni2+、Cr6+、Cu2+浓度 梯 度 为 5、10、15、20、30、40、50、80、100、200、300、400 mg/L( 以 纯 Mn7+、Ni2+、Cr6+、Cu2+计),对照为纯培养基。将菌株接种于上述培养基中,放入培养箱中,24 h后进行观察。(2)不同重金属浓度胁迫下菌株JYZ-SD5生长曲线的测定:将菌株JYZ-SD5接种到不同重金属浓度的液体NB培养基中,用微生物全自动生长曲线分析仪(Bioscreen C,Finland),每隔2 h测定不同盐胁迫下的OD600值,并以时间为横坐标,OD600值为纵坐标绘制菌株JYZSD5的生长曲线。
1.2.6 根际细菌JYZ-SD5对水杉促生试验 用接种环挑取单菌落于含有50 mL NA液体培养基的100 mL三角瓶中,28℃、200 r/min振荡培养24 h。获得细菌发酵液,将上述发酵液稀释5倍,施用到2年生水杉盆栽苗中,每株施用50 mL稀释液(菌体量5×108CFU/mL),对照组接种等量摇培24 h后的NA培养基。施菌后实生苗置于室外,在施菌后20 d、40 d后采用手持叶绿素仪(SPAD-502 plus,Japan KONICA MINOLTA 制造公司)测定处理组和对照组相对叶绿素含量并测其苗高和地径。
1.2.7 根际细菌JYZ-SD5的鉴定 (1)菌体形态、特征观察及生理生化指标测定:参照《伯杰细菌鉴定手册》和《常见细菌系统鉴定手册》对菌株JYZ-SD5进行生理生化测定,描述菌落形态、特性。(2)16S rDNA基因序列分析及系统进化树:以提取的细菌总DNA为模板,用细菌16S rDNA的一对通用引物(上游引物27f(5'-AGAGTTTGATCCTGGCTCAG-3'), 下 游 引 物1492r(5'-TACGGCTACCTTGTTACGACTT-3') 对 细菌基因组DNA进行扩增。PCR反应体系:mix10 μL,上下游引物各 1 μL,DNA 模板 1 μL,ddH2O 7 μL。PCR反应条件:94℃预变性5 min;94℃变性30 s,55℃退火30 s,72℃延伸1 min,33个循环;72℃延伸10 min。将获得的PCR产物于1%的琼脂糖凝胶中电泳,EB染色,紫外灯下拍照。有条带则将获得的PCR产物送南京金斯瑞生物技术公司纯化测序。所测得的16S rRNA 与 Ez Taxon 和 Gen Bank中的序列进行基因序列比对,用 MEGA 8.0构建系统发育树。
1.2.8 数据处理 所测数据采用 Excel 2013 软件绘图,用prism cracked软件进行数据统计分析。
2 结果
2.1 根际细菌JYZ-SD5菌株的固氮、解磷和解钾特性
测定表明,根际细菌JYZ-SD5菌株在固氮培养基上可以生长(图1-A),在解有机磷培养基上生长出现透明圈(图1-B),说明该菌具有固氮、解有机磷的能力;同时在解钾能力检测中,发现对照组中钾含量为7.9 μg/mL,处理组中钾含量为11 μg/mL,比对照提高了2.1 μg/mL,菌株JYZ-SD5的解钾能力为39%(表1)。

表1 菌株JYZ-SD5解钾能力测定
2.2 根际细菌JYZ-SD5菌株产IAA能力
采用Salkowski比色法测定表明,在含有色氨酸培养液的比色反应中其颜色为淡红色,说明菌株JYZ-SD5具有一定产IAA的能力。采用标准IAA制作标准曲线定量分析,得出菌株JYZ-SD5培养1 d后在不含色氨酸的培养液中分泌的IAA浓度为3.938 5 μg/mL;在含L-色氨酸的培养液中IAA浓度达到7.152 0 μg/mL,且菌株JYZ-SD5在培养4 d后产IAA量达到最高。在不含色氨酸的培养液中分泌的IAA浓度最高为6.818 6 μg/mL;在含L-色氨酸的培养液中IAA浓度最高为12.199 6 μg/mL(图2)。
2.3 根际细菌JYZ-SD5菌株对几种林木病原真菌的拮抗活性
测定表明,该菌株胞外代谢产物对链格孢、七叶树壳梭孢、异色状拟盘多毛孢的抑菌效果较明显,其中对银杏叶枯病菌链格孢抑菌效果最大,抑菌率为20.22%(表2)。菌株JYZ-SD5与供试林木病原真菌的平板对峙结果表明,该菌株对茶树轮斑病菌异色状拟盘多毛孢,链格孢,茶树云纹叶枯病菌山茶球座菌,水杉赤枯病菌细极链格孢的抑制效果较为明显(图3)。
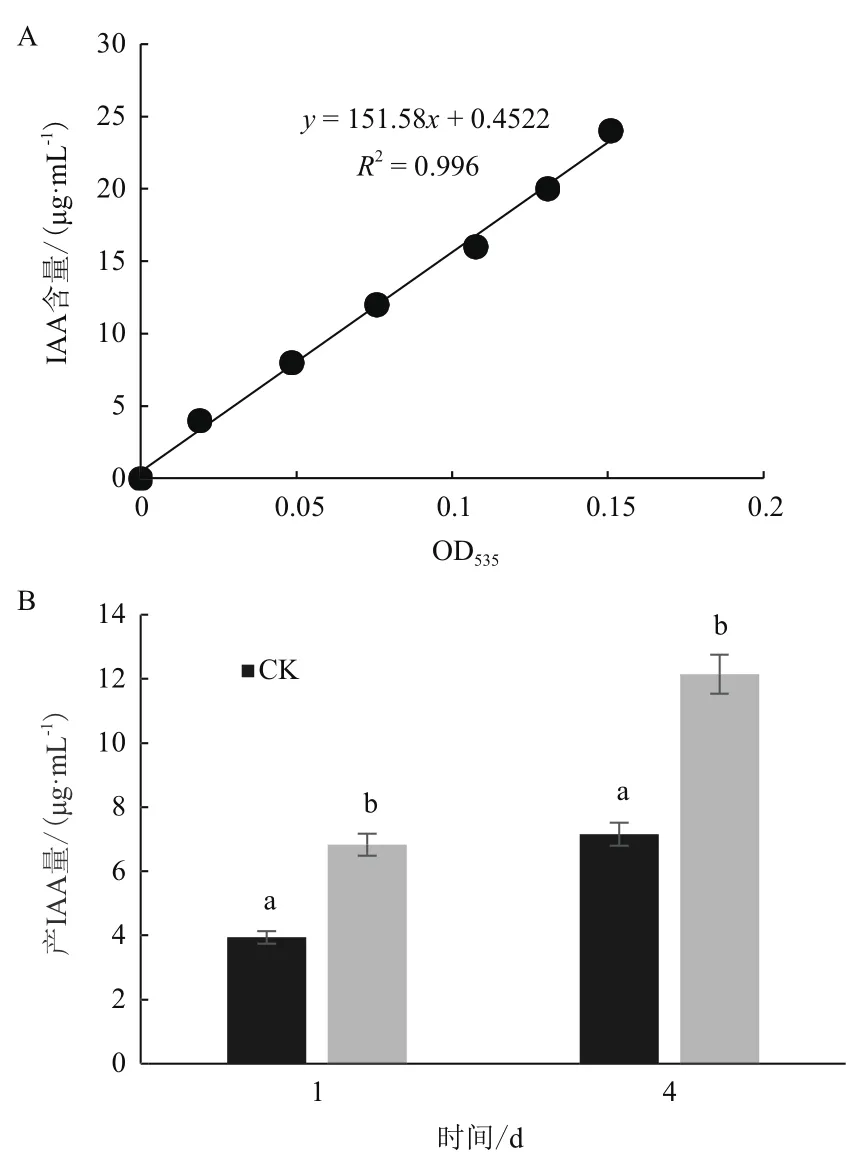
图2 根际细菌JYZ-SD5的IAA定量检测
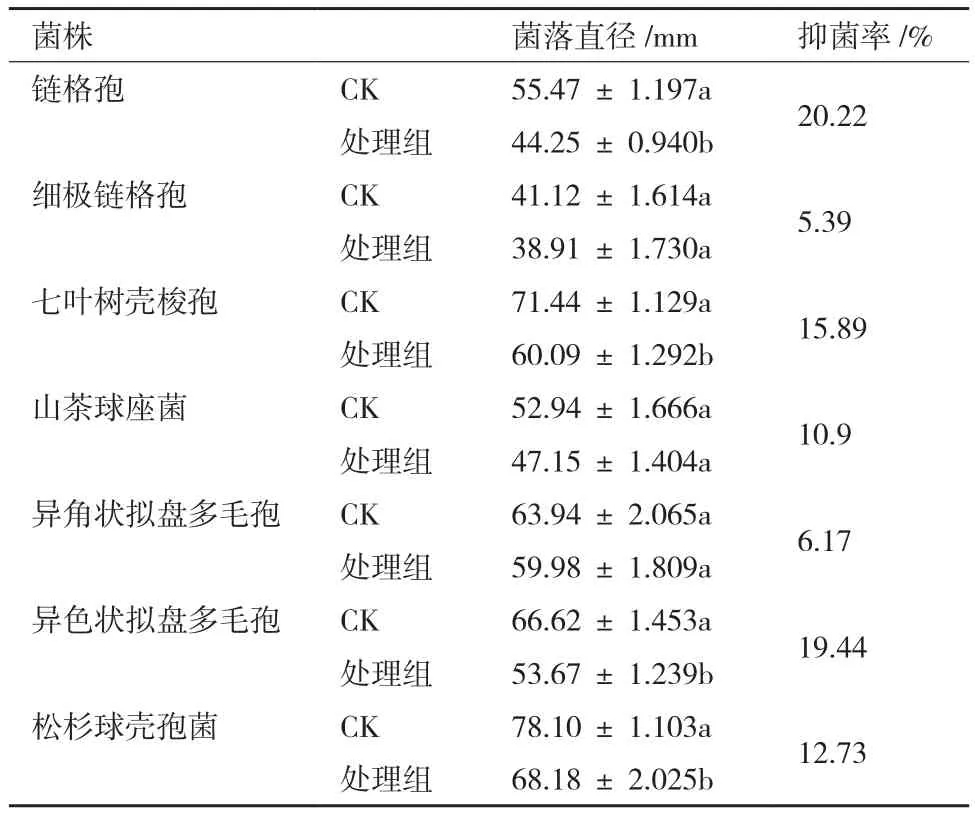
表2 根际细菌JYZ-SD5胞外代谢产物对供试林木病原真菌的抑菌率
2.4 根际细菌JYZ-SD5菌株的蛋白酶和纤维素酶活性
菌株JYZ-SD5在蛋白酶检测培养基、纤维素酶检测培养基上出现明显透明圈(图4),表明该菌含有蛋白酶和纤维素水解酶,具有一定的水解蛋白质和分解纤维素的能力。
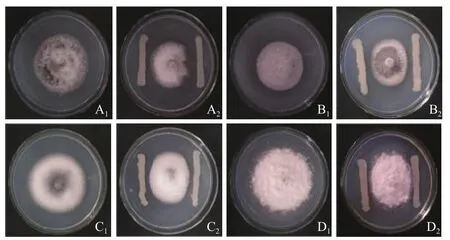
A:链格孢;B:细极链格孢;C:山茶球座菌;D:异色状拟盘多毛孢。其中,A1-D1为CK;A2-D2为接菌划线处理
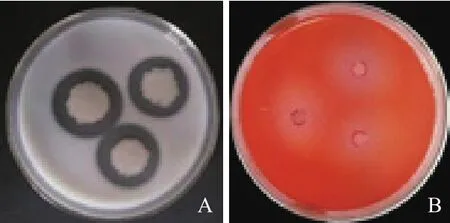
图4 根际细菌JYZ-SD5蛋白酶(A)和纤维素酶(B)检测
2.5 根际细菌JYZ-SD5菌株的金属耐受性
该菌在Mn7+、Gr6+存在的供试浓度下均可以生长,耐受浓度最高均可达到400 mg/L,在Ni2+浓度为0-200 mg/L可以较好的生长,在Cu2+浓度为5 mg/L及以上时该菌无法生长。在Gr6+、Ni2+浓度不超过200 mg/L时,该菌与在正常生长条件下时相比无明显差异;在Mn7+存在的情况下,该菌达到稳定期的时间明显推迟,说明Mn7+的存在会抑制该菌的繁殖速度,且当Mn7+浓度在200-300 mg/L时,该菌前30 h生长较好,30 h以后菌体量突然减少,说明在200-300 mg/L浓度下,该菌的耐受性减弱,培养后期会出现大量菌体死亡的现象。综合来看,重金属浓度在0-200 mg/L之间时,该菌对重金属胁迫耐受能力为 Ni2+> Gr6+> Mn7+> Cu2+;当重金属浓度高于200 mg/L时,只能在Mn7+、Gr6+存在的情况下缓慢生长(表3,图5)。
2.6 林木根际细菌JYZ-SD5对水杉的促生作用
施菌处理后,水杉实生苗的苗高和地径相对于对照组均有明显提高。施菌20 d后苗高和地径净增长量均提高了200%左右;施菌40 d后,水杉叶绿素相对含量相对于对照增加了37.8%,苗高和地径的净增长量分别提高了74%和88%,施菌处理和对照处理间存在显著性差异。说明接种根际细菌JYZSD5菌株对水杉的生长有显著的促进作用(图6)。

表3 林木根际细菌JYZ-SD5菌株在不同重金属胁迫下24 h后的生长状况

图5 根际细菌JYZ-SD5在不同重金属胁迫下的生长曲线

图6 根际细菌JYZ-SD5菌株对水杉的促生作用
2.7 林木根际细菌JYZ-SD5菌株鉴定
2.7.1 形态特征及生理生化特性 菌株JYZ-SD5在NA培养基上菌落为近圆形,边缘不整齐,乳白色,菌落中间无凸起,湿润、有光泽(图7-A、7-B)。菌体短杆状,无鞭毛,革兰氏阳性菌,在芽孢染色观察中可看到着绿色的芽孢,说明产芽孢(图7-C)。
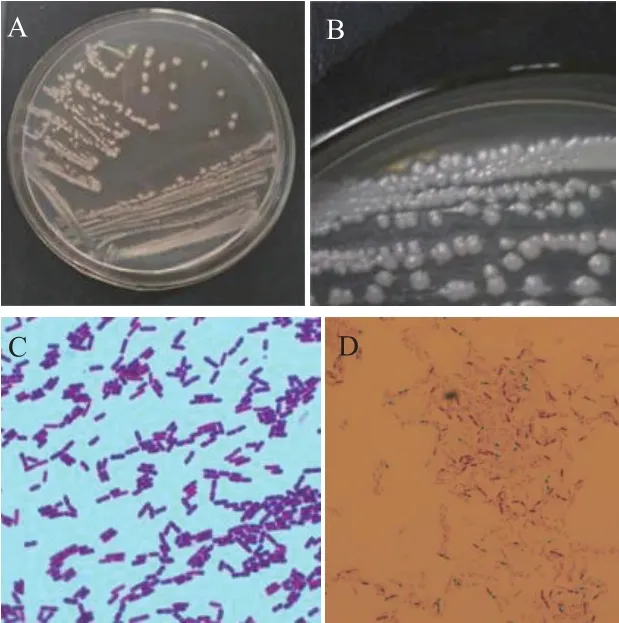
图7 根际细菌JYZ-SD5在NA平板上培养24 h后的菌落形态(A、B)及革兰氏染色(C)、芽孢染色(D)结果
菌株JYZ-SD5在VP实验、甲基红实验、吲哚实验、硝酸还原实验中结果呈阳性,可以水解淀粉、明胶。接触酶和氧化酶实验结果为阳性,在糖、醇发酵实验中,葡萄糖产酸检验结果为阳性,苯丙氨酸脱氨酶、木糖、甘露醇检验结果呈阴性(表4),结果与拟蕈状芽孢杆菌(Bacillus paramycoides)的特征基本相符。

表4 JYZ-SD5菌株生理生化测定结果
2.7.2 16S rDNA序列分析及系统进化树 菌株JYZSD5的16S rDNA扩增产物,用1%琼脂糖凝胶进行电泳检测结果,在1 500 bp附近有一条明亮的PCR目的条带,且无其他非特异性条带。表明16S rDNA扩增获得的目的DNA片段可用于测序。经测序获得菌株JYZ-SD5的16S rDNA基因有1 420个碱基(GenBank 登录号 :MH894221)。
通过将菌株JYZ-SD5与 Ez Taxon 和 GenBank 中的序列进行基因序列比对,并将同源性最高、且已定名的模式菌的序列信息,通过 MEGA8.0 软件构建系统发育树(图8),菌株JYZ-SD5与拟蕈状芽孢杆菌NH24A2模式菌株聚在一起,相似性达99%。根据系统发育树相似性分析,结合形态、生理生化特征比对,初步鉴定菌株JYZ-SD5为拟蕈状芽孢杆菌。
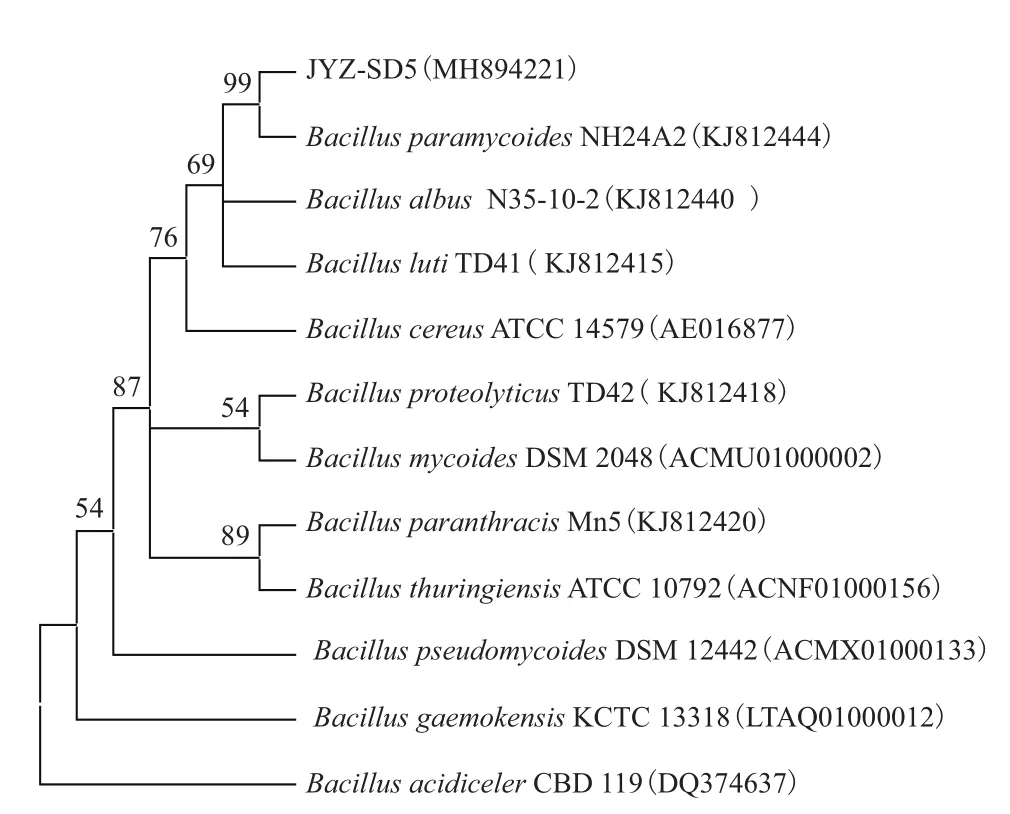
图8 基于16S rRNA基因序列构建的JYZ-SD5菌株系统发育树
3 讨论
植物根际细菌一般通过提高植物对根际养分吸收、产生植物激素类物质等促进植物生长[16-17]。从研究结果看,林木根际细菌JYZ-SD5具有固氮、解有机磷和解钾的能力,具有较高的产IAA能力,不加色氨酸前体下产IAA量为6.818 6 μg/mL。Yasmi[18]等发现具有固氮、解磷和产IAA能力的两种细菌Exiguobacteriumsp.和Stenotrophomonassp.对卡琪花蒂玛(Labisia Pumila)有促生作用。本研究通过盆栽试验发现,林木根际细菌JYZ-SD5对水杉有很好的促生作用,能明显提高水杉的株高和地径,施菌40 d后,相对于对照组苗高和地径的净增长量分别提高了74%和88%。说明林木根际细菌JYZSD5存在某种促进植物生长的机制。结合其固氮、解磷、解钾,产IAA的能力分析,推测该根际细菌对水杉的促生机制可能是通过提高植物对土壤氮、磷、钾养分的吸收,同时产生一定的促生物质来实现。
研究证实,部分重金属耐受性较高的根际细菌,在重金属污染地区可修复被污染的土壤,提高植物在污染地区生长状况[19];如Krishnendu等[20]发现Enterobacter aerogenesK6菌株对 Cd2+、Pb2+和 As3+的耐受性高,并且在镉胁迫下施用该菌,水稻幼苗生长显著增强。本研究发现,根际细菌JYZ-SD5菌株具有良好的耐重金属的能力,对Mn7+、Gr6+的耐受性最高均可达400 mg/L,对重金属Ni2+的耐受最高也可达200 mg/L。并且林木根际细菌JYZ-SD5菌株可以产纤维素酶和蛋白酶,而蛋白酶和纤维素酶能够降解真菌细胞壁中的蛋白质和纤维素,破坏病菌菌丝,使病菌生长受到抑制[21];同时本实验也证明该根际细菌JYZ-SD5对链格孢的抑菌率可达20.22%,对其他林木病原菌也具有一定的抑菌效果。由此推测JYZ-SD5菌株在重金属污染地区及林木病害防治方面具有较好的应用价值。
林木根际细菌JYZ-SD5经鉴定为拟蕈状芽孢杆菌,属芽孢杆菌属。拟蕈状芽孢杆菌是Liu等[22]2017年通过多相分类学方法,依据表型和16S rRNA基因序列分析数据,认为其属于蜡状芽孢杆菌中的低dDDH和ANI值菌株,而重新被划分出的一个新种Bacillus paramycoidessp. nov. type strain NH24A2T,该新种的模式菌株来自海洋。而本研究中将来自林木根际的菌株JYZ-SD5鉴定为拟蕈状芽孢杆菌,这是该菌分离自陆地生态系统的首次报道。研究结果表明拟蕈状芽孢杆菌JYZ-SD具有较好的促生抗逆特性,具有作为一种新型的微生物菌株资源开发应用的潜力。
4 结论
本研究通过对菌株JYZ-SD5生物学特性进行检测,得到该菌具有固氮、解磷、解钾、产IAA能力,对林木生长具有明显促进作用,对某些林木病原真菌具有一定的拮抗效果,对Ni2+、Gr6+、Mn7+等重金属耐受性较高,并通过形态观察、生理生化特性分析、16S rDNA序列分析将菌株JYZ-SD5鉴定为拟蕈状芽孢杆菌。
猜你喜欢
中国土壤与肥料(2021年5期)2021-12-02
湖南饲料(2021年4期)2021-10-13
农药科学与管理(2019年6期)2019-11-23
农药科学与管理(2019年6期)2019-11-23
农药科学与管理(2019年8期)2019-11-23
农民致富之友(2018年12期)2018-06-29
天然产物研究与开发(2018年3期)2018-05-07
现代园艺(2017年23期)2018-01-18
现代园艺(2017年21期)2018-01-03
中国蔬菜(2016年8期)2017-01-15
