
黄土丘陵沟壑区植被恢复演替过程中根系行为特征
2019-05-13 01:50常恩浩徐国策赵宾华苏远逸冯朝红
生态学报 2019年6期
常恩浩,李 鹏,*,肖 列,徐国策,赵宾华,苏远逸,冯朝红
1 西安理工大学省部共建西北旱区生态水利国家重点实验室,西安 7100482 旱区生态水文与灾害防治国家林业局重点实验室,西安 710048
黄土丘陵地区因特殊而深厚的黄土母质与较为严重的水土流失等,土壤保水、持水性能差,养分含量低,因而土壤理化性质是黄土丘陵区植被恢复与建设的主要限制因子之一[1]。植被的恢复也可以起到减少水土流失以及改良土壤环境的作用。但植被恢复过程缓慢而又复杂,随着退耕年限延长植被会发生演替[2]。根系作为植物直接接触土壤的器官,对土壤环境响应敏感,在植被群落的演替过程中占有十分重要的地位。对不同演替阶段植被根系行为的研究,可揭示不同演替阶段植物群落根系的结构和功能特征的一般变化规律[3-4]。因此,研究植被恢复演替过程中根系内在行为机制及其与土壤环境响应,对于合理加快植被恢复进程,提高水土资源环境利用效率,促进植被建设与生态环境协调发展具有重要意义。
随着根系取样、分析技术的进步,国内外学者利用根表面积、根长密度、根体积密度、比根长等指标,研究植物根系分布特征及其与土壤性状的关系[5-9],取得了一定的研究成果。研究证明:根系长度能反映根系在土体中的稠密度和穿插、缠绕能力,更为直观且易于测量[10-11];根系生物量则反映了生态系统获得能量的能力,为获取足够的养分和水分,植物须维持适量的根系生物量[12]。植物维持根系生物量需投入大量光合产物[13],其消耗的光合产物主要用于吸收土壤养分和水分,而根长密度和比根长又决定根系吸收养分和水分的能力[14]。可见根系生物量与形态指标的非线性耦合技术应用,才是综合反映植被根系与土壤环境响应的关键。
非线性科学为研究生命系统与环境系统之间的关系和作用提供了有效手段[15]。应用分形理论,研究人员对根系的结构特征进行了更加深入细致的研究。研究表明,根系分形维数能够反映出土壤环境变化对根系生长和发育程度的影响[16]。应用根系生态位理论,研究人员掌握了植物群落特性、种间搭配以及生态功能的重要科学。研究表明,合理的植物配置模式有利于退耕地植被根系生态位的恢复,实施退耕还林还草工程是有效增加根系生态位、提高根系多方面功能的重要途径。退耕植物群落根系行为特征的研究中,根系与土壤环境响应关系是根系行为特征总结的重要依据,在有关土壤有机碳研究中,植被根系研究受到了越来越多的关注[17- 18]。黄土高原是世界上水土流失最严重的地区之一,但该地区也是中国的重要农业生产基地之一[19]。因此,采用非线性生态模拟技术,进行黄土区退耕植物群落的演替发展过程中,根系结构与功能恢复状况、相关关系以及对土壤有机碳影响的研究极为迫切。
本文以空间序列代替时间序列的方法,结合黄土区植被演替过程[2]和当地主要植被类型调查结果,选取退耕时间轴中典型的4种优势种群落,调查根系分布格局、组成状况,以基于径级的根系分维数(简称:根长分维数)表征根系结构特征,以根系生态位表征根系功能特征,分析根系行为与土壤有机碳的相关关系。以期了解黄土丘陵区植被恢复过程中,根系内在的行为特征及其与土壤性质的响应关系,为区域生态建设的物种优化筛选、加快生态恢复速度等方面提供理论依据。
1 材料与方法
1.1 研究区概况
研究区位于陕西省绥德县韭园沟乡王茂沟流域,是黄河中游黄土丘陵沟壑区第1副区具有典型代表性的1条流域,是无定河左岸的1条2级支沟,流域面积5.97 km2。自1949年以来,为改善黄土丘陵区严重的水土流失状况,黄委会绥德水土保持科学试验站将王茂沟流域作为试验性治理小流域之一,同时开展沟道淤地坝工程和坡面治理工程。直至90年代,响应国家退耕还林(草)政策,该流域坡耕地陆续弃耕。
流域坐标为110°20′26″—110°22′46″E,37°34′13″—37°36′03″N,海拔高度940—1180 m,属大陆季风性气候,四季分明,多年平均气温10.2℃,多年平均降水量513 mm,多集中在汛期(6—9月),约占全年降水的73.1%。土壤主要以黄绵土为主,覆盖厚度20—30 m,采样坡地全部位于流域主沟道左岸的阳坡中,避免其他景观因子对实验结果产生影响。流域位置与采样点如图1所示。

图1 研究区位置示意图Fig.1 Location of study area
1.2 样品采集
2015年5月下旬进行根系、土壤样品采集。分别选取退耕2 a、8 a、15 a、21 a的坡地作为样地,在每个退耕地内设置2块样地,每块样地设3—5个样方,样方大小均为2 m×2 m,样地调查情况如表1。根据以往研究,黄土区草本类植物根系主要垂直分布于0—50 cm土壤中[20- 21],因此本研究采用内径9 cm根钻分层(10 cm)垂直向下采集至50 cm。从根钻内的土样中挑拣出所有根系,最后将土壤与根系样品分开放入密封袋带回实验室;土壤容重样品采集方法为:在每个根系采样点旁边,挖掘50 cm深的土壤剖面,使用小环刀(100 cm3)分别取0—10 cm、10—20 cm、20—30 cm、30—40 cm、40—50 cm土层的原状土带回实验室。各样地土壤理化性质情况如表2。
1.3 样品分析
1.3.1根系分析
根系样品用清水洗干净后,将其分为4个直径等级:0>D≥0.5 mm,0.5>D≥1 mm,1>D≥2 mm,2>D≥5 mm。用根系扫描仪EPSON TWAIN PRO(32 bit)的16位灰度模式扫描,精度为300 DPI。再使用根系分析软件WinRHIZO分析扫描图片,于是得到各径级的根长、根表面积、根尖数和直径等根系特征参数。最后将根系放入烘箱中,恒温65℃烘干48 h后称重,称量结果精确到0.01 g。

表1 样地调查情况

表2 样地土壤理化性质情况
1.3.2土壤有机碳和容重测定
土壤容重数据由烘干称重法获得,土壤有机碳含量采用德国耶拿公司的有机碳分析仪HT1300固体模块测定。
1.4 数据处理
所有数据采用Excel 2010和SPSS 20.0软件进行分析。利用单因素方差分析(one-way ANOVA)比较不同数据组间的差异。用Pearson相关系数评价不同因子间的相关关系。各土层根系的根长除以对应土壤体积即为根长密度(RLD,Root length density),同样方法获取各土层根系生物量(RWD,Root weight density),两者比值即为比根长 (SRL,Specific root length)[22]。
基于径级的根长分形维数可以反映根系的级配组成,使相同土体单元内不同径级的根系长度得到定量化描述。计算公式[23]如下:
1-(di/dmax)3-D=Wi(δ>di)/Wo
(1)
式中,di表示两根系径级di与di+1间的平均值(di= (di+di+1)/2);dmax为根系最大径级的平均直径;Wi(δ>di)为大于di的积累根系长度;Wo为根系各径级根长的总和。如果以lg(Wi/W0)、lg(di/dmax)分别为横坐标和纵坐标,便可得出3-D是lg(di-dmax)和lg(Wi-Wo)的斜率,D为基于径级的根长分形维数,可通过回归方法求出。
参照胡建忠等[24]提出的方法对根系生态位指数(RENI)进行计算,公式如下:仿照群落生态学研究方法,将群落根系视为“标准地”,分布于每个土体层次的每个径级根系视为“植物种”,根质量、根长、根条数等作为“植物种”的观测变量,最终计算公式为:
(2)
式中,E为群落根系的生态位指数;Wmax、Lmax、Nmax、Smax、Dmax分别为根质量Wij、根长度Lij、根条数Nij、根表面积Sij、根直径Dij指标的最大值。
而层径级根系生态位指数Eij计算公式为:
(3)
式中,Eij为层径级根系生态位指数,这样就可以得到每个土壤或径级根系生态位指数。
上式的意义是将处在三维空间R3的根系分为垂直维:土层Di(i=1、2、3…),即0—10 cm、10—20 cm、20—30 cm、30—40 cm、40—50 cm;水平维:根系径级Rj(j=1、2、3…),即0—0.5 mm,0.5—1 mm,1—2 mm,2—5 mm;指标维:Ik(k=1、2、3…),即根质量W、根长度L、根条数N、根表面积S、根直径D。则有:
Z=f(Di,Rj,Ik)Z∈R3
(4)
式中,Z为等同于植物群落标准地概念不同层次,不同径级的根系参数集合。
2 结果与分析
2.1 不同退耕年限植被根系分布特征
表3显示了4种不同退耕年限植物群落根长密度、根生物量、根系直径和比根长在0—50 cm土壤中的平均值。可以看出根长密度随演替进行先增大后减小,在退耕15 a时,平均根长密度达到最大值31.04 mm/cm3。平均根生物量、根系直在退耕21 a时分别达到最大值3.35 mg/cm3、1.47 mm,其中生物量在群落演替的2—21 a中,逐渐增大。说明植被恢复初期,群落中的主要物种大多为一年生草本植物,其根系较为细长,随着植被演替的进行,多年生草种逐渐占据了主要地位,相应根系生物量逐渐累积,根系直径逐渐变大。平均比根长在群落演替的2—21 a中,逐渐由16.39 mm/mg减小至7.37 mm/mg。比根长是单位根生物量的根系长度,比根长越大则表明根系多以毛细根为主。因此,有学者研究证明,比根长代表了根系收益和花费的关系[25-26]。这也说明,随着植被演替的进行,植物吸收的土壤资源用于根系生物量累积的比率逐渐增大。这可能是,植物群落物种多样性增加,结构、组成和稳定性增强的结果[27]。

表3 根系指标平均值(标准误)
图2显示了根系各指标在土壤剖面的垂直分布特征。各演替阶段植被根长密度均随土壤变深而减小,不同土层之间根长密度存在显著性差异(P<0.05)。
根系生物量的垂直分布特征与根长密度相近,均为随土壤变深而减小,并且在各土层之间存在显著性差异(P<0.05)。
随着土壤深度的增加,根系直径呈现略微降低的趋势。植物群落演替的2—21 a中,根系直径在各土层之间无显著性差异(P>0.05),演替发展至21 a根系直径在各土壤层表出了现显著差异性,说明植物群落演替至21 a时,根系平均直径在土壤垂直梯度中表现出了渐变特征,也可能说明根系的功能性变异。
植物群落演替的2—21 a中,比根长垂直分布呈现出极不规律性,说明各土层的根系均在行使其吸收土壤养分或水分的功能[28]。

图2 不同演替阶段根系垂直分布Fig.2 Vertical distribution of root system in different succession stages同一退耕年限中,具有相同的小写字母表示根系指标在不同土层间差异不显著,显著水平P=0.05
2.2 不同径级根长密度分布
根系的根长密度决定植物吸收土壤水分或养分的能力[29],因此选取不同径级根长密度作为植物根系特征研究的主要对象。由图3可以看出,不同退耕演替阶段中不同径级的根长密度差异较大。从相同土层来看,不同群落根长密度随径级的变化表现出不一致性,其中退耕2 a和8 a的群落中根长密度最大的径级为0>D≥0.5 mm,根长密度随径级增大而减小,原因可能是这两种群落退耕年限较短,群落组成较为简单,进而影响了根长在不同径级的分布。退耕15 a和21 a在表层0—10 cm土层中都均以0.5>D≥1 mm径级的根长密度最大,且表现为偏锋曲线特征。表层0—10 cm土壤中,退耕8 a的铁杆蒿群落差异最大,最大根长密度是最小值的1.86倍,其他依次为 2 a(1.57倍)、21 a(1.36倍)、15 a(1.27倍)。
随着土壤变深,不同径级的根长密度逐渐减小。第1径级(0>D≥0.5 mm)根系:表层0—10 cm土壤中根长密度分别是底层(40—50 cm)1.80—5.26倍,退耕21 a最小(1.80倍),退耕15 a最大(5.26倍),此外,第1径级(0>D≥0.5 mm)在表层和底层间的差异相比其余径级最小;第2径级(0.5>D≥1 mm)根系:表层土壤根长密度是底层的0.85—46.05倍;第3径级(1>D≥2 mm)根系:表层土壤根长密度是底层的0.97—16.68倍;第4径级(2>D≥5 mm)根系:表层土壤根长密度是底层的0.54—87.38倍。表明退耕植被群落中,径级越大土壤表层与底层的根长密度差异越大,径级越小则的差异越小。

图3 不同径级根长密度的垂直分布Fig.3 Vertical distribution of root length density at different diameter levels
2.3 不同退耕年限根系分维数、生态位指数
根系分形结构是植物根系构型应对环境异质性的表型可塑性结果,可反映植物对生长环境的适应策略[30]。受遗传特性、群落环境、土壤空间异质性等诸多因素的影响,植物群落在生存和发展过程中会根据自身立地条件,适时调整、转换或补偿自身功能,以最优策略完成“生存、生长、繁殖”的三大目的,最终改善了根系结构及其生理性状的生态适应性。因此将分形理论用于根系研究,可以提高定量描述根系结构的可靠性,从生态和生理角度分析根系构型特征[31]。基于径级的分形维数反映了退耕群落各径级根系的结构状态,分形维数越大,说明较细的根系占比例越多,即根系结构较为单一。分形维数越小,则表明根系在不同径级分布的越均匀,即根系结构较为完整和复杂。
退耕群落根系在各土层的分形特征如图4所示,可以看出,不同退耕群落根长分维数介于2.36—2.93之间。退耕演替发展的2—21 a中,植物群落根系分维数呈显著减小性趋势,这充分说明了随着植被演替的发展,各径级根长均占有一定比例,即根系的结构完整性和复杂程度逐渐增加。
退耕2 a和8 a的群落在0—50 cm土层的分维数均最大(P<0.05),表明这2种群落不同径级根系的分布较不均匀,主要以径级为0>D≥0.5 mm的根系为主。值得指出的是,这2种群落根长分维数在0—20 cm土层中较小,仅在20—50 cm土层均较大。说明根系在0—20 cm土层的分布相对均匀,而在20 cm以下的土壤中,随着土壤深度增加根系结构迅速单一化,径级较小的根系迅速增多。这类现象与2种群落退耕年限较短,各类草种根系向深层生长的能力较弱有关。在0—10 cm土层,退耕21 a的根长分维数最小(2.45),说明不同径级根系长度分布均匀,这主要是由于灌木群落退耕年限较长,群落植物种丰富、覆盖度较高的影响。
不同群落根系生态位指数可以反映根系在不同土层的生态功能状况,生态位越大,说明根系的功能性越强,在所处土壤环境的竞争能力越强。如图4所示,退耕演替发展的2—21 a中,植物群落根系生态位指数呈显著增大性趋势,这充分说明了随着植被演替的发展,根系的生态功能逐渐增加。
不同退耕群落的生态位指数在各土层的变化规律相似,均以表层最大,并且随土壤深度增加而减小。结合根系分维数所表现的结构特征,说明根系的结构越复杂,其生态位指数越大,即功能性越强。这一结果可以说明根系结构的复杂程度直接决定了根系功能性的强弱。

图4 根长分维数与根系生态位指数Fig.4 Root length dimension and root ecologic index具有相同的大写字母表示分维数或生态位指数在不同演替阶段间的差异不显著,显著水平P=0.05
2.4 根系行为特征与土壤性质相关关系
退耕植物群落土壤有机碳含量与容重在各土层的分布特征如图5所示,可以看出,不同退耕群落土壤有机碳含量在3.72—8.39 g/kg之间。退耕演替发展的2—21 a中,土壤有机碳含量存在一定的增大趋势,说明随着植被演替的发展,土壤有机碳含量存在缓慢的恢复现象,而方差检验下的增大趋势并不显著(P>0.05),这说明在黄土区植被自然恢复的21 a内,土壤有机碳含量还未发生较为显著的改善。土壤容重随群落演替的发展也未发生明显改变,随土壤深度的变化不显著,说明植被自然恢复的21 a中,未对土壤容重产生较大影响。
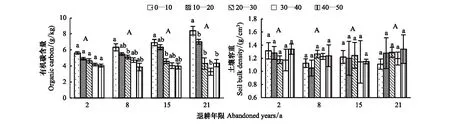
图5 退耕地土壤有机碳含量与容重分布Fig.5 Distribution of soil organic carbon content and bulk density in abandoned land同一退耕年限中,具有相同的小写字母表示有机碳或容重在不同土层间的差异不显著;具有相同的大写字母表示在不同演替阶段间的差异不显著,显著水平P=0.05
采用 Pearson 典型相关分析研究土壤有机碳、容重、根长密度、分维数、根系生态位指数以及不同径级根长所占比例的关系(表4)。可以看出,土壤容重与其余任何因子均不存在显著相关性(P>0.05),说明退耕植物群落演替发展的2—21 a中,土壤容重不受植物根系的影响。
土壤有机碳与根长密度、生态位指数、P0.5>D≥1mm、P1>D≥2mm呈极显著或显著正相关(P<0.01)或(P<0.05),相关系数分别为0.803、0.742、0.614、0.477。土壤有机碳含量与根系生态位指数呈极显著正相关说明基于统计学和生态学理论,结合多种根系特征参数得出的根系生态位指数,对土壤有机碳的富集起到极显著作用,从而证明根系丰富土壤有机碳的过程是根系综合生态功能性作用的体现。
根长分维数与0>D≥0.5 mm根长所占比例极显著正相关(r=0.989,P<0.01),表明分形维数越大,根系长度越集中分布于0—0.5 mm径级内。分维数与径级0.5>D≥1 mm、1>D≥2 mm、2>D≥5 mm根长所占比例呈极显著负相关(P<0.01),表明分形维数越小,中等或较大径级的根长密度越大,从而根长在不同径级内的分配趋向于均匀化,即根系结构的完整性增强。分维数与根系生态位指数呈极显著负相关(P<0.01),这充分的说明分维数越大根系分布越均匀,即物种越丰富、根系结构越复杂,从而群落根系竞争力和功能性越强。因此,根长分维数与根系生态位指数呈极显著负相关的逻辑性成立。

表4 根系的结构、功能特征与土壤理化性质的相关系数
* 表示在 0.05 水平(双侧)上显著相关,**表示在 0.01 水平(双侧)上显著相关;P0>D≥0.5mm:直径介于0—0.5 mm间的根长占总根长的百分比;P0.5>D≥1mm、P1>D≥2mm、P2>D≥5mm依次类推
3 讨论与结论
黄土高原地区大面积的“退耕”工程,使该区域陆续出现了大量的弃耕坡地。又由于黄土区显著的季风气候,新生植物群落对有效土壤资源的获取能力决定了它们的竞争力和生产力[21]。因此,植物群落发展出不同的根系分布、生长策略来应对高时空异质性的土壤水分和养分环境。从生态学的角度来看,前人的研究焦点是探讨使植物适应这些极端气候或环境变化的机制[32],并确定哪些根系指标是植物吸收土壤水分、养分的最大贡献者。大多数研究者认为毛细根或拥有较大表面积比的细根可以有效提高获取土壤资源的能力,而有些学者则认为粗根或老根则可以更好的保持植物取得的土壤水分和养分[33]。显然,在根系获取与保持土壤资源间存在一个临界点。Riva等[34]就临界点问题开展了研究,在80多种植物根系的分析后发现根系形态特征指标与预期不同,其中根系生物量、根系直径的分析结果与根长、根面积等存在显著差异,进一步说明了根系特征多重维度变异的可能性。基于径级的根系分形维数很好的将根系结构特征量化,可以利用不同植物群落根系的分维数来横向对比。随着演替进行,植物群落逐渐走向稳定,其地下根系生态系统应逐渐趋于复杂化,而不是以结构单一的细根为主,即以提高获取土壤资源利用率为主要目的群体。因此本文分析结果中,基于径级的根系分形维数随植被演替进行逐渐减小顺理成章。
根系生态位的理论则是针对上述根系多重维度变异情况提出了一种多维度根系计算方法。其原理是将本研究所有的植物群落看作一个生态系统,并选择这一生态系统中各根系指标的最大值,在多维度空间进行非线性耦合,即垂直维度、水平维度和指标维度。将这三个维度的生态位指数之和,定义为某一植物群落根系生态位指数,随后便可利用不同植物群落根系生态位指数来横向对比。根系生态位指数是对根系综合生态功能特征描述的一种新型方法,综合反映了作为客体生态环境空间的垂直分布、作为主体的生物根系径级分布以及根系各种指标分布的三方面特征。
综上所述,在根系生态学研究中,一个复杂性难题是如何通过根系结构特征反映根系的生态功能情况,即根系对土壤界面相互作用、资源获取等行为特征问题[35- 36]。然而,这一问题在不同的植物群落、不同气候特征、不同土壤立地条件下有很大差异,这就使得能够真实反映地下根系生态系统的研究尤为复杂和困难。本文提出基于径级的根系分形维数和根系生态位指数,较为科学、客观,并具有生态学和统计学理论作为基础,在短时期根系的研究中或受控的研究条件下运用,较为可行且可操作性强。最终,通过对退耕植物群落根系及土壤的有机碳的研究得出主要结论为:根长密度、根生物量、根系直径随着退耕演替进行缓慢增大,比根长则持续减小。退耕演替发展的2—21 a中,基于径级的根系分维数由2.77显著减小至2.59(P<0.05),即根系结构在植被恢复的过程中逐渐趋于完整。根系生态位指数则较好的量化了根系的功能性,2—21 a中,生态位指数由3.75显著增大至9.37(P<0.05)。根系生态位指数与土壤有机碳含量极显著正相关(P<0.01),即根系功能性的恢复可以显著的提高土壤有机碳含量。基于径级的根系分维数与生态位指数呈极显著负相关(P<0.01),即根系的结构特征直接决定了根系综合生态功能,表现为结构越复杂,功能性越强。
猜你喜欢
广西林业科学(2022年4期)2022-11-07
林业调查规划(2022年3期)2022-06-13
小小说月刊·下半月(2021年8期)2021-09-05
园艺与种苗(2021年3期)2021-05-26
世界地质(2021年1期)2021-04-07
中国农村水利水电(2020年5期)2020-06-15
水利规划与设计(2020年1期)2020-05-25
动漫界·幼教365(大班)(2018年3期)2018-05-14
湖南大学学报·自然科学版(2016年12期)2017-05-12
江苏农业科学(2016年4期)2016-06-14
