
Morphometric discrimination between females of two isomorphic sand fly species, Phlebotomus caucasicus and Phlebotomus mongolensis (Diptera:Phlebotominae) in endemic and non-endemic foci of zoonotic cutaneous leishmaniasis in Iran
2019-05-13 12:21:50AzadAbsavaranMehdiMohebaliVahidehMoinVaziriAlirezaZahraeiRamazaniAmirAhmadAkhavanFaribaMozaffarianSayenaRafizadehYavarRassi
Azad Absavaran, Mehdi Mohebali, Vahideh Moin-Vaziri, Alireza Zahraei-Ramazani, Amir Ahmad Akhavan, Fariba Mozaffarian, Sayena Rafizadeh, Yavar Rassi✉
1Department of Medical Entomology and Vector control, School of Public Health, Tehran University of Medical Sciences, Tehran, Iran
2Department of Medical Parasitology and Mycology, School of Public Health, Tehran University of Medical Sciences, Tehran, Iran
3Department of Medical Parasitology and Mycology, School of Medicine, Shahid Beheshti University of Medical Sciences, Tehran, Iran
4Insect Taxonomy Research Department, Iranian Research Institute of Plant Protection, Tehran, Iran
5Ministry of Health and Medical Education, Tehran, Iran
Keywords:Leishmaniasis Phlebotomus caucasicus Phlebotomus mongolensis Morphometry Discriminant Functional Analysis Principal Component Analysis Iran
ABSTRACT Objective: To delineate reliable morphological characteristics for identifying and separating female Phlebotomus caucasicus and Phlebotomus mongolensis which exist sympatrically in the main foci of zoonotic cutaneous leishmaniasis in Iran.Methods: Sand flies were collected using sticky trap papers from active colonies of rodent burrows installed from 16 catching sites. Morphometric measurements were analyzed of 87 Phlebotomus caucasicus and 156 Phlebotomus mongolensis. Univariate and multivariate analysis were carried out to determine significant morphometric variables for discrimination of the two species. Finally, seven morphological characteristics of 65 female Phlebotomus caucasicus and 124 female Phlebotomus mongolensis were described.Results: Univariate and multivariate analyses of 10 morphometric variables via Discriminant Function Analysis (DFA) and Principal Component Analysis (PCA) showed that five morphometric variables had an accuracy of 100% for discriminating female Phlebotomus caucasicus and Phlebotomus mongolensis. Moreover, PCA revealed that the five morphometric variables with the highest loadings separated these two species. Morphological studies on antennal flagellum (and its associated structures) and mouth-parts of female specimens demonstrated significant differences in several structures.Conclusions: The results show that morphological and morphometrical features can be used to discriminate two female isomorphic species, Phlebotomus caucasicus and Phlebotomus mongolensis accurately.
1. Introduction
Sand flies belong to the subfamily Phlebotominae which is one of the six Psychodidae subfamilies with more than 800 species described worldwide, of which 98 species are proven and suspected vectors of Leishmania parasites[1-3].
Zoonotic cutaneous leishmaniasis (ZCL) due to Leishmania major(L. major) is endemic in rural regions of 17 out of 31 provinces of Iran and approximately 80% of reported leishmaniasis cases in the country are ZCL form. Phlebotomus papatasi is the main vector of ZCL to human in Iran. Phlebotomus (Paraphlebotomus)caucasicus (Ph. caucasicus) group belongs to the subgenus Paraphlebotomus, which has been considered as a complex species including Ph. caucasicus, Phlebotomus mongoloensis (Ph.mongolensis) and Phlebotomus andrejevi, playing a main role in maintenance of enzootic cycle of L. major among rodent reservoir hosts. Ph. caucasicus and Ph. mongolensis not only participate in the transmission cycle of ZCL among reservoir hosts but also play an important role as secondary vectors in the transmission of leishmaniasis to humans[4].
Despite the fact that the males of Ph. caucasicus, Ph. mongolensis and Phlebotomus andrejevi are distinguishable by their external genitalia, the females of these species are indistinguishable morphologically and have similar taxonomic characters[5].Indistinguishable morphological characters of females of Ph.caucasicus group species prevent the direct investigation of their role in epidemiology of ZCL, and maintaining of L. major transmission and other Leishmania species of gerbils (reservoir hosts), especially in localities that Ph. caucasicus and Ph. mongolensis are present sympatrically.
Phlebotomine sand flies of the subgenus Paraphlebotomus Theodor, 1948 exist mainly in palearctic areas and are comprised of 14 species[6]. The morphological characteristics of females of the subgenus Paraphlebotomus include: the pharynx having a large scale-like teeth with smooth margins, which appears like a network and spermathecae usually having the terminal segment much larger than the other segments. The morphological characteristics of males of this subgenus include: fleshy basal lobes on the inner surface of the coxite bearing tufts of long setae and the styles with only four spines[7].
There are eight species and two subspecies of the subgenus Paraphlebotomus in Iran: Phlebotomus alexandri Sinton, 1928;Phlebotomus andrejevi Shakirzyanova, 1953; Ph. caucasicus Marzinowsky, 1917; Phlebotomus grimmi Porchinski, 1876;Phlebotomus jacusieli Theodor, 1947; Phlebotomus kazeruni Theodor and Mesghali, 1964; Ph. mongolensis Sinton, 1928; Phlebotomus sergenti sergenti Parrot, 1917 and Phlebotomus sergenti similis Perfil’ev, 1963[8].
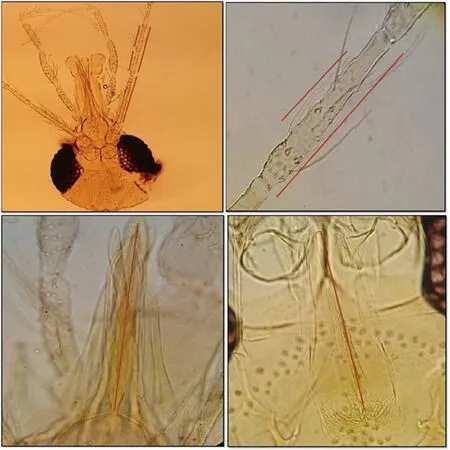
Figure 1. Morphometric characters used to discriminate females of Phlebotomus caucasicus and Phlebotomus mongolensis. (A) Length of antennal segment 3 (A3),segment 4 (A4), segment 5 (A5) and segments 4 and 5 (A4+A5). (B) Length of ascoid on A4 (Ascoid4) and ratio of Ascoid4/A4. (C) Length of the labrum or epipharynx (Epi). (D) Length of the pharynx (Ph.L).
Many attempts have been made in Iran to differentiate the females of Ph. caucasicus and Ph. mongolensis using molecular methods by Cytochrome b (Cytb) and Internal Transcribed Spacer 2 (ITS2)markers[9,10]. These molecular techniques are often time-consuming and also do not separate these species morphologically.
According to the role of Ph. caucasicus and Ph. mongolensis in ZCL transmission cycle, finding reliable morphological method for identification of these female sand flies is very valuable.
The morphological characters of the sand flies head such as antennal flagellumers and its associated structure (Ascoid or Sensilla chaeticum) have been used for differentiating of Phlebotominae sand flies at inter and intra-species levels[11,12].
Traditional morphometric investigations and multivariate analysis of morphometric characters have been used to resolve taxonomic problems in Phlebotominae sand flies, especially in identifying the closely related species such as isomorphic or cryptic species[13].
The present study, describes the main morphological characteristics for separating the females of 203 Ph. caucasicus and Ph. mongolensis collected from allopatric localities of ZCL foci in central and southern Iran (Isfahan and Fars provinces, respectively) and females of 40 Ph. mongolensis collected from non-endemic focus of ZCL in north-western Iran (Ardabil province), which have not been described previously. These taxonomic tools and morphological characters described in this study are useful methods for separation of closely related species of Ph. caucasicus group and for epidemiological investigations of ZCL in Iran.
2. Materials and methods
2.1. Sand fly specimens and processing
Ph. caucasicus group sand flies were collected from different allopatric localities in Isfahan, Fars and Ardabil provinces (central,southern and north-western Iran, respectively) and from sympatric locations in Golestan province (north-eastern Iran) (Table 1). Sand flies were collected using sticky trap papers from active colonies of rodent burrows from 16 catching locations during June to October 2016. The sticky trap papers were installed after sunset and were collected before sunrise. Collected sand flies were conserved in 96% ethanol for subsequent processing. For species morphological identification, the head and the last two abdominal segments of each female specimen were dissected in a drop of sterile normal saline using sterile dissecting needles and were mounted on slide in a drop of Berlese’s or Puri’s medium.
In this study, in order to reduce the incorrect identification resulting from the close morphological similarity between the two species,only females of Ph. caucasicus and Ph. mongolensis from allopatric locations (Isfahan, Fars and Ardabil provinces) were examined.Confirmation of species identification for these female species was carried out on associated males, which can be distinguished only by morphological characters in external genitalia (Table 1). Ph.caucasicus group sand flies were analyzed from eleven allopatric localities, being four in Isfahan province (Habib-abad, Ali-abad,Abbas-abad and Siyan) four in Fars province (Raja-abad, Kouhsabz,Sedegh-abad and Band-e-amir) and three in Ardabil province (Khiav,Qourt tappeh and Alni).
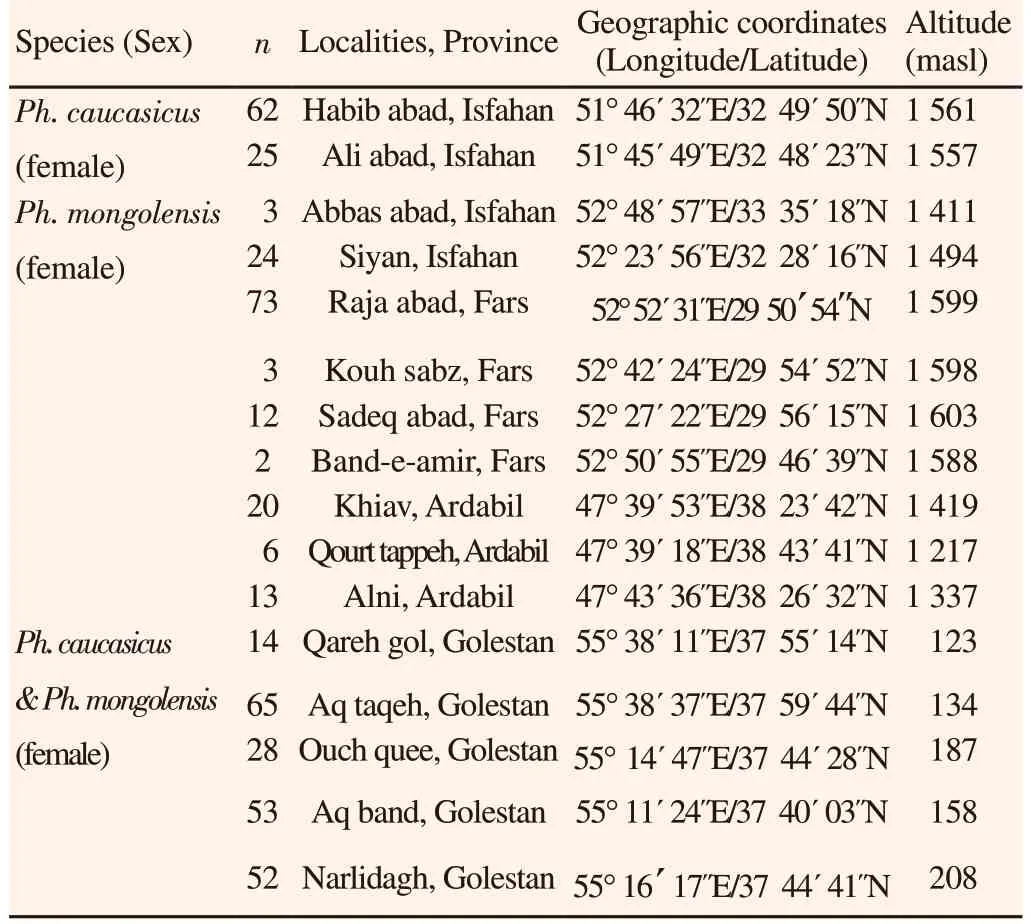
Table 1. Collection sites and number of Ph. caucasicus group sand flies collected from sympatric (Golestan) and allopatric (Isfahan, Fars and Ardabil)localities in Iran.
2.2. Morphometric analysis
Measurements of morphometric characters were analyzed from allopatric localities on a total of 87 females Ph. caucasicus and 156 females Ph. mongolensis (totally 243 specimens). 10 morphometric characters located on the head of sand fly specimens which were measured, including: 1) Length of antennal segment 3 (A3), 2)Length of antennal segment 4 (A4), 3) Length of antennal segment 5 (A5), 4) Length of antennal segments 4 and 5 (A4+A5), 5) Ratio of A3/A4+A5, 6) Length of ascoid on A4 (Ascoid4), 7) Ratio of Ascoid4/A4, 8) Length of labrum or epipharynx (Epi), 9) Ratio of A3/Epi and 10) Length of Pharynx (Ph.L) (Figure 1). Morphometric characters were measured using calibrated ocular micrometer mounted on the ocular lens and also 10× and 40× objective lens according to the size of the measured character in millimeters (mm)units.
2.3. Statistical analysis
Univariate statistical analysis of 10 morphometric characters (Mean,Standard deviation, and Ranges) was carried out for two mentioned species. In order to check the normality of data and the homogeneity of variances, Kolmogorov-Smirnov test and Levene’s test were used respectively. The significant mean differences of measured characters between Ph. caucasicus and Ph. mongolensis were evaluated using the Independent Samples t-test for the parametric data and Mann-Whitney test for the nonparametric data. The significant level of statistical analysis was P<0.05 for all measures of morphometric characters.
Moreover, multivariate statistical analyses for measured data of morphometric characters were used. For determining the most important morphometric variables for discrimination of the two species, Discriminant Function Analysis (DFA) was used. With the stepwise discriminant analysis, the probability of a priori grouping and their coefficients was extracted to maximize the ratio of between species variation to within-species variation. Box’s M test for the multivariate assumptions of equality of covariance matrices was used. Another statistical multivariate analysis used to assess the data set of morphometric variables was Principal Component Analysis(PCA). The PCA scores were computed using vector products with the original data. All statistical analyses of the morphometric data were done using SPSS version 24 and PAST 2.17 softwares.
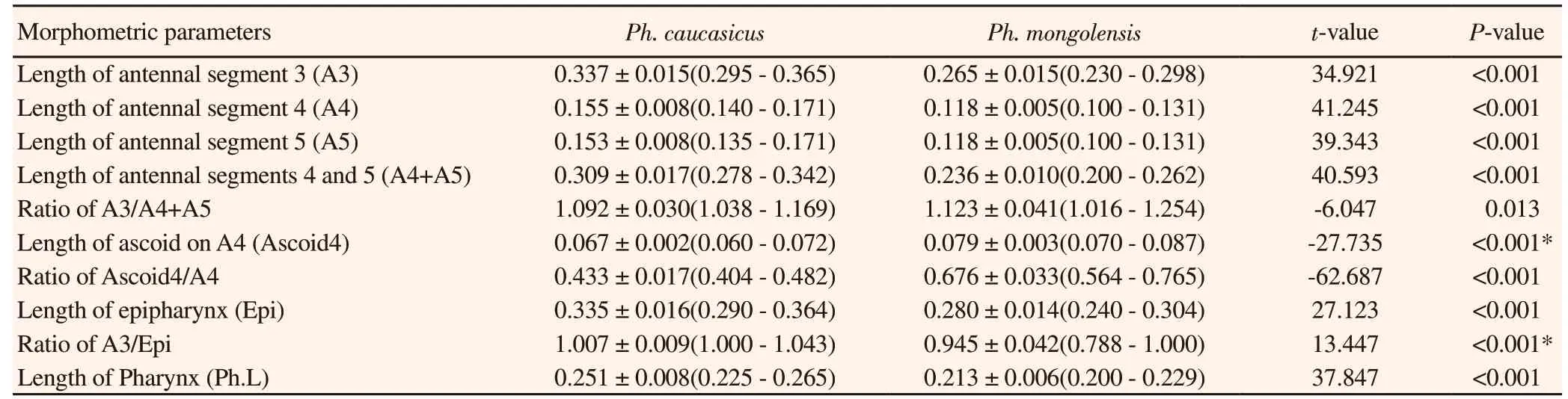
Table 2. Values of 10 morphometric measurements (in mm) and ratios of females Phlebotomus caucasicus and Phlebotomus mongolensis collected from allopatric locations in Iran.
2.4. Morphological analysis
Using light microscope, seven external and internal morphological characteristics on 65 females Ph. caucasicus (of 87 specimens) and 124 females Ph. mongolensis (of 156 specimens) were observed.These morphological characters includes: 1) reaching out the distal tip of ascoid3 (Ascoid3 apex)o the distal margin of A3, 2) relative length of ascoid on A4 (Ascoid4), 3) number of observed antennal segments with magnification of 40X objective lens, 4) appearance of pharyngeal armature, 5) palpal formula, 6) number of spermathecal segments and 7) size and shape of apical segment of spermatheca.
3. Results
3.1. Univariate analysis of morphometric data
Statistically, the results of univariate analysis of 10 measured morphometric characters showed significant differences between the specimens of Ph. caucasicus and Ph. mongolensis (P≤0.05).Morphometric characters for Ph. caucasicus tended to be significantly larger in the length of A3, A4, A5, A4+A5, Epi and Ph.L compared with Ph. mongolensis, but Ph. mongolensis tended to be more significantly larger in the length of ascoid on A4 (Ascoid4) compared with Ph. caucasicus. The results of the Kolmogorov-Smirnov test for normality of data and Levene’s test for the homogeneity of variances applied to each character were not significant (P>0.05) except for ascoid 4 and A3/Epi (Table 2).
3.2. Multivariate analysis of morphometric data
3.2.1. Discriminant Function Analysis (DFA)
Box’s M test for the homogeneity of covariance matrices was significant (M=46.2, P<0.001). Significant separation between Ph.caucasicus and Ph. mongolensis was achieved following the stepwise discriminant analysis selected five most informative variables out of the 10 measured morphometric characters, including length of antennal segment 3 (A3), length of antennal segments 4 and 5(A4+A5), length of ascoid on A4 (Ascoid4), ratio of Ascoid4/A4 and length of epipharynx (Epi). One significant discriminant function was retained by the stepwise procedure, which had the highest ratio of between group to within-group sum of squares (Wilk’s lambda =0.036, χ2 = 790.3, df= 5, P<0.001). The coefficients of standardized and unstandardized canonical discriminant weights and discriminant loadings of each of these morphometric characters are presented in Table 3. The canonical discriminant function evaluated at group means are -6.889 for Ph. caucasicus and 3.842 for Ph. mongolensis,indicating the discriminant scores for Ph. caucasicus were far smaller than that of Ph. mongolensis. The goodness-of-fit of this model in classifying the two groups was analysed using the eigenvalue of 26.68. Canonical correlation was 0.982 and showed that 100% of the total variation between Ph. caucasicus and Ph. mongolensis was explained by the discriminant function.
The histogram of the discriminant scores of the individuals of Ph.caucasicus and Ph. mongolensis based on analysis of discriminant function revealed obvious discrimination between the two mentioned species (Figure 2).
3.2.2. Principal Component Analysis (PCA)
The initial principal component analysis of 10 measured morphometric characters indicated that two axes had an eigenvalue of 1.00 or higher. The first principal component (PC1) and second principal component (PC2) accounted for 86.6% and 7.4% of overall variance in Ph. caucasicus and Ph. mongolensis specimens respectively and when combined, these factors accounted for 94.1 % of overall variance(Table 4 and 5).
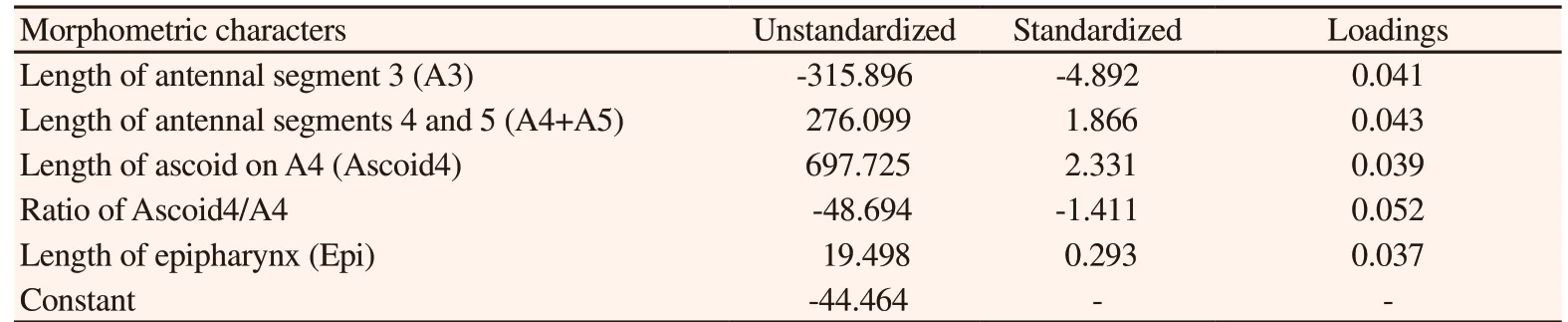
Table 3. Canonical discriminant coefficients and loadings for discriminant function 1.
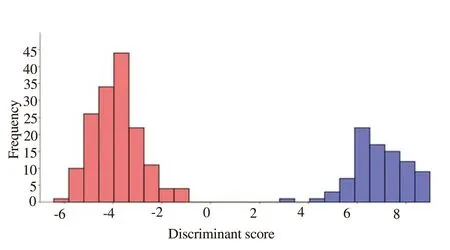
Figure 2. Frequency distribution of discriminant scores for females ofPhlebotomus caucasicus (Red bars) and Phlebotomus mongolensis (Blue bars).
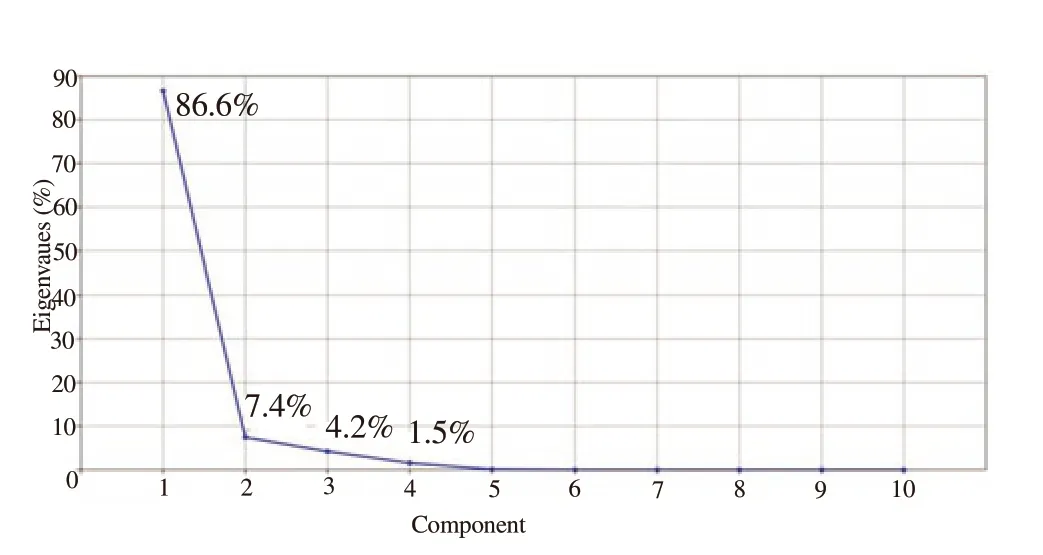
Figure 3. A scree plot highlighting the relationship between the eigenvalues and the number of axis in a principal component analysis of 10 morphometric measurements of Phlebotomus caucasicus and Phlebotomus mongolensis.
The scree plot indicates the relationship between the relative significance of the eigenvalues and the number of axis (Figure 3).
The result of PCA scatterplot and principal component matrix,confirmed that which measurements are really different and separate the females Ph. caucasicus and Ph. mongolensis. The first axis (X axis; PC1) was highly negatively correlated to the ratio of ascoid4/A4 (loadings ≥ 0.8), and correlated to the length of antennal segment 3 (A3) and the length of antennal segments 4 and 5 (A4+A5)(loadings 0.25). The second axis (Y axis; PC2) was positively correlated with the length of ascoid on A4 (Ascoid4) (loadings ≥0.8), and correlated to the ratio of A3/Epi (loadings ≥ 0.45) (Table 5 and Figure 4).

Table 4. The eigenvalues, percent variance and cumulative variance of the axes from the principal component analysis of 10 morphometric measurements of Phlebotomus caucasicus and Phlebotomus mongolensis.
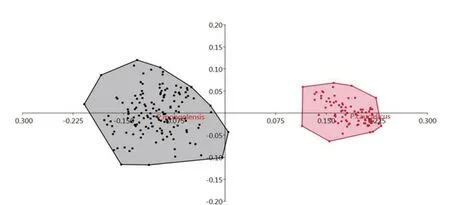
Figure 4. Principal Component Analysis (PCA) of females of Phlebotomus caucasicus (pink color) and Phlebotomus mongolensis (Grey color).
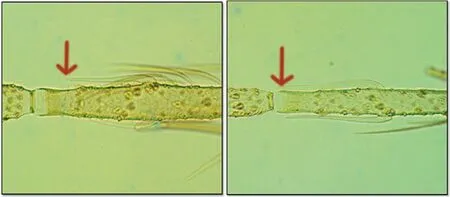
Figure 5. Reaching out the Ascoid3 apex to the distal margin of A3 in females of (A) Phlebotomus caucasicus and (B) Phlebotomus mongolensis.

Figure 6. Relative length of Ascoid4/A4 in females of (A) Phlebotomus caucasicus and (B) Phlebotomus mongolensis.

Figure 7. The number of observed antennal segments with magnification of 40X objective lens in females of (A) Phlebotomus caucasicus and (B) Phlebotomus mongolensis.
3.3. Morphological analysis
The distal tip of ascoids on A3 (Ascoid3 apex) extends to the distal margin of A3 (flagellomere I) in 95.1% of examined Ph. mongolensis specimens while, in 6.2% of examined Ph. caucasicus specimens(Figure 5). Another morphological characteristic was observed, the antennal Ascoid4, that showed differences in its relative length on A4 (flagellomere Ⅱ) for both Ph. caucasicus and Ph. mongolensis.Relative length of Ascoid4/A4 in Ph. caucasicus is about ¼ or length of segment which observed in 95.4% of specimens. While,relative length of Ascoid4/A4 in Ph. mongolensisis is about length of segment which was observed in 96% of specimens (Figure 6).The number of observed antennal segments with magnification of 40× objective lens is two segments (A3 and about length of A4)for 92.3% of Ph. caucasicus specimens and three segments (A3, A4 and about length of A5) for 93.5% of Ph. mongolensis specimens(Figure 7). The structure of pharynx for both Ph. caucasicus and Ph.mongolensis is composed of network of posterior lines and a small group of blunt and hard teeth in the middle armature. The median teeth in Ph. caucasicus are opaque with smooth outlines (81.5%)while, the median teeth in Ph. mongolensis are clear with thick outlines (79%) (Figure 8). The spermathecae structure and palpal formula show resemblance in the shape and the number of segments(rings) and length of palpomere respectively, for both Ph. caucasicus and Ph. mongolensis and no significant differences were found(Figure 9). All the data regarding sample size and percentage are listed in Table 6.

Table 5. Characterisation of Phlebotomus caucasicus and Phlebotomus mongolensis using the loadings of principal component analyses on 10 morphometric variables.
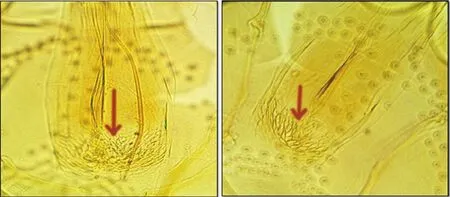
Figure 8. Appearance of pharyngeal armature in females of (A) Phlebotomus caucasicus and (B) Phlebotomus mongolensis.
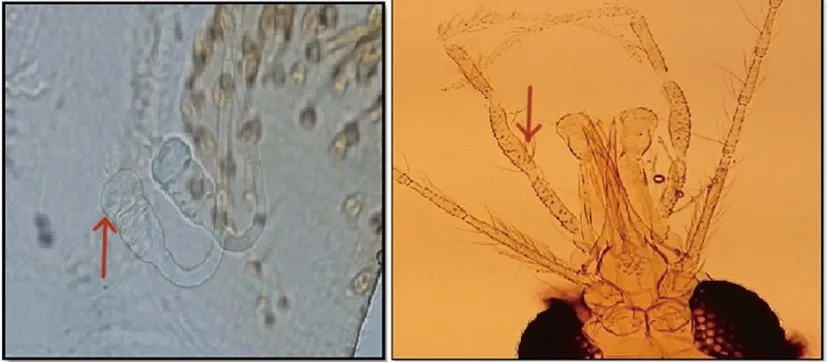
Figure 9. Structure of spermatheca and palpal formula in females of Phlebotomus caucasicus and Phlebotomus mongolensis.
4. Discussion
Identification of Phlebotominae sand flies species as a disease vectors and discriminating among isomorphic or cryptic species are the main factors in epidemiological study on leishmaniasis[14].
This study attempts to investigate the morphological identification for isomorphic species of Ph. caucasicus group and to address whether morphological and traditional morphometric analyses can be applied to discriminating and identification of females of Ph.caucasicus and Ph. mongolensis specimens collected from different geographical regions in Iran. The results demonstrate that the measurements of antennal segments and its associated structures as well as mouth-parts could be used to reliably separate the females of
Ph. caucasicus and Ph. mongolensis.
Ph. caucasicus, firstly reported from Transcaucasia, a predominant species of mud deserts and mud hills, rarely attacks human beings and usually lives in rodent burrows[7]. This species is mainly distributed in the north of Iran, in the Elburz Mountains, east to Mashhad and becomes rarer towards the south. This species is a central and western asiatic species which extends from west to the Caucasus and obviously enters Iran from the north[15]. Natural promastigote infection were isolated from Ph. caucasicus collected from gerbil and jird burrows in the main focus of Isfahan province in Iran and typed by isoenzymes assays as L. major zymodeme MON-26[16]. Natural promastigote infection rates from 3.3%-20% have been reported in Ph. caucasicus specimens collected from rodent burrows[17,18]. This species was also found naturally infected with promastigotes in a new focus of visceral leishmaniasis in northwest of Iran[19]. Furthermore, L. major and Leishmania gerbilli were detected and identified in Ph. caucasicus by Nested PCR and DNA sequence analysis from specimens collected from rodent burrows and animal shelters in Isfahan and Golestan Provinces, respectively[5].These findings accompanied by 20% human blood index indicate that Ph. caucasicus is a natural vector of L. major in central Iran and a secondary vector to humans in the country[20].
The first phenotypic and genotypic variations studies on males Ph.caucasicus in Iran, based on morphometric and molecular analyses respectively, was carried out according to the antennal segments and external genitalia. According to differences in implantation of the two distal spines on the style, the number of setae on the basal lobe of the coxite and the length of A3 in males Ph. caucasicus confirmed the existence of two morphotypes C and G within Ph.caucasicus populations. Morphotype C, described as Ph. caucasicus,was reported in all foci of ZCL, but morphotype G, described as Phlebotomus grimmi, was collected only in three provinces in the north-western part of the country that are free of ZCL. Sequencing of mtDNA showed a partial correlation between morphotypes and haplotypes. A correlation between haplotypes and the geographicalorigin of the specimens was also observed[21].
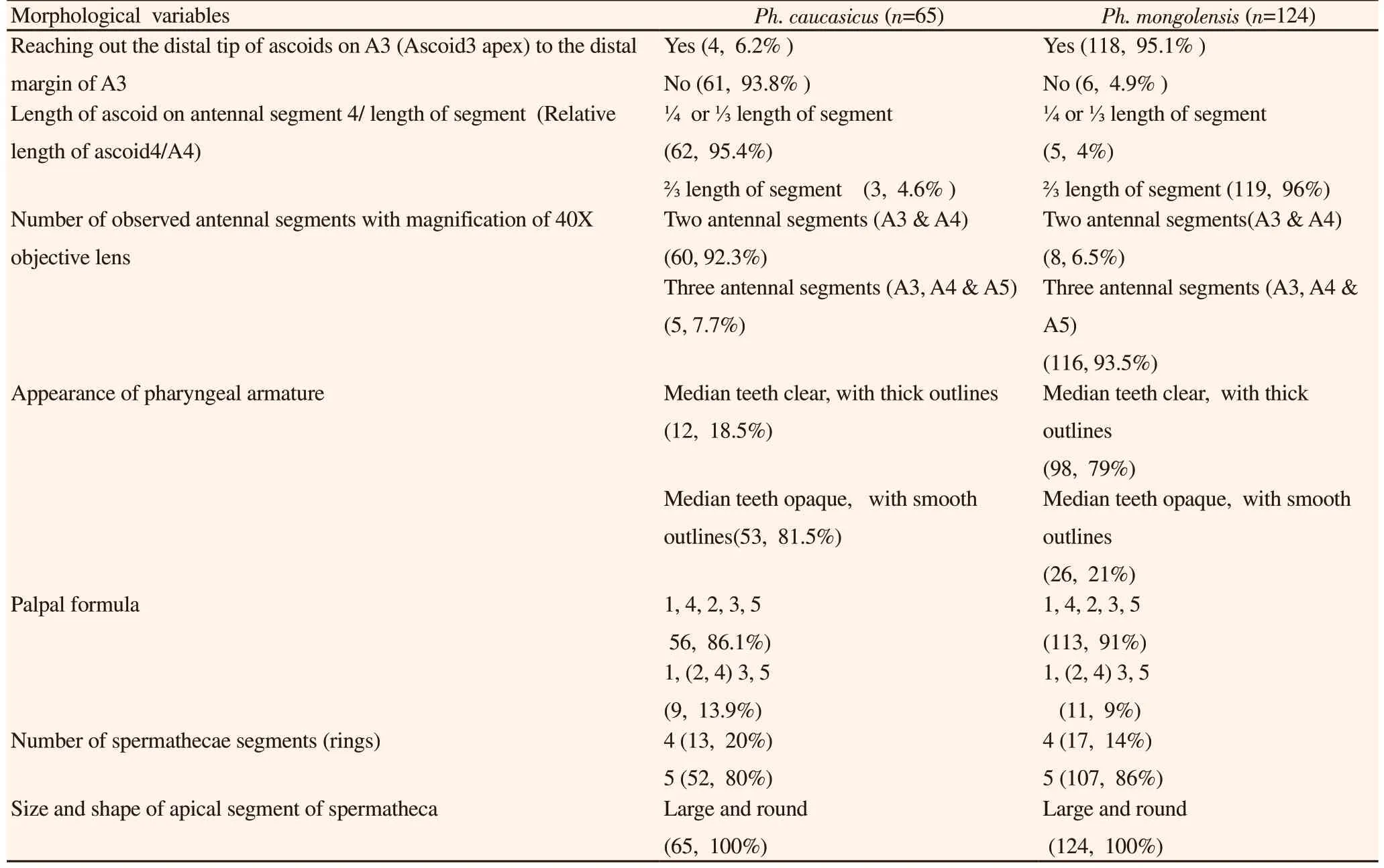
Table 6. Results of seven observed morphological variables of females Phlebotomus caucasicus and Phlebotomus mongolensis collected from allopatric regions of Iran[n(%)].
The first full description of Ph. mongolensis was given by Raynal.Ph. mongolensis, usually inhabits in rodent burrows in mud,sandy desert and highlands of China, Mongolia and Kazakhistan.This species is resistant to cold winter and prefer to suck blood of squirrels and is a main vector of VL in Central Asia and Kazakhistan[7]. Nadim and Seyedi-Rashti found this sand fly naturally infected with promastigote in gerbil burrows in Iran[22].
In the present research we have thoroughly studied 7 external and internal morphological characteristics and 10 external and internal morphometric characters of allopatric females of Ph. caucasicus and Ph. mongolensis to determine reliable morphological characters for identifying and separating sympatric female specimens of these species. Out of 10 studied morphometric characters, five discriminating characters including length of antennal segment 3(A3), length of antennal segments 4 and 5 (A4+A5), length of ascoid on A4 (Ascoid4), ratio of Ascoid4/A4 and length of epipharynx(Epi) were selected by Discriminant Function Analysis (DFA) with high discriminant loading scores and confirmed the morphological characters described in this study.
However, according to the loadings on these five variables, the relative length of ascoids on antennal segment4 (ratio of Ascoid4/A4)was found to be the most discriminating character differentiating Ph.caucasicus (Ascoid4/A4 = 0.404 - 0.482, n=87) from Ph. mongolensis(Ascoid4/A4 = 0.564 - 0.765, n=156) in allopatric localities. In Phlebotomine sandflies, ascoids (sensilla chaetica) form an important taxonomic character to differentiate between the species and were used to differentiate closely related species of leishmaniasis vectors especially among the species of Phlebotomus papatasi, Phlebotomus bergeroti and Phlebotomus duboscqi[13].
Theodor and mesghali stated that the females of Ph. mongolensis are difficult to distinguish from that of Ph. caucasicus, since both have a weakly developed pharyngeal armature. The only difference is the presence of long ascoids on the antennae in Ph. mongolensis, while those of the females of Ph. caucasicus are short[15]. These results support the findings of our study regarding the length of Ascoid4 and the ratio of ascoid4/A4.
The current study according to the multivariate statistical analysis(DFA and PCA), confirms that Ph. caucasicus and Ph. mongolensis can be separated by the length of antennal segment 3 (A3), length of antennal segments 4 and 5 (A4+A5), length of ascoid on A4(Ascoid4), ratio of Ascoid4/A4 and ratio of A3/Epi.
In overall, according the results of univariate and multivariate morphometric (DFA and PCA) and morphological analyses in present study, the following morphological characteristics can be used to identify and discriminate between females of Ph. caucasicus and Ph. mongolensis:
1-Morphometric analyses (DFA & PCA):
(1)Length of antennal segment 3 (A3): Length of A3 in Ph.caucasicus (range: 0.295 - 0.365 mm) is longer than the length of A3 in Ph. mongolensis (range:0.230 - 0.298 mm)
(2) Length of antennal segments 4 and 5 (A4+A5): Length of
A4+A5 in Ph. caucasicus (range: 0.278 - 0.342 mm) is longer than the length of A4+A5 in Ph. mongolensis (range: 0.200 - 0.262 mm)
(3)Length of ascoid on A4 (Ascoid4): Length of Ascoid4 in Ph.caucasicus (range: 0.060 - 0.072 mm) is shorter than the length of Ascoid4 in Ph. mongolensis (range: 0.070 - 0.087 mm)
(4)Ratio of Ascoid4/A4: <0.5 in Ph. caucasicus (range: 0.404 -0.482); > 0.5 in Ph. mongolensis (range: 0.564 - 0.765)
(5)Length of epipharynx (Epi): Length of epipharynx in Ph.caucasicus (range: 0.290 - 0.364 mm) is longer than the length of epipharynx in Ph. mongolensis (range: 0.240 - 0.304mm)
(6)Ratio of A3/Epi: 1 in Ph. caucasicus (range: 1.000 - 1.043); 1 in Ph. mongolensis (range: 0.788 - 1.000)
2-Morphologic analyses:
(1)The distal tip of ascoids on A3 (Ascoid3 apex) in Ph. caucasicus specimens does not extends to the distal margin of A3 (93.8%)while, the Ascoid3 apex in Ph. mongolensis specimens extends to the distal margin of A3 (95.1%)
(2)Relative length of Ascoid4/A4 in Ph. caucasicus is about or length of segment (95.4%) while; relative length of Ascoid4/A4 in Ph. mongolensis is about length of segment (96%)
(3)Observation of two antennal segments (A3 and about length of A4) with magnification of 40X objective lens for Ph. caucasicus specimens (92.3%), and three antennal segments (A3, A4 and about length of A5) for Ph. mongolensis specimens (93.5%)
(4)The structure of pharynx for both Ph. caucasicus and Ph.mongolensis composed of network of posterior lines and a small group of blunt and hard teeth in the middle armature. The median teeth in Ph. caucasicus are opaque with smooth outlines (81.5%),while, the median teeth in Ph. mongolensis are clear with thick outlines (79%)
(5)The females of Ph. caucasicus share some of its characters with Ph. mongolensis such as pharyngeal armature weakly developed,less sclerotized lateral groups of teeth, palpal formula (1,4,2,3,5 or 1,(2,4),3,5) and 4-5 segmented spermatheca with global apical segment.
The results of this study indicated that morphological and morphometrical analyses were very important to species identification and discrimination in the cases of cryptic or isomorphic species. Further studies should be done to clarify the more morphological characters that contribute to the differences between females of Ph. caucasicus and Ph. mongolensis and to the different morphotypes of these species.
Acknowledgement
This study was supported by Tehran University of Medical Sciences(Project No. 27252).
Conflict of interest statement
The authors declare that there is no conflict of interest.
 Asian Pacific Journal of Tropical Medicine2019年4期
Asian Pacific Journal of Tropical Medicine2019年4期
- Asian Pacific Journal of Tropical Medicine的其它文章
- Disseminated nocardiosis due to Nocardia otitidiscaviarum: A case report and literature review
- Latent tuberculosis infection among medical students in Malaysia
- Misinformation on salt water use among Nigerians during 2014 Ebola outbreak and the role of social media
- Wild chive oil is an extremely effective larvicide against malaria mosquito vector Anopheles stephensi
- Characterization of the salivary microbiome in healthy Thai children
- Herbal remedies, vaccines and drugs for dengue fever: Emerging prevention and treatment strategies