
长汀水土流失区侵蚀劣地马尾松种群动态
2019-05-13 02:06吴语嫣李守中孙眭涛杨贤宇黄思彤
生态学报 2019年6期
吴语嫣,李守中,*,孙眭涛,杨贤宇,黄思彤,王 凯
1 福建师范大学湿润亚热带山地生态国家重点实验室培育基地,福州 3500072 福建师范大学地理科学学院,福州 3500073 地理学国家级实验教学示范中心(福建师范大学),福州 350007
植物种群动态是植物种群生态学研究的核心问题之一[1- 2],主要研究生存在特定环境空间里的植物种群随时间呈现的个体数量消长和分布变迁规律[1]。植物种群内部个体的生命过程(出生、成长、发育、繁殖和死亡)由物种生物学特性与生境条件共同作用的结果来决定[3],不同生活史阶段的个体呈现不同生命率,会直接影响植物种群数量动态及其发展趋势[4]。种群统计是种群动态研究的基础,通过统计调查能够迅速获取种群的出生、死亡、增殖以及年龄结构等直观数据,研究生活史各阶段的种群数量变化特征及其变化原因[5]。目前年龄结构分析、静态生命表、存活曲线、生存分析和时间序列预测是植物种群动态研究中普遍应用的重要统计方法[6],应用到灌木、长寿命乔木和濒危、稀有树种的很多研究中[7- 13],揭示了植物种群生物学特性与环境间的关系和种群在群落中的地位和作用[14- 18],为种群的管理和开发利用提供理论指导[2,14- 15,17,19- 20]。
马尾松(Pinusmassoniana),松科松属,适生能力强、分布范围广、生长迅速,是我国亚热带红壤水土流失区生态恢复与重建的乡土树种和先锋树种[21- 22]。福建省长汀县是我国典型的花岗岩红壤侵蚀区,马尾松次生林是当地最主要的森林类型,马尾松种群成为植被组成中的重要部分。另一方面,由于亚热带红壤生态系统的潜在脆弱性[23],该地区降雨侵蚀力大、地表径流冲刷强、地形破碎、地表水热变化剧烈、土壤持水性差且肥力极低[24- 25],生长于侵蚀劣地的马尾松面临极端环境因素带来的生存胁迫,常常形成生长受抑制、稀疏矮小的“小老头松”林。作为开展水土流失后续治理措施的关键前提,整体把握侵蚀劣地的马尾松种群生存现状和发展状况十分必要。
本研究以空间代替时间,以径级结构代替年龄结构,依据某一特定时间断面不同年龄阶段的植株数量配置建立静态生命表,应用生存分析理论和时间序列预测模型,分析侵蚀退化生境马尾松种群的存活状态和数量变化规律,探究其生物学特性与环境间的适应度,并根据种群发展的薄弱环节,提出相应的抚育措施,以期为长汀水土流失区侵蚀劣地“小老头松林”的管理与改造提供重要依据。
1 研究区概况
研究区位于福建省长汀县河田镇(25°30′—25°44′N,116°16′—116°34′E),地处武夷山脉南麓,地形以低山、丘陵为主。属中亚热带季风气候,年平均气温18.3℃,年均降水量1700 mm,降雨主要集中在4—7月,季节性干旱常见于7—8月。土壤类型为抗蚀性差且酸性强的山地红壤,保水保肥能力低。由于历史上长期以来的人类掠夺式开发,原生植被几乎消失殆尽[26],现存马尾松次生林多为20世纪80年代经过飞播、封育和改造措施形成。而“小老头松”林则主要分布于环境恶劣的花岗岩侵蚀坡地[27],长期封禁下的植被稀疏、自我恢复能力弱,乔木层仅以马尾松为主,灌木层有稀少的木荷(Schimasuperba)、黄瑞木(Adinandramillettii)、油茶(Camelliaoleifera)和胡枝子(Lespedezabicolor)、草本植物有芒萁(Dicranopterisdichotoma)、芒草(Miscanthussinensis)等。
2 研究方法
2.1 样地设置与种群调查
综合考虑土壤、坡度、海拔、人为干扰、植被覆盖度等指标,选择了长汀县河田镇伯公岭和伯湖村的“小老头松”林为研究对象,共设置4个30 m×30 m样地。采用相邻网格法将每个样地划分成36个5 m×5 m小样方,对样方内马尾松进行每木调查,记录每株个体的基径、树高和冠幅等指标,同时测定并记录样地的生境信息,样地基本情况见表1。
B1:伯湖村1样地; B2:伯湖村2样地; B3:伯公岭3样地; B4:伯公岭4样地
2.2 年龄结构的确定和量化分析方法
由于实际工作中无法追踪马尾松所有个体的生长周期,相关研究认为同一树种的龄级和径级对环境的反应规律具有一致[28],因此本文采用空间推时间,径级结构代替年龄结构的方法分析种群结构动态[16,18,29- 30]。基于样地调查数据和马尾松生活史特点,将植株划分为幼龄期、壮龄期和成熟期三个生长阶段。具体龄级划分如下:新生苗设置为Ⅰ级;树高≤25 cm,基径≤20 mm时为Ⅱ级;25 cm<树高≤50 cm,基径≤40 mm时为Ⅲ级;树高>50 cm,基径按20 mm划分为1个龄级,40 mm<Ⅳ≤60 mm,……,Ⅸ>140 mm。共划分9个龄级,其中Ⅰ—Ⅲ龄分别表示繁殖前期的新生苗、幼苗、幼树,Ⅳ—Ⅵ龄表示具有繁殖能力的壮龄个体,Ⅶ—Ⅸ龄表示成熟个体。

(1)
(2)
(3)
(4)

2.3 静态生命表、存活曲线与生存分析
建立生命表能直观反映种群从出生到死亡的动态过程,是种群统计的核心[32]。编制静态生命表的方法[33]如下:存活数ax为径级x内出现的个体数,标准化存活量lx=ax/a0×1000,标准化存活量的自然对数值为lnlx,死亡量dx=lx-lx +1,死亡率qx=dx/lx,区间寿命Lx=(lx+lx +1)/2,总寿命Tx=ΣLx,期望寿命ex=Tx/lx,消失率Kx= lnlx-lnlx +1。
存活曲线是对生命表的重要反映[15],本文以存活量lx的对数值为纵坐标,以龄级为横坐标作出马尾松种群存活曲线。
生存分析采用4个函数即生存函数S(t)、累积死亡率函数F(t)、死亡密度函数f(t)、危险率函数λ(t),用于分析马尾松种群动态[34- 35]。公式如下:
S(t)=S1·S2·S3·…·Si
(5)
F(t)=1-S(t)
(6)
f(t)=[S(t-1)-S(t)]/hi
(7)
λ(t)=2(1-Si)/[hi(1+Si)]
(8)
式中,Si为存活率,hi为龄级宽度。
2.4 时间序列预测
采用线性平稳时间序列分析中的一阶移动平均模型对马尾松种群年龄结构动态进行预测,具体算法如下[9]:
(9)

3 结果与分析
3.1 马尾松种群年龄结构
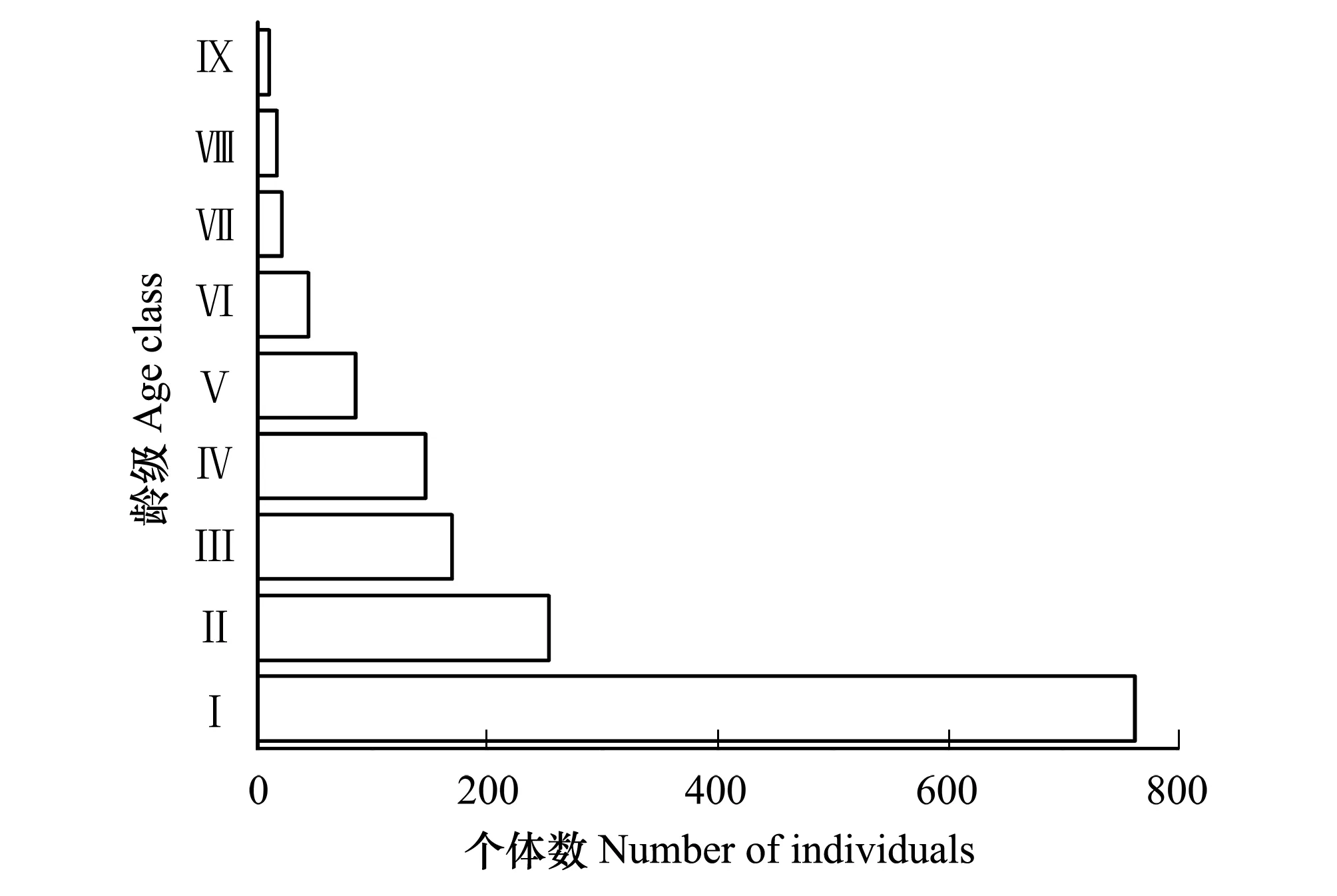
图1 马尾松种群年龄结构图 Fig.1 Age structure of Pinus massoniana population in plotⅠ:新生苗;Ⅱ:H(树高,height of individuals)≤25 cm,BD(基径,basal diameter of individuals)≤20 mm; Ⅲ:25 cm
由图1可知,幼龄期(Ⅰ—Ⅲ龄)植株数量丰富,占种群的78.58%。Ⅰ龄个体超过种群总数量一半,使马尾松年龄椎体呈现宽基底;Ⅱ龄数量是Ⅰ龄的33.16%,Ⅲ龄数量是Ⅱ龄的66.80%,表明幼龄阶段个体续存率低。壮龄期(Ⅳ—Ⅵ龄)植株占种群比例的18.24%,年龄锥体横柱从下往上缓慢变窄。成熟期(Ⅶ—Ⅸ龄)植株只占种群年龄比例的3.18%,个体数量变化不大。因此,马尾松不同龄级个体数量分布呈基部极宽顶部狭窄的金字塔形。

表2 马尾松种群龄级结构动态变化指数
3.2 马尾松种群静态生命表和存活曲线
由马尾松静态生命表看出(表3),实际存活数(ax)逐渐减少,死亡出现在种群各个生活史阶段;反映个体死亡概率的特定年龄死亡率(qx)在Ⅰ龄级最大,幼龄期(Ⅰ—Ⅲ龄)逐渐减小,壮龄期(Ⅳ—Ⅵ龄)逐渐变大,成熟期(Ⅳ—Ⅵ龄)波动变化,反映个体平均生存能力的生命期望值(ex)在Ⅱ龄级最大,Ⅱ—Ⅴ龄级逐渐下降,后期Ⅴ—Ⅶ龄级略有升高,之后再次下降,说明种群在死亡过程中,不同生活史个体的适应力差异显著;消失率kx反映了个体对总死亡效应的贡献,幼龄期、壮龄期和成熟期个体分别占总死亡效应的38.11%、44.80%和17.09%,说明壮龄期是死亡的重要阶段;新生苗占总死亡效应的25.40%,新生苗是死亡的主体,对死亡贡献最大。
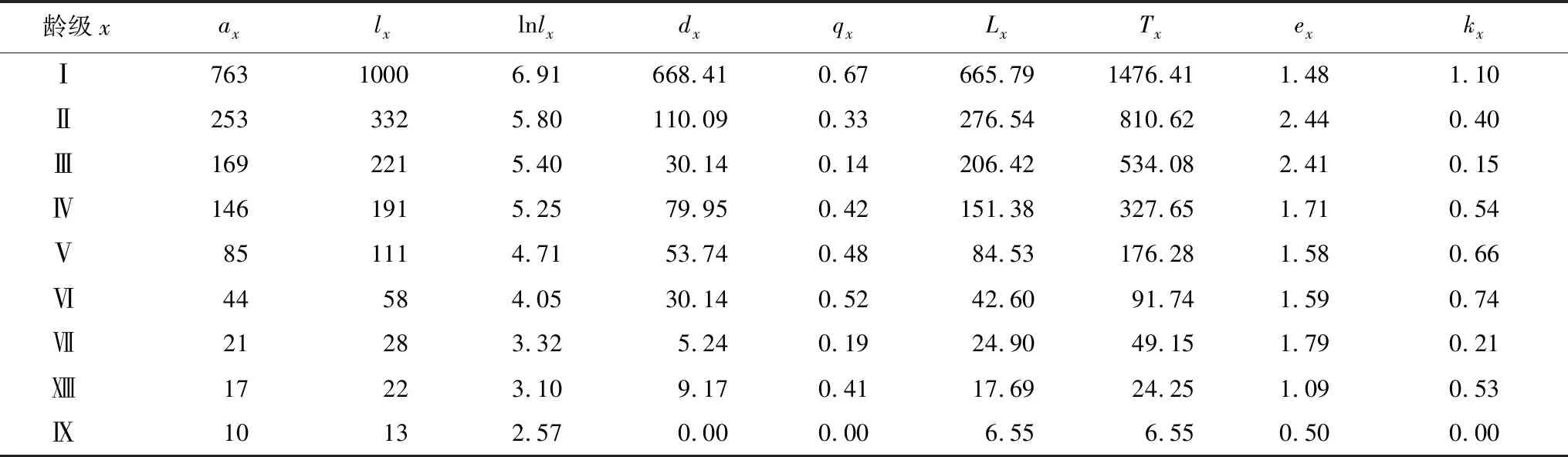
表3 马尾松种群的静态生命表
x:龄级 Age class;ax:存活数 Survival number;lx:存活量 Survivors;dx:死亡量 Death;qx:死亡率 Mortality rate;Lx:区间寿命 Span life;ex:期望寿命 Life expectancy;kx:消失率Vanish rate

图2 马尾松种群存活曲线Fig.2 Survive curve of Pinus massoniana population
存活曲线按Deevey的划分一般有3种基本类型[36]:Ⅰ型是凸曲线,表示幼体存活率高,老年个体死亡率高,在接近生理寿命前只有少数个体死亡;Ⅱ型是对角线,表示整个生活期中的死亡率较稳定;Ⅲ型是凹曲线,表示幼体死亡率高,以后的死亡率低而稳定。由图2可看出,马尾松种群存活曲线早期呈现“凹凸凸”的变化趋势。Hett和Loucks检验估算存活状况符合Deevey-Ⅱ型存活曲线还是Deevy-Ⅲ型存活曲线时,分别采用指数方程式Nx=N0e-bx和幂函数式Nx=N0x-b这两种数学模型[37]。通过模型拟合的决定系数与Deevey-Ⅱ型有更高的相关性(表4),判断得出:马尾松种群存活曲线属于Deevey-Ⅱ型,即种群总体上死亡率相近且相对稳定。

表4 存活曲线的拟合方程
3.3 马尾松种群生存分析
由图3可知,随着龄级的增长,马尾松种群的生存函数S(t)单调下降,累积死亡率函数F(t)单调上升,且两条曲线的斜率在Ⅵ龄级前变化明显,在Ⅵ龄级后平缓,说明种群在幼龄期和壮龄期生存状况不稳定,成熟期适应生境的生存能力提高。
由图4可知,马尾松种群死亡密度函数f(t)与危险率函数λ(t)变化趋势一致,都在Ⅰ—Ⅱ龄急剧下降,Ⅱ龄级后平缓波动,Ⅰ龄级处于死亡密度函数和危险率函数最高峰值,壮龄期(Ⅳ—Ⅵ龄)的死亡密度都较大,说明新生苗和壮龄个体是马尾松种群增长过程中的脆弱部分。

图3 生存率S(t)和累积死亡率F(t)函数曲线Fig.3 Survival rate S(t) and cumulative mortality rate F(t) functional curves

图4 死亡密度函数f(t)和危险率函数λ(t)曲线Fig.4 Mortality density f(t) and hazard rate λ(t) functional curves
3.4 马尾松种群数量的时间序列分析
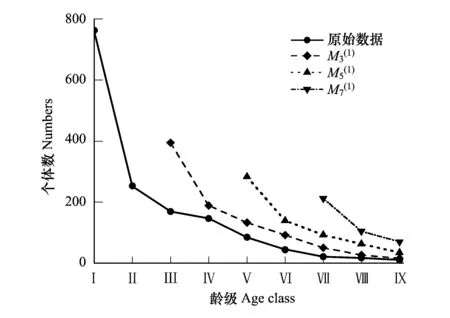
图5 马尾松种群数量动态时间序列预测 Fig.5 Time series prediction of Pinus massoniana population经过3、5、7龄级时间后的种群大小;(1):移动平均法中的一次移动平均法标识
基于马尾松种群的统计数据,利用时间序列预测模型,对经过3、5、7龄时间后种群各龄级内的个体数量进行预测。可看出(图5),随时间推移,各龄级的个体数量均呈现增加趋势,如经过3、5、7龄级时间后,第Ⅶ龄级个体数量分别增加了138.10%、86.00%和127.96%,Ⅷ龄级个体数量分别增加了58.82%、133.33%和66.67%,表明未来马尾松种群呈增长趋势。随着成年个体的增多必然会加剧侵蚀退化贫瘠生境中的土壤养分和水分争夺,抑制个体生长,形成更多“小老头松”。
4 讨论
4.1 马尾松种群结构特征与适宜性
种群年龄结构是植物对外界环境适应性的体现,反映林木更新过程中经受的重大干扰事件[38]。调查结果显示,样地内马尾松种群结构呈基部极宽顶部狭窄的金字塔形,属于增长种群但增长性低。马尾松天然更新能力强,在自然干旱环境的长期选择下,种子已经具有适应水分胁迫机制[39],能在贫瘠生境内萌发并建成幼苗;样地内林木稀疏,植被覆盖度低,尤其春季光热充足、降水丰富,有利于马尾松土壤种子吸水萌发。通过野外调查发现种群首年生产松果数达734颗,次年新生幼苗数达763株,一定程度上说明了马尾松种源充足,能以有性繁殖保证新生苗的数量,因此马尾松的生物学特性是新生苗建立的关键。
建成限制是制约更新的重要机制[40],年龄结构中只有约33.16%的新生幼苗成长至幼苗阶段,表明样地环境筛的强烈过滤作用是新生苗定居困难的重要因素。该地降水频繁,土壤侵蚀严重,基岩裸露,养分含量极低,尤其夏季暴雨后光照辐射强烈,干热化程度异常,地温最高达60℃,新生苗的定居直接受极端生境因子的影响,植株数量大幅度减少。幼苗是种群天然更新的瓶颈[41],幼苗根系浅,个体小,生理生态发育不完全[21],在频繁遭受生境带来的土壤掩埋、干热胁迫、养分不足的限制后,个体数量继续减少,约66.80%的幼苗发育成幼树。因此,严酷的生存环境严重降低了幼龄个体的续存率。
幼龄个体数量为种群更新发育提供物质基础,维持种群稳定性[7],种群中的成年个体占种群总量的21.42%,其中,9.68%的个体来源于幼龄的补充,所以低续存下的幼龄仍能进入种群更替层,补充成年个体,促进种群更新。
4.2 马尾松种群动态特征和发展趋势
植物种群生命表和存活曲线的分析可以推断种群时间上的动态,反映种群与环境的竞争关系[42]。本研究中,种群不同生活史阶段的死亡率和生存能力差异显著,Ⅰ龄是种群死亡的最大贡献者,这一时期的新生苗需要争夺较多土壤水分以满足生存,同时抵抗外界环境变化的能力最为脆弱[43],因此新生苗阶段以高死亡率为代价应对个体间竞争和生境强烈胁迫效应[16];幼苗阶段生命质量值最高,说明经历前期的环境筛选作用后,植株抗性增强,生理活动旺盛;马尾松从幼树成长为大树的过程中,由于植株进入营养生长和生殖生长,对养分、水分、光照等资源需求增加,有限的生境资源导致种群内部资源竞争激烈自疏作用增强,死亡个体逐渐增多,种群在整个壮龄期的死亡率和消失率持续升高、生存能力下降,在Ⅵ龄迎来死亡率次高峰,Ⅵ龄个体处于壮龄后期,生理生态特征发育成熟,抵御生境胁迫的能力提高,生命期望值略有上升;种内竞争的加剧导致进入成熟期的个体数量锐减,内部生存资源的竞争压力减小,加上Ⅶ龄个体适应生境的能力提升,此时的死亡率下降,生命期望值提高;研究表明,植物生长发育过程中,生长、繁殖、防御等机能间存在资源分配的权衡,某一过程的投入增加要以减少另一过程投入为代价[44- 45],成熟个体后期挤向主林层的过程中需要最优化权衡分配资源来适应环境的变化,马尾松成熟个体是产生种子的主要来源[21],繁殖生命过程需要植株配置大量资源,那么抵御复杂生境因子的资源投入和生长资源投入相对减少,植株生长十分不利,死亡率上升,生命力下降。
尽管不同阶段马尾松种群的生存和死亡波动较大,但马尾松存活曲线呈现Deevey-Ⅱ型,表明种群总体上处于动态稳定状态。已有的研究表明,光源竞争导致的数量消长是引起马尾松种群结构动态变化的主导因子,在生境条件严酷的马尾松疏林地,光源充足,马尾松能够自我维持[46]。时间序列分析预测,未来 3、5、7龄级时间后马尾松壮龄和成熟龄个体逐渐增多,说明了马尾松作为强阳性树种,环境适应力强,种群更新良好,能适应侵蚀劣地生境,体现了马尾松作为该地区生态恢复与重建树种的优越性。
4.3 侵蚀劣地马尾松种群抚育与经营措施
以上分析结果表明,侵蚀劣地马尾松林当前阶段幼龄个体众多,上层木少且普遍低矮,水土流失区的反复侵蚀,地形破碎陡峭、土壤持水保肥能力差、干热化严重等很容易加重幼龄个体的低续存率问题。未来随着更多成年植株进入林冠层,土壤旱瘠矛盾会使壮龄个体间资源竞争更为突出,加剧成熟龄个体繁殖和生长资源分配失衡,导致更多成年马尾松衍化成生长受限繁殖能力不足的“小老头松”,难以发挥亚热带森林应有的生态效益。
针对马尾松种群不同阶段的生长发育特点采取抚育措施是“小老头松”林可持续发展的关键。由于生境胁迫和种内竞争的共同作用,种群在幼龄期和壮龄期生存很不稳定,采取人工措施对其干预有一定合理性。短期内,草本植物的覆盖有利于改善近地面小气候,防治水土流失,建议种草提高植被覆盖度来减轻水土流失对小个体带来的生存压力和小个体间的养分争夺。对于既要应对个体间资源竞争又要权衡繁殖和生长的资源分配的成年个体,应施肥促进其生长和繁殖质量。
猜你喜欢
农业灾害研究(2022年6期)2022-12-02
湖南生态科学学报(2022年4期)2022-11-11
热带作物学报(2022年9期)2022-10-17
农业科技与信息(2022年12期)2022-08-09
河南农业大学学报(2020年2期)2020-05-22
绿色科技(2019年5期)2019-11-29
山西林业科技(2019年1期)2019-05-27
中国粮油学报(2018年12期)2018-03-19
中国果菜(2016年9期)2016-03-01
飞言情A(2014年12期)2014-12-17
