
lnteraction effect of nitrogen form and planting density on plant growth and nutrient uptake in maize seedlings
2019-05-10 06:13WANGPengWANGZhangkuiSUNXichaoMUXiaohuanCHENHuanCHENFanjunYuanLixingMlGuohua
WANG Peng, WANG Zhang-kui, SUN Xi-chao, MU Xiao-huan, CHEN Huan, CHEN Fan-jun, Yuan Lixing, Ml Guo-hua
Department of Plant Nutrition, College of Resources and Environmental Sciences, China Agricultural University, Beijing 100193, P.R.China
Abstract High planting density is essential to increasing maize grain yield. However, single plants suffer from insuff icient light under high planting density. Ammonium () assimilation consumes less energy converted from radiation than nitrate (). It is hypothesized that a mixed / supply is more important to improving plant growth and population productivity under high vs. low planting density. Maize plants were grown under hydroponic conditions at two planting densities (low density: 208 plants m-2 and high density: 667 plants m-2) and three nitrogen forms (nitrate only, 75/25/ and ammonium only). A signif icant interaction effect was found between planting density and N form on plant biomass. Compared to nitrate only, 75/25/ increased per-plant biomass by 44% under low density, but by 81% under high density. Treatment with 75/25/increased plant ATP, photosynthetic rate, and carbon amount per plant by 31, 7, and 44% under low density, respectively, but by 51, 23, and 95% under high density. Accordingly, carbon level per plant under 75/25/was improved, which increased leaf area, specif ic leaf weight and total root length, especially for high planting density, increased by 57, 17 and 63%, respectively. Furthermore, under low density, 75/25/increased nitrogen uptake rate, while under high density, 75/25/ increased nitrogen, phosphorus, copper and iron uptake rates. By increasing energy use eff iciency, an optimum / ratio can improve plant growth and nutrient uptake eff iciency, especially under high planting density. In summary, an appropriate supply of in addition to nitrate can greatly improve plant growth and promote population productivity of maize under high planting density, and therefore a mixed N form is recommended for high-yielding maize management in the f ield.
Keywords: maize, planting density, /ratio, carbon, nutrient uptake, root morphology
1. lntroduction
Maize (Zea mays L.) mainly grows on aerobic soils dominated by nitrate. High planting density is believed to be a major factor for increasing maize yield (Ciampitti and Vyn 2012; Hernández et al. 2014). However, because light availability per plant decreases with increasing planting density, the yield per plant decreases with increasing planting density. Thus, the eff icient use of energy within a plant becomes essential for plant growth under high planting density. As mentioned above, compared to nitrate, ammonium assimilation requires much less energy (Salsac et al. 1987). Therefore, it is hypothesized that partial ammonium in addition to nitrate supply should increase energy use eff iciency and improve plant growth and population productivity, especially under high planting density. At present, the interaction effect between N form and planting density on plant growth has not been determined, especially from the viewpoint of energy consumption. The aim of this study was to test the above hypothesis, identify the underlying physiological mechanism and provide insight for the recommendation of N management in high-yielding maize production.
2. Materials and methods
2.1. Experimental procedures
Hydroponic experiments were conducted in a growth chamber with light intensity of 350 µmol m-2s-1, day/night temperature of 28/22°C and 60% relative humidity. Seeds of hybrid ZD958 were sterilized by 10% (v/v) H2O2for 30 min, rinsed with deionized water and soaked in saturated CaSO4for 6 h, then transferred to germinate on f ilter paper in dark conditions. When the primary root was 1.5 cm long, the seeds were transferred to culture in rolled papers. When the seedlings had one expanded leaf, their endosperm was removed and were transferred to a container (55 cm ×45 cm×35 cm) with low (208 plants m-2) and high (667 plants m-2) planting density (Appendix A). Plants were supplied with modif ied Hoagland nutrient solution containing 0.5 mmol L-1K2SO4, 0.6 mmol L-1MgSO4·7H2O, 0.3 mmol L-1KH2PO4, 0.5 mmol L-1CaCl2·2H2O, 1 µmol L-1H3BO3, 0.5 µmol L-1MnSO4·H2O, 0.5 µmol L-1ZnSO4·7H2O, 0.2 µmol L-1CuSO4·5H2O, 0.07 µmol L-1Na2MoO4·2H2O, and 0.1 mmol L-1Na-Fe-EDTA. In our preliminary experiment, plant growth was the highest at the NH4+to NO3-ratio of 75 to 25% (Appendix B) and the optimum N concentration was 1 mmol L-1(Appendix C). Therefore, N was supplied in 1 mmol L-1with three different/ratios (nitrate only, 75/25/and ammonium only) using KNO3and/or (NH4)2SO4. MgSO4and K2SO4were added to balance potassium difference in the solution (Gu et al. 2013). Solution p H was adjusted to 5.8 every 6-12 h. The containers were randomly placed and their places were changed frequently. The nutrient solution was aerated continuously and renewed every 3 days.
2.2. Gas exchange and chlorophyll f luorescence parameters
Twelve days after N treatments, 4 h after lighting, the most recent fully expanded leaf was used to measure net photosynthetic rate (Pn), conductance to H2O (Cond), intercellular CO2concentration (Ci) and transpiration rate (Tr) for 8 plants from each treatment. All parameters were measured by a portable photosynthesis system (Li6400; LI-COR, Lincoln, NE, USA) coupled to standard red/blue LED broadleaf cuvette (6400-02B; LI-COR) and a CO2mixer (6400-01; LI-COR) at a light intensity of 400 µmol m-2s-1. Measurements were obtained at a leaf temperature of (28±0.5)°C and a CO2concentration inside the chamber of (400±1) µmol L-1.
2.3. Shoot growth and root morphological parameters
Fifteen seedlings from each treatment were sampled on 0, 3, 6, 9, 12 and 15 days after transplanting, respectively. Leaf area was measured by a ruler. The total root length was determined by WinRHIZOTM2003b Software (Regent Instruments, QC, Canada) after root image was obtained with a scanner.
2.4. ATP
Samples of ground fresh shoots and roots (100 mg) were put in the centrifuge tube, 900 µL of 0.01 mol L-1PBS buffer (p H=7.3) was added, and the mixture was centrifuged at 3 000 r min-1for 20 min. The supernatant was used to determine ATP concentration using an ELISA Kit (Crowther et al. 1995). There were six biological replicates for each treatment.
2.5. Carbon content
Milled dry shoot and root samples (80 mg) were used to measure C concentration using an NC analyzer (vario MACRO cube, ELEMENTAR, Germany). There were f ive biological replicates for each treatment.
2.6. Mineral elements
About 200 mg shoot and 100 mg root sample were used to measure N, P and K (Murphy and Riley 1962; Nelson and Sommers 1973). The concentration of Zn, Mn, Fe, Cu and Mg and other micro mineral elements was determined using a plasma emission spectrometer ICP-AES (OPTIMA 3300 DV, Perkin-Elmer, USA). The standard sample IPE200 (WEPAL, The Netherlands) was used to correct the error.
2.7. Parameter calculation
Based on above measurements, we calculated the following parameters.
Leaf area=Length×Width×k
Where, k value is 0.75 if a leaf is fully expanded, and is 0.5 if a leaf is not fully expanded (Gallais et al. 2006).
Specif ic leaf weight (SLW)=Leaf biomass per plant/Leaf area per plant (Pearce et al. 1969)
Specif ic root length (SRL)=Total root length per plant/Root biomass per plant (Smit et al. 2013)
Nutrient uptake rate=(W2-W1)/(t2-t1)×ln[(R2/R1)/(R2-R1)]
Where, W1and W2are the nutrient contents of the plants at times (t) 1 and 2, and R1and R2are total root length measured at t1and t2, respectively (Williams 1948).
2.8. Statistical analysis
Mean and standard deviation were calculated using Microsoft EXCEL 2010 and SigmaPlot 10.0. Data were analyzed by two-way analysis of variance (ANOVA) with SPSS 17.0 and SAS 9.0.
3. Results
3.1. Biomass
Compared to nitrate only or ammonium only, the mixed form of nitrate and ammonium increased shoot, root and total biomass of maize on either a per plant or population basis (Fig. 1). Under low planting density, per plant biomass was similar between nitrate only (1.54 g) and ammonium only (1.57 g). Compared to nitrate only, the mixed form of nitrate and ammonium increased per plant biomass by 40% at low planting density. Under high planting density, plants grown in ammonium only (0.71 g) had higher biomass per plant than with nitrate only (0.56 g). Compared to nitrate only, mixed form of nitrate and ammonium increased biomass per plant by 81% at high planting density.
Population biomass was signif icantly greater under high planting density compared to low planting density (Fig. 1-F). Under low planting density, the mixed N form increased population biomass by 40 and 38% compared to nitrate only and ammonium only, respectively. Under high planting density, the mixed N form increased population biomass by 81 and 43% compared to nitrate only and ammonium only, respectively.
3.2. ATP
ATP concentration and content of the shoot and root were affected by both planting density and N form. Root ATP concentration and content were also affected by the interaction of N form and planting density (Table 1). High planting density increased ATP concentration in the shoot but not in the root. However, ATP content per plant was reduced by high density in both shoot and root. Compared to nitrate only, shoot ATP content in plants treated with the mixed N form was increased by 36 and 37% under low and high planting density, respectively. Root ATP content was increased by 44 and 69% under low and high planting density, respectively (Table 1).
3.3. Photosynthetic rate, transpiration rate, intercellular CO2 concentration and stomatal conductance
Compared to nitrate only, the mixed N form did not change photosynthetic rate, transpiration rate, or intercellular CO2concentration under low planting density. Under high planting density, however, the mixed N form increased photosynthetic rate and transpiration rate by 23 and 28%, respectively. Intercellular CO2concentration was decreased by 21%. The stomatal conductance to H2O was affected slightly by N forms under both low and high density (Table 2). In general, plants treated with ammonium only had the highest photosynthetic rate and transpiration rate.
3.4. Carbon concentration and carbon content
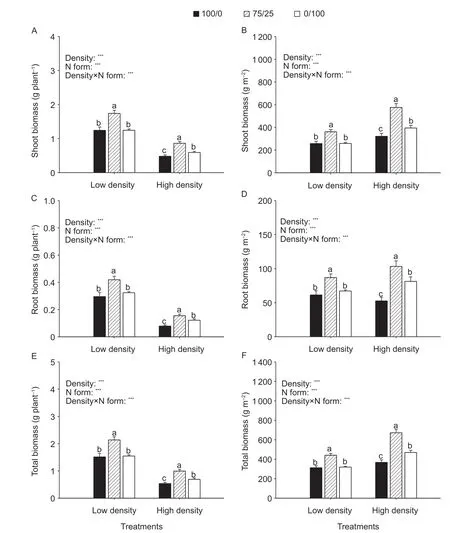
Fig. 1 Shoot biomass, root biomass and plant biomass of per plant (A, C and E) and unit area (B, D and F) as affected by N forms and planting density. 100/0, 75/25 and 0/100 represent nitrate only, 75/25/ and ammonium only, respectively. Values are mean and SE (n=10). At the same pant density, significant differences at P<0.05 are shown with different letters. *, **, and ***indicate significant difference at 0.05, 0.01 and 0.001 probability levels, respectively.
In the shoot, carbon concentration was not affected by N form under low planting density, but was 8% higher in the mixed N form compared to nitrate only (Fig. 2-A and B). In the root, carbon concentration was 6% higher in the mixed N form compared to nitrate only under both low and high planting density (Fig. 2-C and D). There was no difference in carbon concentration between the mixed N form and ammonium only in both shoot and root under either low or high planting density. Compared to nitrate only, the mixed N form increased carbon content by 42 and 92% in the shoot, and by 50 and 108% in the root, under low and high planting density, respectively. In total, the mixed N form increased carbon amount per plant by 44 and 95% under low and high planting density, respectively.
3.5. Leaf area and specif ic leaf weight
The effect of N form and planting density on leaf area (LA) per plant was identical to that on shoot biomass (Fig. 3-A). Compared to nitrate only, leaf area in mixed N form increased leaf area by 21% under low planting density and by 57% under high planting density. Specif ic leaf weight (SLW) was similar among different N forms under low planting density (Fig. 3-B). Under high planting density, SLW in the mixed N treatment was 1.17 times of that in nitrate only treatment. The number of leaves per plant under different N forms was not different under low or high planting density (Appendix D).
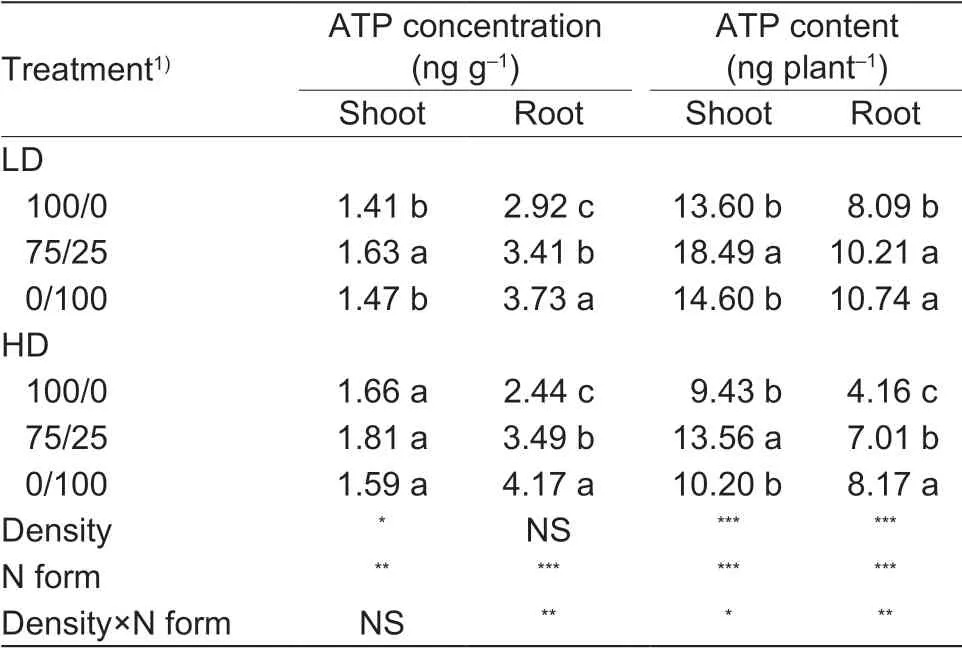
Table 1 Shoot and root ATP concentration and ATP content as affected by N form and low and high plant density
3.6. Root length and specif ic root length
There was no difference in total root length per plant (TRL) between nitrate only and ammonium only treatment (Fig. 4-A and B). TRL at mixed N form was 1.25 and 1.63 times higher than that in nitrate only under low and high planting density, respectively. Specif ic root length (SRL) was signif icantly higher under nitrate only than that under the mixed N form or ammonium only treatment in both plant densities (Fig. 4-C and D).
3.7. Nutrient uptake rate
The uptake rate of all nutrients was reduced by high planting density (Table 3). Compared to nitrate only, the mixed N form increased nutrient uptake rate of N by 12%, and reduced that of Fe and Mn by 12 and 43%, respectively, without much effect on the other nutrients (P, K, Ca, Cu, Mg and Zn) under low planting density. Under high planting density, the mixed N form increased N, P, Cu and Fe uptake rate by 18, 38, 43, and 42%, respectively, without much effect on the other nutrients. Compared to nitrate only, shoot P, K, Ca, Mg, Mn and Zn concentrations decreased under low planting density in the mixed N form, without much effect on N, Cu, and Fe concentrations, while Ca, Mg and Mn concentrations of shoots decreased under high density, without much effect on N, P, K, Cu, Fe and Zn concentrations (Appendix E).
4. Discussion
Insuff icient light and carbohydrate assimilation are the main factors limiting maize plant growth under high planting density (Rossini et al. 2011; Ciampitti and Vyn 2012; Timlin et al. 2014). During photosynthesis, light energy is converted into chemical energy which is further used for carbon assimilation and various biological functions (Marschner 2011). Nitrogen assimilation consumes about 25% of the total energy in the plant (Bloom et al. 1989, 1992). Therefore, if an appropriate amount of ammonium is supplied instead of nitrate, the chemical energy can be saved for assimilation of more carbon, to increase plant growth. The results of this study support this relationship. Compared to nitrate only, the mixed N form signif icantly increased plant biomass: total plant biomass was increased by 40% under low density, but by 81% under high density, suggesting that a mixed form of ammonium and nitrate has an even greater effect on plant growth under high planting density in which light level per plant is poor and within-plant energy form can be more limiting for photosynthesis and plant growth.
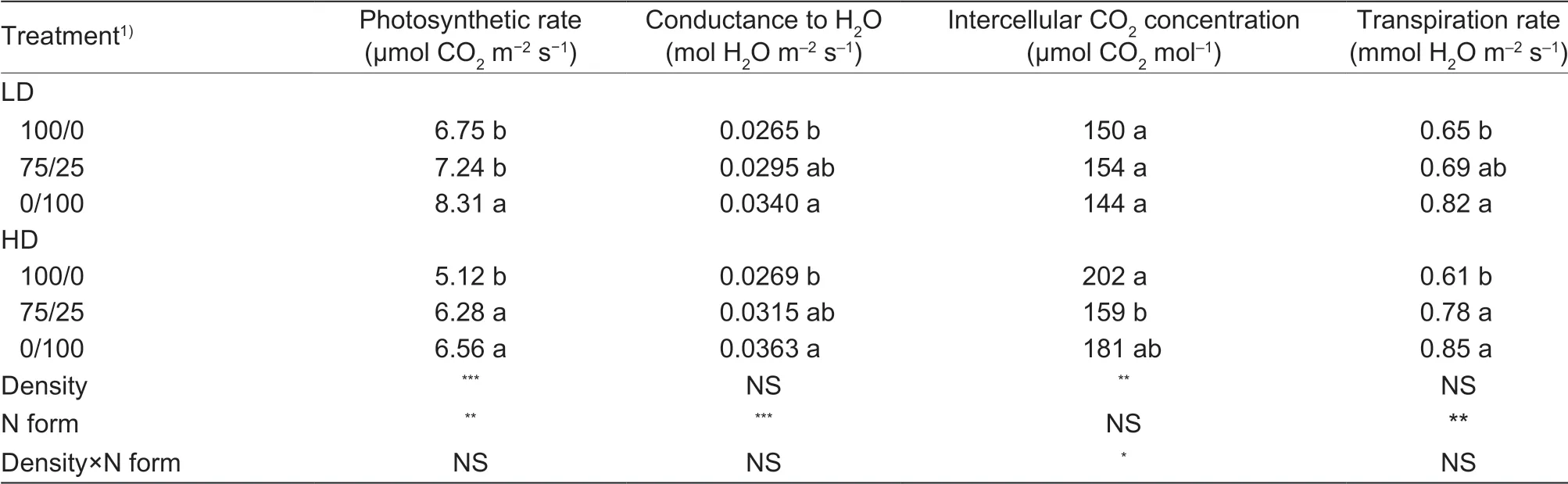
Table 2 The net photosynthetic rate, conductance to H2O, intercellular CO2 concentration and transpiration rate as affected by N forms and low and high planting density
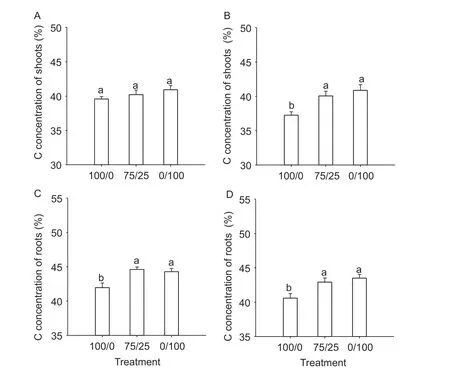
Fig. 2 Carbon (C) concentration of shoots and roots of plants grown in different N forms under low (A and C) and high planting density (B and D). 100/0, 75/25 and 0/100 represent nitrate only, 75/25/ and ammonium only, respectively. Values are mean and SE (n=5). Under the same panting density, significant differences at P<0.05 are shown with different letters.
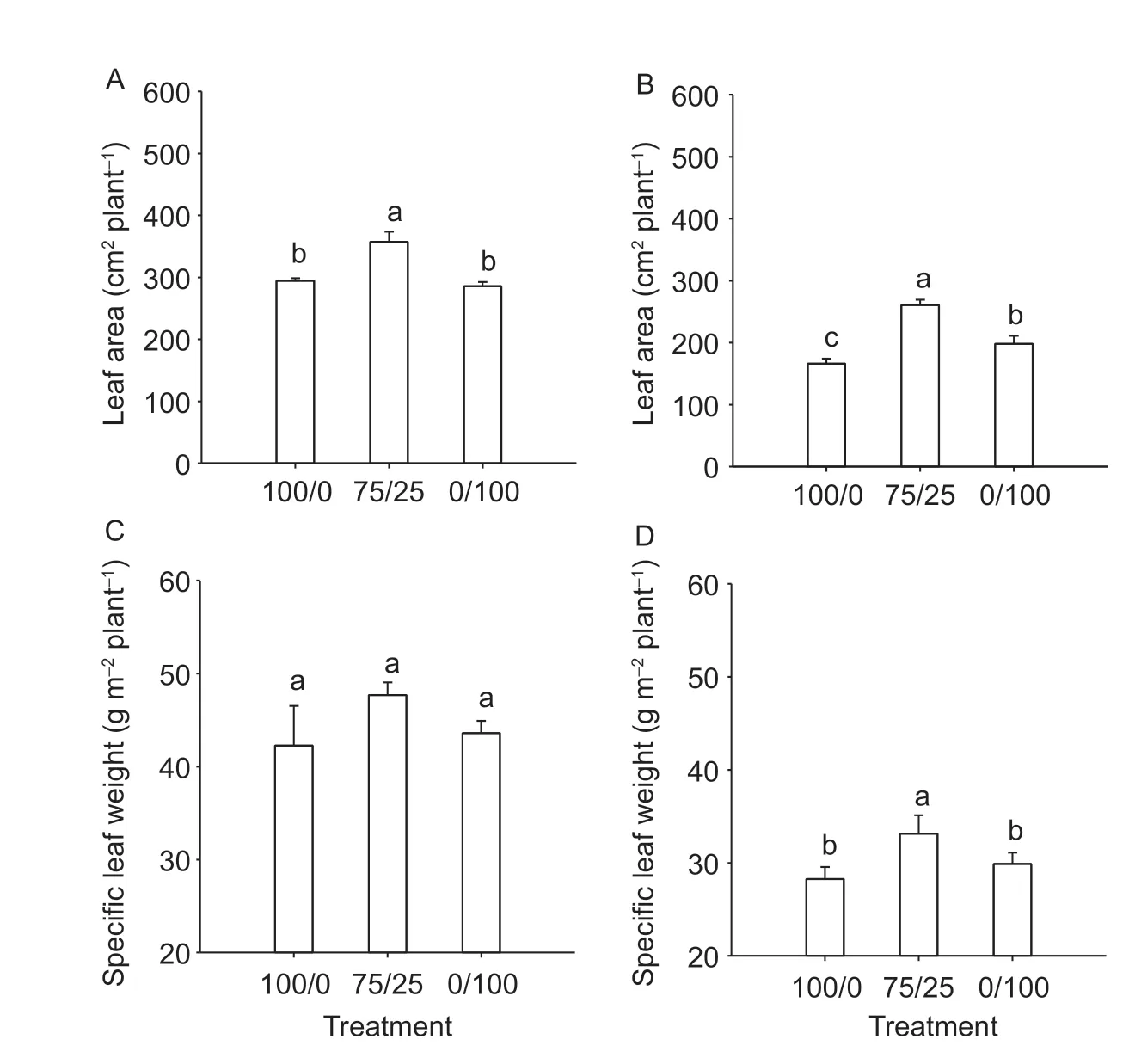
Fig. 3 Leaf area and specific leaf weight per plant grown in different N forms under low (A and C) and high (B and D) planting density. 100/0, 75/25 and 0/100 represent nitrate only, 75/25/ and ammonium only, respectively. Values are mean and SE (n=5). Under the same panting density, significant differences at P<0.05 are shown with different letters.

Fig. 4 Total root length and specific root length per plant grown in different N forms under low (A and C) and high (B and D)planting density. 100/0, 75/25 and 0/100 represent nitrate only, 75/25/ and ammonium only, respectively. Values are mean±SE (n=5). At the same day, significant differences at P<0.05 are shown with different letters. NS, no significant difference.
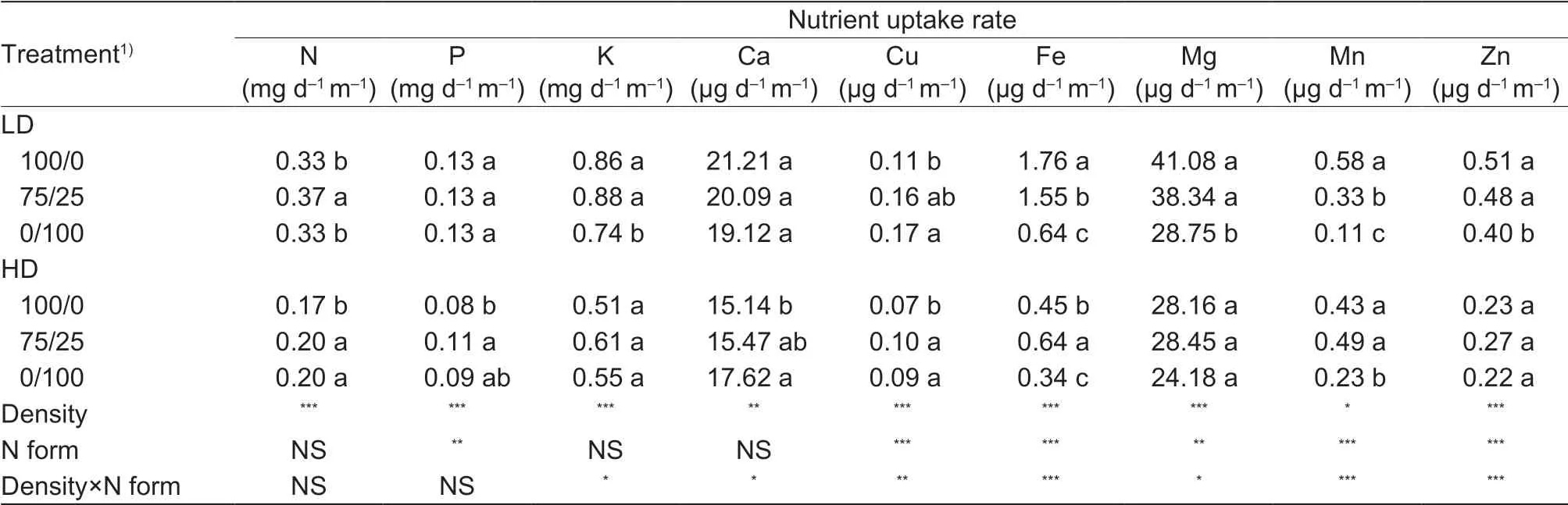
Table 3 Uptake rate of N, P, K, Ca, Cu, Fe, Mg, Mn and Zn per unit root length as affected by N forms under low and high plant density
In this study, compared to nitrate only, ATP content per plant under the mixed N form increased 31% at low planting density and 51% at high planting density. The high ATP content in the plant contributed to the difference in ATP consumption between assimilation of nitrate only and the mixed N form, because the assimilation of nitrate requires more ATP (Fuhrer and Erismann 1984; Salsac et al. 1987; Bloom et al. 1992). The saved ATP allows more photosynthesis and carbon accumulation (Werf et al. 1988; Sage 1994; Guo et al. 2007), and therefore led to further ATP accumulation. Compared to low density, plants grown under high planting density receive poor light form and suffer from insuff icient ATP supply. Therefore, the ATP saved in the mixed N form should further increase photosynthesis and carbon assimilation, which in turn, increase ATP level in the plant. In this study, the mixed nitrogen form increased photosynthetic rate by 23% under high planting density, but only 7% under low planting density (Table 1). Accordingly, the intercellular CO2concentration of the leaf was reduced by 21% by the mixed nitrogen form under high planting density, but not low planting density (Table 2). As a result, compared to nitrate only, total carbon content per plant under mixed nitrogen form increased by 95% under high planting density, but only by 44% under low planting density (Fig. 2). The stomatal conductance was slightly affected by the mixed N form, suggesting that the increased photosynthesis is not contributed by CO2transport (Table 2). The improvement of carbon level in the plant explained the larger leaf area under the mixed N form compared to nitrate only under high but not low planting density. The improvement of carbon levels under high planting density in the mixed N form is further supported by the difference in specif ic leaf weight. For a species or individual, low specif ic leaf weight is typically a response to light limitations that result in decreased carbon gain (Baird et al. 2017), just as shown in the high planting density treatment in this study. The mixed N form increased SLW, with greater effect under high vs. low planting density (Fig. 3).
Compared to the mixed N form, ammonium only had higher photosynthetic rate but lower shoot ATP content and lower carbon content per plant. The reason is that, compared to the mixed N form, ammonium only had lower leaf area. In tobacco, in comparison to nitrate only, ammonium is unfavorable for leaf expansion because of low cytokinin levels (Walch et al. 2000). In the current study, leaf area was the same between nitrate only and ammonium only treatments. The effect of ammonium on leaf growth may be species-dependent or related to the ammonium concentration supplied.
High planting density reduced total root length per plant, a result of insuff icient supply of carbon from shoot to root. However, SRL was also increased by high planting density (Fig. 4), suggesting that an increase of SRL is an adaptive response to insuff icient carbohydrate (Van Hees 1997; Ostonen et al. 2007; Xie et al. 2007). Compared to nitrate, the mixed N form reduced SRL while increasing total root length under low and high planting density, indicating that carbon status at mixed N form was relatively improved, especially for high planting density.
Early calculations suggested that up to 36% of total respiratory energy cost, expressed as ATP consumption, is required for ion uptake (Werf et al. 1988; Marschner 2011). Kurimoto et al. (2004) reported that 76% of total energy cost was required to low-aff inity nitrate inf lux across the plasma membrane of cereal roots (Kurimoto et al. 2004). In this study, the uptake rates of all nutrients were reduced by high planting density, possibly due to the insuff icient carbohydrate and therefore insuff icient ATP production in the roots. Interestingly, nutrient uptake rates under mixed N form was improved for N, P, Cu and Fe uptake rates, possibly because of improved carbon and energy status in the roots. Even under low planting density, the mixed N form increased N uptake rate compared to nitrate only.
5. Conclusion
The main results in the present study are summarized in Fig. 5. Compared to nitrate only, a mixed form of ammonium and nitrate at an optimum ratio can improve maize plant growth. This effect was greater under high planting density in which light availability was insuff icient. The mixed form of ammonium and nitrate increased ATP, which contributed to higher net photosynthetic rate, and more carbon accumulation. Higher carbon level led to larger leaf area, which in turn helped to increase carbon accumulation and plant ATP level. As a result, shoot and root biomass was increased. Furthermore, higher carbon level increased the total root length and nutrient uptake rate of N, P, Cu and Fe. Compared to mixed N form, the supply of ammonium only did not increase leaf area, which results in a reduced effect on shoot biomass.

Fig. 5 The model explaining the effect of N forms on maize growth under low and high planting density. Mixed N form compared to nitrate only (A and B) and ammonium only compared to nitrate only (C and D) under low and high planting density for ZD958 under 1 mmol L-1 nitrogen. Solid arrows represent the promotion effect; solid lines and pentagrams represent the suppression effect; dotted arrows represent feedback facilitation.
Acknowledgements
This work is supported by the National Basic Research Program of China (2015CB150402) and the National Natural Science Foundation of China (31672221 and 31421092).
Appendicesassociated with this paper can be available on http://www.ChinaAgriSci.com/V2/En/appendix.htm
 Journal of Integrative Agriculture2019年5期
Journal of Integrative Agriculture2019年5期
- Journal of Integrative Agriculture的其它文章
- Characterization of TaCOMT genes associated with stem lignin content in common wheat and development of a gene-specif ic marker
- Phenotypic characterization and genetic mapping of the dwarf mutant m34 in maize
- Morphological diversity and correlation analysis of phenotypes and quality traits of proso millet (Panicum miliaceum L.) core collections
- Field identif ication of morphological and physiological traits in two special mutants with strong tolerance and high sensitivity to drought stress in upland rice (Oryza sativa L.)
- Crosstalk of cold and gibberellin effects on bolting and f lowering in f lowering Chinese cabbage
- Foliar spraying of aqueous garlic bulb extract stimulates growth and antioxidant enzyme activity in eggplant (Solanum melongena L.)