
Impacts of Sletr1-1 and Sletr1-2 mutations on the hybrid seed quality of tomatoes
2019-05-10 06:14:20SyarifulMubarokHiroshiEzuraAnasKusumiyatiNeniRostiniErniSuminarGungunWiguna
Syariful Mubarok, Hiroshi Ezura, Anas, Kusumiyati, Neni Rostini, Erni Suminar, Gungun Wiguna
1 Department of Agronomy, Universitas Padjadjaran, Sumedang 45363, Indonesia
2 Tsukuba Plant Innovation Research Center, University of Tsukuba, Tsukuba 305-8572, Japan
3 Graduate Program of Plant Science, Faculty of Agriculture, Universitas Padjadjaran, Sumedang 45363, Indonesia
4 Vegetable Crops Research Institute, Ministry of Agriculture, Bandung 40391, Indonesia
Abstract Shelf life is an important breeding trait in tomato, especially for the tomato production in subtropical and tropical regions. Previously we have isolated and characterized ethylene receptor mutants, Sletr1-1 and Sletr1-2 from mutant population based on Micro-Tom cultivar. Sletr1-1 showed insensitivity to ethylene while Sletr1-2 showed reduced sensitivity to ethylene. We also have demonstrated that the traits are useful for extending fruit shelf life of the hybrid tomato cultivars. For commercializing the hybrid cultivars, the seed quality is another important trait. In this study, we evaluated the effects of the Sletr1-1 and Sletr1-2 mutations on the seed quality characteristics of F1 hybrid lines generated by crossing Sletr1-1 and Sletr1-2 with three commercial tomato cultivars, Intan, Mutiara and Ratna. Sletr1-1 mutation conferred insensitivity to ethylene in the F1 hybrid seedlings, resulting in negative effects including reduced germination rate, vigor index and emergence speed index. Interestingly Sletr1-2 mutation had almost no effect on the seed quality characteristics of the F1 hybrid lines, suggesting that Sletr1-2 was suitable for producing high quality of hybrid seeds.
Keywords: ethylene receptor, mutant, hybrid quality seed, tomato
1. Introduction
Tomato (Solanum lycopersicum L.) is one of the important horticultural crops in the tropics and subtropics, because it contains high levels of micro- and macro-nutrients such as vitamins, minerals, f iber and other compounds desirable for human health. Since one of the major concerns in tomato production is a postharvest loss of fruits, considerable efforts have been made on breeding new tomato cultivars conferring long shelf life for the fruits. Both natural and induced mutants giving extended fruit shelf life were used in the breeding programs. Several genetic mutants affecting tomato ripening, such as ripening-inhibitor (rin), Colorless non-ripening (Cnr), non-ripening (nor), Never-ripe (Nr), Sletr1-1 and Sletr1-2 have been isolated and characterized (Lanahan et al. 1994; Wilkinson et al. 1995; Vrebalov et al. 2002; Okabe et al. 2011). Sletr1-1 and Sletr1-2 mutations could signif icantly extend the shelf life of tomato fruits, and Sletr1-2 mutation especially conferred prolonged fruit shelf life without a reduction in carotenoid content (Okabe et al. 2011). Subsequently, we demonstrated the use of Sletr1-1 and Sletr1-2 mutations in hybrid tomato breeding; Sletr1-2 mutation can extend fruit self life in the F1hybrid lines with minimal effects on the agronomic traits (Mubarok et al. 2015, 2016).
Seed quality is another important breeding trait that is important for commercializing hybrid seeds. Since the 1920s, many studies have shown that ethylene regulated seed quality by affecting the seed germination (Egley 1982). At present, it is an established fact that in many species, the application of ethylene or the ethylene-releasing compound, breaks primary dormancy, secondary dormancy in seeds and accelerates the germination of non-dormant seeds (Kepczynski 1985; Esashi 1991). Other studies have shown that ethylene is involved in enhancing seed dormancy, vigor and the time required for radicle emergence as a measure of seed vigor (AOSA 1983; Khan 1994). The evidence of the reduction of seed quality in an ethylene-insensitive mutant was observed in the Nr mutant that affects seed vigor (Siriwitayawan 2003). Numerous studies showed that a reduced ethylene production in chickpea, sunf lower and lettuce is associated with the induction of thermodormancy (Corbineau et al. 1988; Prusinski and Khan 1990; Gallardo et al. 1991). Therefore, when using ripening-related mutants in a breeding program selecting for long shelf life, the effects of the mutations on seed quality characteristics should be carefully evaluated.
Consequently, when generating new F1hybrid cultivars with a long shelf life using the Sletr1-1 and Sletr1-2 mutations, the effects of these mutations should be clearly evaluated. In this study, we evaluated the effects of the Sletr1-1 and Sletr1-2 mutations on the seed quality characteristics of F1hybrid lines generated by crossing Sletr1-1 and Sletr1-2 with three commercial tomato cultivars, Intan, Mutiara and Ratna.
2. Materials and methods
2.1. Plant materials and growth conditions
Sletr1-1 and Sletr1-2 are the mutants, which isolated and characterized from a mutant population based on Micro-Tom cultivars (Okabe et al. 2011). Sletr1-1 showed insensitivity to ethylene whereas Sletr1-2 showed reduced sensitivity to ethylene. The varieties Intan, Mutiara, and Ratna are the commercial pure-line cultivars, which were generated by the Vegetable Crops Research Institute, Indonesia. Those three cultivars are beef tomatoes that are adaptive for the Indonesian climate. Those plants were cultivated under normal condition using a combination of growing media of soil+compost+peat moss (1:1:1, v/v) in the greenhouse from August to November 2017, with a day temperature of 33°C and night temperature of 22°C. The hybrid seeds were generated by crossing these mutants, Sletr1-1 and Sletr1-2, and the wild-type Micro-Tom (WT-MT) as a pollen donor, with the three pure-line cultivars, Intan, Mutiara and Ratna, as a pollen recipient. The hybrid F1seeds were harvested from the red fruit of the cross between the ethylene receptor mutants and pure-lines cultivars. Harvested seeds were rinsed with water to remove the gel around the seed, and then they were dried at room temperature. After that, the dried seeds were kept in storage at 4°C.
2.2. Seed quality analysis
To analyze the seed quality of mutant and hybrid lines, several parameters including seed germination, emergence speed index and vigor index have been investigated. Seed analysis was conducted based on the ISTA analysis protocol for seed testing (ISTA 2017). In brief, 400 seeds were germinated in wet paper using the top paper method and this was repeated four times. They were distinguished and counted for normal seed characterized by the root and shoot being well developed and abnormal seed characterized by the root or shoot being not well grown, the structure being deformed or disproportionate and otherwise improperly developed. Germination rate was counted based on the percentage of normal seed at 5 and at 7 days after showing (DAS), respectively, using the following equation:
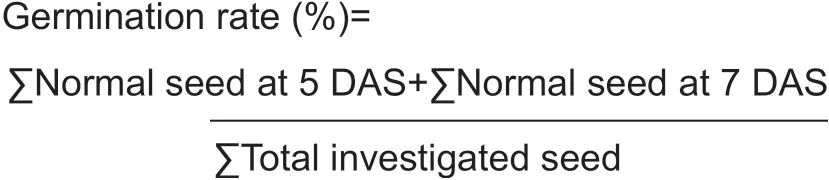
Vigor index was calculated as a percentage of normal seed that was counted at 5 DAS, whereas emergence speed index was counted daily for 14 days and calculated according to Maguire (1962).
2.3. Triple response assay to ethylene
The triple response assay was conducted to investigate the ethylene sensitivity of F1hybrid lines according to the method of Mubarok et al. (2015). Brief ly seeds were sterilized for 20 min and grown in a 50-mL glass bottle sealed with rubber septum and containing 10 mL of 1/2 Murashige and Skoog (MS) medium (Murashige and Skoog 1962). Total of 5 µL L-1of exogenous ethylene was injected into the sealed glass bottles containing seeds and kept in the dark at 25°C for 7 days.
2.4. Statistical analysis
A completely randomized design was used for this experiment. The data were represented as the mean value±SE of four replicates. For the statistical data analysis, data were subjected to Student's t-tests at P<0.05 for comparisons between the investigated data and the control.
3. Results
Under homozygous condition, the reduction of seed quality was detected in Sletr1-1 and Sletr1-2 mutant alleles. Reduction of germination rate of Sletr1-1 and Sletr1-2 was 20 and 16% lower than WT-MT, and also for emergence speed index was 48 and 43% lower than WT-MT for Sletr1-1 and Sletr1-2, respectively. For vigor index, mutation in Sletr1-2 showed the comparable effect compared with WT-MT, but mutation in Sletr1-1 signif icantly decrease the vigor index (Fig. 1). Improving the seed quality as an effect of Sletr1-2 mutation was detected in heterozygous lines of Sletr1-2 F1, but the effect of Sletr1-1 mutation in improving the seed quality did not detect in Sletr1-1 F1 (Fig. 2). Sletr1-1 F1seeds exhibited a reduction in the germination rate compared to WT-MT F1seeds in all the parental backgrounds, Intan, Mutiara and Ratna (Fig. 3-A). Based on this result, the Sletr1-1 mutation gave a similar response on the reduction of germination rate for several different genetic backgrounds. The data showed that the lowest germination rate was detected in the hybrid line of Sletr1-1×Intan, 29.5% lower than WT-MT F1hybrid seeds. On the other hand, it can be seen that the reduction of germination rate did not occur in Sletr1-2 F1seeds. Even when the Sletr1-2 was crossed with varieties having a different genetic background, the germination rate was not changed compared to WT-MT F1seeds.
The negative impact of the Sletr1-1 mutation was also observed in the reduction of seed vigor index and emergence speed index. However, the negative impact was not detected in the Sletr1-2 F1hybrid seeds from all parental backgrounds (Fig. 3). The reduction of vigor index was signif icantly detected in Sletr1-1 F1seed with a parental background of Intan and Mutiara, but it was not detected under the Ratna parental background. The vigor index of Sletr1-1 F1hybrid from Intan and Mutiara parental backgrounds was 3.5 and 1.5 times lower than the vigor index of WT-MT F1seeds, respectively. The reduction of emergence speed index of hybrid seeds as an effect of Sletr1-1 mutation was detected in the F1lines from all parental backgrounds. The emergence speed index of Sletr1-1 F1hybrid seeds was 7.36, 9.27, and 8.99 or 6.44, 5.14, and 4.23 lower than WT-MT F1hybrid seeds for the parental background of Intan, Mutiara and Ratna.

Fig. 1 Impact of Sletr1-1 and Sletr1-2 mutations on the seed quality (germination rate, vigor index and emergence speed index). WT-MT, wild-type Micro-Tom. The mean value±SE (four replicates) followed by the asterisk are signif icantly different from the control (WT-MT) based on Student's t-tests at P<0.05.
A reduction of ethylene sensitivity was observed in all of the Sletr1-1 F1and Sletr1-2 F1hybrid lines by increasing the length of shoot and roots on seedlings under 5 µL L-1of exogenous ethylene exposure. Sletr1-1 conferred insensitivity to the seedlings of those hybrid lines, whereas Sletr1-2 conferred reduced sensitivity to the seedlings of those hybrid lines. Under 5 µL L-1of exogenous ethylene, hypocotyl and shoot length of all of Sletr1-1 F1hybrid lines signif icantly longer than WT-MT F1hybrid lines. The shoot length of Sletr1-1 F1hybrid lines were 7.07, 6.51 and 6.5 cm longer than WT-MT F1hybrid line under a parental background of Intan, Ratna and Mutiara (Fig. 4-A). Whereas, the root length of Sletr1-2 F1hybrid line were 2.92, 3.27 and 3.23 cm longer than WT-MT F1hybrid line under parental background of Intan, Mutiara and Ratna (Fig. 4-B).
4. Discussion

Fig. 2 Representative pictures for seed growth of WT-MT F1, Sletr1-1 F1 and Sletr1-2 F1 hybrid lines crossed with parental background of Intan. WT-MT, wild-type Micro-Tom.

Fig. 3 Impact of Sletr1-1 and Sletr1-2 mutation on the seed quality (germination rate, vigor index and emergence speed index) of F1 hybrid lines in different parental backgrounds, Intan, Mutiara, and Ratna. WT-MT, wild-type Micro-Tom. The mean value±SE (four replicates) followed by the asterisk are signif icantly different from the control (WT-MT F1 line) based on Student's t-tests at P<0.05.
It has been proposed that the Sletr1 mutant alleles (Sletr1-1 and Sletr1-2) and Sletr1 mutant allele (Sletr1-4) are potential breeding materials for extending shelf life of tomato fruits (Okabe et al. 2011; Mubarok et al. 2019). Commercialization of new hybrid lines using Sletr1-1 or Sletr1-2 would contribute to reducing the postharvest loss of tomato fruits. Sletr1-1 and Sletr1-2 have a mutation in the f irst and second transmembrane domain of the ethylene receptor gene (SlETR1), respectively; therefore the effect of ethylene can be minimized and f inally the fruit shelf life was increased (Okabe et al. 2011; Mubarok et al. 2015). However, it is speculated that its mutation might affect the seed quality, which is another important breeding trait in tomato. Therefore this study evaluated the effects of Sletr1-1 and Sletr1-2 mutations on the hybrid seed quality.
High quality seeds are required for the commercialization of hybrid seeds. Several parameters are used to determine the seed quality such as seed germination, seed vigor and emergence speed index. Numerous studies demonstrate that ethylene production has a correlation with the seed germination, therefore suggesting the involvement of ethylene in the regulation of seed development, germination and dormancy (Matilla 2000). Corbineau et al. (2014) stated that the application of exogenous ethylene improves germination in numerous species and stimulates the germination under non-optimal environmental conditions. Inhibition of ethylene action and response by the mutation of ethylene receptor gene resulted in a different seed quality characteristics depend on the mutation location in SlETR1 gene. Undesirable effects of Sletr1-1 mutation compared to the control were observed in the Sletr1-1 F1hybrid seeds on different parental backgrounds. The Sletr1-1 mutation signif icantly affects the reduction of seed germination, seed vigor and emergence speed index of the F1hybrid lines in all the different parental backgrounds, Intan, Mutiara and Ratna. However, these effects were not detected in the Sletr1-2 F1hybrid lines (Fig. 3). Sletr1-1 mutation conferred complete ethylene insensitivity in the F1hybrid lines. It was shown that the application of 5 µL L-1ethylene did not affect the hypocotyl and primary root elongation. However, in Sletr1-2 F1hybrid lines, ethylene slightly affects the reduction of hypocotyl and primary root elongation (Fig. 4), which correspond with our previous study (Mubarok et al. 2015). These results suggest that Sletr1-1 and Sletr1-2 mutations have the same phenotype effect on different parental background, which exhibited a dominant inheritance pattern.

Fig. 4 Ethylene sensitivity of Sletr1-1 and Sletr1-2 F1 hybrid lines under 5 µL L-1 of exogenous ethylene exposure. WT-MT, wildtype Micro-Tom. The mean value±SE (four replicates) in with or without ethylene treatment followed by asterisks is signif icantly different from the control (WT-MT F1 line) based on Student's t-tests at P<0.05.
Reduction in seed quality due to Sletr1-1 mutation in this study is consistent with the Nr mutant seeds that exhibited a reduction in seed quality in Rutgers and Ailsa Craig parental backgrounds (Siriwitayawan et al. 2003). The same undesirable effects on seed quality for both mutations might be due to the same mutation location in ethylene receptor gene. Those mutants had a mutation located in the f irst transmembrane domains, which is an important region for ethylene binding. Amino acid substitution of P51L in the Sletr1-1 mutant as a result of mutation in the f irst transmembrane domain is similar to the amino acid substitution of P36L in Nr (Mubarok et al. 2015). Another study showed a poor germination rate is also observed in the Arabidopsis mutant etr1-1, ethylene-insensitive mutants, which did not respond to exogenous ethylene (Bleecker and Schaller 1996; Bleecker et al. 1988). Based on these studies, we conf irm that seeds that are strongly impaired for ethylene perception exhibited slower germination compared to wild-type seeds indicating reduced seed quality, whereas those only weakly impaired for ethylene perception exhibited almost similar germination to the wild-type seeds.
Abscisic acid (ABA) plays a role in the induction of dormancy and in the maintenance of dormancy during seed imbibition (Cutler et al. 2010; Nambara et al. 2010), whereas ethylene regulates seed germination and dormancy by decreasing sensitivity to endogenous ABA (Matilla 2000; Arc et al. 2013). Ethylene insensitivity led to a greater sensitivity to exogenous ABA and greater ABA biosynthesis (Beaudoin et al. 2000; Ghassemian et al. 2000). Inhibition of ethylene perception in Sletr1-1 F1hybrid lines might affect the increasing ABA biosynthesis; therefore it affects the reduction of seed germination and vigor. Interestingly in Sletr1-2 F1hybrid seeds, there was no clear reduction in seed quality. The germination rate of Sletr1-2 F1hybrid lines seed relative to WT-MT F1hybrid seed might be due to the Sletr1-2 mutation being a moderate ethylene insensitive mutation that displays some limited ethylene perception (Okabe et al. 2011). Therefore, ethylene may still affect the seed germination and development. Several studies on tobacco seed showed that ethylene also acts in up-regulation of the gene for 1,3-glucanase, an enzyme participating in endosperm rupture (Leubner-Metzger et al. 1998; Leubner-Metzger and Meins 2000).
5. Conclusion
Sletr1-1 and Sletr1-2 mutations resulted in the different effect on the seed quality of F1hybrid lines. Any undesirable effects of Sletr1-2 mutation on the seed quality, i.e., germination rate, vigor index and emergence speed index have not been detected in Sletr1-2 F1hybrid seeds. We conclude that the Sletr1-2 mutant has a potential as a contributor to breeding programs for the new hybrid tomato cultivars with improved fruit shelf life.
Acknowledgements
This work was supported by a grant of Superior Applied Research of Higher Education (006/ADD/SP2H/DRPM/VIII/2017) from the Ministry of Research, Technology and Higher Education, Indonesia. We thank the National BioResource Project (NBRP), MEXT, Japan for providing the seeds of S. lycopersicum cv. Micro-Tom, Sletr1-1 and Sletr1-2. We thank the Vegetable Crops Research Institute, Ministry of Agriculture, Indonesia for providing the seeds of S. lycopersicum cvs. Intan, Mutiara and Ratna. We also thank all of the members of our laboratory for helpful discussions throughout the work.
 Journal of Integrative Agriculture2019年5期
Journal of Integrative Agriculture2019年5期
- Journal of Integrative Agriculture的其它文章
- Mulching with plastic f ilm improved the root quality of summersown sweet potato (Ipomoea batatas (L). Lam.) in northern China
- The chemical treatments combined with antagonistic yeast control anthracnose and maintain the quality of postharvest mango fruit
- Comparison of phenolic prof iles and antioxidant activities in skins and pulps of eleven grape cultivars (Vitis vinifera L.)
- Effect of grazing time and intensity on growth and yield of spring wheat (Triticum aestivum L.)
- Polyaspartic acid mediates the absorption and translocation of mineral elements in tomato seedlings under combined copper and cadmium stress
- lnteraction effect of nitrogen form and planting density on plant growth and nutrient uptake in maize seedlings