
Crosstalk of cold and gibberellin effects on bolting and f lowering in f lowering Chinese cabbage
2019-05-10 06:13SONGShiweiLElYulingHUANGXinminSUWeiCHENRiyuanHAOYanwei
SONG Shi-wei, LEl Yu-ling, HUANG Xin-min, SU Wei, CHEN Ri-yuan, HAO Yan-wei
College of Horticulture, South China Agricultural University, Guangzhou 510642, P.R.China
Abstract The f lower stalk is the product organ of f lowering Chinese cabbage (Brassica campestris L. ssp. chinensis var. utilis Tsen et Lee), which is cultivated extensively in South China. Flower stalk formation and development, including bolting and f lowering, determine the yield of f lowering Chinese cabbage; however, the bolting and f lowering mechanisms remain to be explored. To elucidate these processes, we studied the effects of low-temperature and gibberellin (GA) treatments, and their interaction, on stem elongation, bolting time, f lowering time, hormone content, and cell morphology in stem of f lowering Chinese cabbage. The results showed that both cold and GA treatments accelerated bolting time, stem elongation, and f lowering time. Moreover, cold and GA cotreated plants displayed additive positive effects. In addition, cold treatments increased the GA, indole-3-acetic acid, and cytokinin contents and altered cell size in the shoot apices of f lowering Chinese cabbage. Treatment with uniconazole, a GA synthesis inhibitor, strongly delayed bolting time, stem elongation, and f lowering time, whereas GA, but not cold treatment, rescued this inhibition, indicating that low temperature accelerates bolting and f lowering not only through inducing GA in the shoot apices, but also other ways. These results provide a theoretical basis for further dissecting the regulatory mechanism of bolting and f lowering in f lowering Chinese cabbage.
Keywords: bolting, f lowering, stem elongation, cold, gibberellin
1. lntroduction
The bolting and f lowering characteristics of cruciferous vegetables have been studied for many years (Amasino and Michaels 2010). Generally, the process of plant f lowering includes f lower bud initiation and differentiation, f lowering organ formation, bolting, and f lowering stages (Jung and Müller 2009; Amasino and Michaels 2010). Following seed germination, cruciferous vegetables undergo a phase of vegetative growth and subsequently enter a low-temperature induction period. After a suff icient period of low-temperature induction, these vegetables bolt and bloom under hightemperature and long-day conditions. The developing period of each of these phases differs among varieties (Sun et al. 2008).
The factors that affect the timing of f lowering and bolting in cruciferous vegetables include temperature, plant hormones, genetic factors, physiology, and biochemical substances (Pressman and Shaked 1988; Lambrechts et al. 1994; Yang et al. 1996; Yan and Hunt 1999). Low temperature is an important environmental signal for stem elongation and f lowering in many winter annuals and biennials. Long-term low-temperature treatment induces changes in the physiological and biochemical states of plants that perceive low temperature (Bunce 1985; Zanewich and Rood 1995; Dahanayake and Galwey 1999; Hébrard et al. 2013; Mathieu et al. 2014; Liang et al. 2018). The meristematic region of the stem is particularly sensitive to low temperature (Dahanayake and Galwey 1999; Mathieu et al. 2014). It has also been revealed that a vernalizationresponsive change in gibberellin (GA) metabolism is restricted to the shoot tip. GA has a strong effect on plant bolting and f lowering (Zanewich and Rood 1995; Yang et al. 1996; Olszewski et al. 2002; Ranwala and Miller 2008; Regnault et al. 2014). For example, exogenous GA3treatment promotes stem elongation and bolting in spring rape (Brassica napus), whereas blocking GA biosynthesis with chlorocholine chloride inhibits stem elongation (Dahanayake and Galwey 1999). GA reportedly is involved in the thermo-induction of f lowering and vernalization responses. During and after cold perception, marked changes in the metabolism and turnover of GA precursors have been reported in Thlaspi arense L. and Raphanus sativus L. (Dahanayake and Galwey 1999). In winter canola (B. napus cv. Crystal), differential GA metabolism between vernalized and non-vernalized plants has been observed, which corroborates the inf luence of vernalization on GA physiology (Zanewich and Rood 1995). In addition to GA, other plant hormones regulate stalk development through a complex regulatory network (Yang et al. 1996; Ross et al. 2011). Reductions in indole-3-acetic acid (IAA), cytokinin (CTK), and brassinolide (BR) contents or partial blockage of their signal transduction results in the inhibition of stem elongation (Xu et al. 1998; Tarkowská et al. 2012; Huang et al. 2017), whereas exogenous application of abscisic acid (ABA) stimulates tuber thickening in potato (Xu et al. 1998).
Flowering Chinese cabbage (Brassica campestris L. ssp. chinensis var. utilis Tsen et Lee) is a variant of a subspecies of Chinese cabbage that originates from South China. The major edible product of f lowering Chinese cabbage is the f lower stalk, the key developmental characteristics of which include stem elongation, thickening, and f lowering (Zhong et al. 2016). The bolting and f lowering of f lowering Chinese cabbage differ from those of other cruciferous vegetables. It is a typical spring plant, in which bolting and f lowering do not dependent on low temperature. Nevertheless, although f lowering Chinese cabbage normally bolts and f lowers without cold treatment, it also displays a response to cold temperature similar to that of other cruciferous vegetables. When the ambient temperature changes during planting, abnormal bolting and f lowering occur, which will reduce the yield and quality of f lower stalks. Therefore, elucidating the mechanisms underlying bolting and f lowering in f lowering Chinese cabbage is important for producers.
Low temperature and GA are the two main factors inf luencing plant bolting and f lowering. In this study, we investigated the effects of GA and a GA synthesis inhibitor (uniconazole) on f lower stalk elongation and f lowering in f lowering Chinese cabbage under both room and low temperatures. The results obtained will help us to understand the effects of low temperature and GA on bolting and f lowering in f lowering Chinese cabbage, and will provide new basic knowledge on the bolting and f lowering mechanisms in crucifers.
2. Materials and methods
2.1. Plant materials and treatments
Seeds of f lowering Chinese cabbage (late bolting-type cultivar Cuilv 80, Guangdong Academy of Agricultural Sciences, Guangzhou, China) were sterilized with 70% ethanol, washed f ive times with sterile water, and germinated in an incubator at 25°C. When the seeds started to germinate, they were divided into two groups. One group was maintained in the 25°C incubator (control), and the other group was exposed to low temperature (2, 7, or 15°C), with each treatment lasting for 1, 3, or 5 days. When the cold treatments were completed, all the seeds were planted in a greenhouse of the South China Agricultural University under 70-85% relative humidity and 28°C/20°C (14 h/10 h) day/night temperature to evaluate bolting time, bolting rate, f lower bud emergence time, f lowering time, and f lowering rate. Each treatment consisted of 30 plants. Thereafter, the plants were subjected to normal temperature and cold treatment (15°C, 5 days) for further studies. When the plants reached the seedling stage, stem length, stem diameter, leaf number, and fresh weight were measured. Each treatment consisted of 30 plants and six plants were selected randomly for measurements at each stage. In addition, the shoot apices (5 mm) were collected for hormone extraction and histological analyses. The same sampling and measurement procedures were performed when plants reached the f lower bud emergence, fast bolting, and f lowering stages.
Plants exposed to normal temperature and cold treatment (15°C, 5 days) were also selected for GA and uniconazole treatments. When plants were at the third true leaf stage, they were sprayed with water (control), GA (200 mg L-1; Shanghai Bo'ao Biological Technology Co., Ltd., Shanghai, China), uniconazole (40 mg L-1, Shanghai Bo'ao Biological Technology Co., Ltd., Shanghai, China), or GA (200 mg L-1) plus uniconazole (40 mg L-1). Plant bolting rate, f lower bud emergence rate, and f lowering rate were recorded daily. When the plants reached the f lowering stage, stem length, stem diameter, leaf number, and fresh weight were measured. Each treatment consisted of 30 plants and six plants were randomly selected at each stage for measurements.
2.2. Histological analysis of the shoot apices of f lowering Chinese cabbage
For histological analysis, tissue samples were collected from the same position on the shoot apices (5 mm) and were immersed in FAA (formalin-acetic acid-alcohol) solution, placed under vacuum for 15 min, and then incubated at 25°C for 72 h. The samples were dehydrated in a gradient series of ethanol and embedded in paraff in (Yan et al. 2015). Sections (8-µm thick) were stained with leagene and observed under a microscope. Approximately 80 to 100 cells were randomly selected to calculate the cell size using Image-Proplus 6.0. The mean cell size is reported. Each treatment was conducted in three replicates.
2.3. Hormone determination of the shoot apices of f lowering Chinese cabbage
Approximately 50 mg of shoot apex tissue was harvested from each plant. To the dissected tissue, 500 µL of 1× phosphate-buffered saline was added. The tissue was ground with a mortar and pestle, and then homogenized by sonication. The homogenate was centrifuged to pellet cellular debris and the supernatant was transferred to a clean tube for ELISA. The GA content of the tissue samples was analyzed using a plant gibberellic acid (GA) ELISA Kit (Shanghai Enzyme-Linked Biotechnology Co., Ltd.) following the manufacturer's protocol. The same protocol was used for IAA and CTK measurements.
2.4. Statistical analysis
Data from each treatment were analyzed by one-way analysis of variance using SigmaPlot (11.1) (Jandel Scientif ic Software, San Rafael, CA, USA), and means were compared using Duncan's test considering P<0.05 as the signif icance threshold.
3. Results
3.1. Cold treatment accelerates bolting and f lower bud emergence in f lowering Chinese cabbage
To elucidate the effects of low temperature and the duration of cold treatment on f lowering and bolting in f lowering Chinese cabbage, we counted the days from the end of temperature treatments to the f irst macroscopic appearance of internodes (bolting) and f lower buds under 2, 7.5, and 15°C temperature treatments, each of which lasted for 1, 3, or 5 days. The results showed that the period until bolting ranged from 27 to 35 days (Fig. 1-A). The shortest time were recorded for plants chilled at 2, 7.5, and 15°C for 5 days and the longest were noted for the plants subjected to room temperature treatment or low-temperature treatments for 1 day. Generally, the time to bolting tended to decrease with an increase in the duration of low-temperature treatment. Time to f lower bud visibility was similarly affected (Fig. 1-B), the only difference being that there were signif icant differences between room temperature-treated and 2°C/ 1 day-treated plants.
3.2. Cold treatment alters f lower stalk growth characteristics
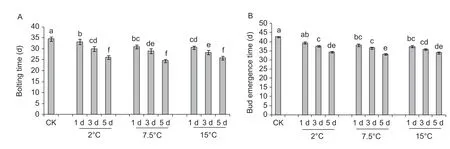
Fig. 1 Bolting (A) and f lower bud emergence (B) time in f lowering Chinese cabbage under cold (2, 7.5, and 15°C) and control (CK, room temperature 25°C) treatments for 1, 3, or 5 days. Statistically signif icant differences in mean values at different sampling points are indicated by different letters (Duncan's test, P<0.05). Bars are SE (n=4).
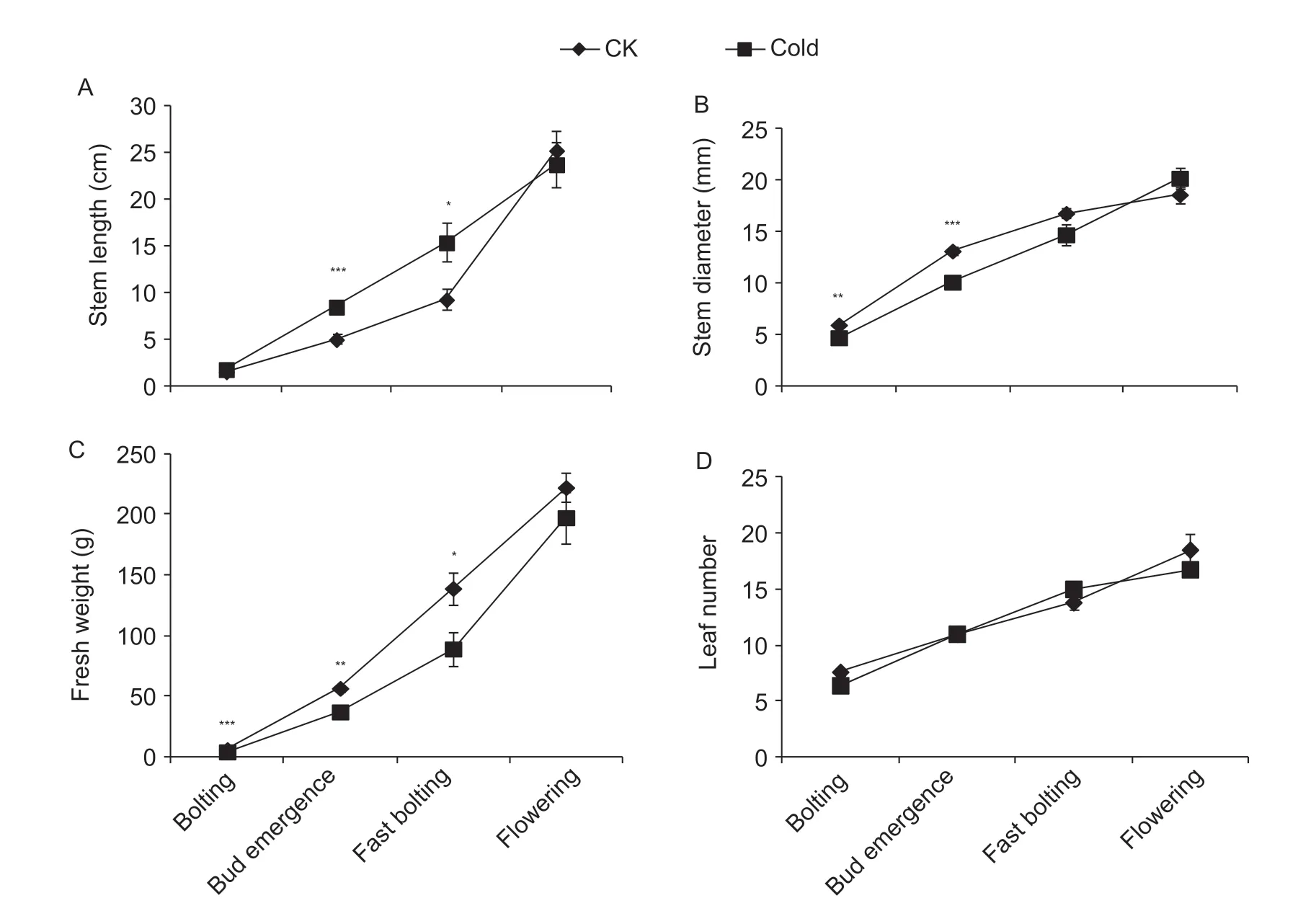
Fig. 2 Physiological measurements of f lower stalks of f lowering Chinese cabbage at bolting, f lower bud emergence, fast bolting, and f lowering stages under 15°C (5 days) and control (CK, room temperature 25°C) treatments. A, stem length. B, stem diameter. C, fresh weight. D, leaf number. *, P<0.05; **, P<0.01; ***, P<0.001 (Student's t-test). Bars are SE (n=4).
As shown in Fig. 2, stems were longer in the cold-treated plants at the fast bolting and f lowering stages, whereas there were no differences at the other two examined stages. Stems were thinner during the f irst two stages (bolting and f lower bud emergence), whereas no signif icant differences were detected at the fast bolting and f lowering stages, which indicates that cold treatment increased f lower stalk length but decreased f lower stalk diameter. Furthermore, we measured the fresh weight and whole leaf numbers at these four stages, and we found that although cold treatment reduced fresh weight production, it had no effect on leaf numbers (Fig. 2).
3.3. Cold treatment alters cell size in shoot apices
As cold treatment shortened bolting time and promoted f lower bud emergence, f lower bud differentiation, which is regulated by plant hormones, may also be affected by cold treatment. We examined this possibility by conducting histological analyses of the shoot apices of f lowering Chinese cabbage under room temperature and cold treatments. Compared with the room temperature treatment, pith cells in transverse sections of shoot apices were smaller at the seedling stage under cold treatment, whereas they were larger when plants had reached the f lower bud emergence stage (Fig. 3-A and C). Under cold treatment, pith cells in the longitudinal sections of shoot apices were smaller than those of room temperature-treated plants at the seedling and f lower bud emergence stages, but they were signif icantly larger at the f lowering stage (Fig. 3-B and D).
3.4. Cold treatment alters hormone contents in shoot apices
As shoot apex differentiation is regulated by plant hormones, we measured the contents of GA, IAA, and CTK in the shoot apices of f lowering Chinese cabbage under cold and control treatments. As shown in Fig. 4, the GA, IAA, and CTK contents were enhanced by cold treatment at the bolting, f lower bud emergence, fast bolting, and f lowering stages, indicating that the promotion of bolting may be attributable to the increased plant hormone contents.
3.5. Cold and exogenous GA treatments have additive positive effects on bolting time
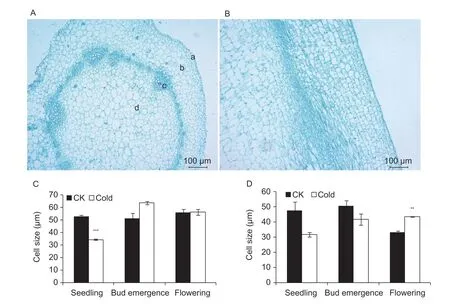
Fig. 3 Histological analysis of shoot apices in cold-treated f lowering Chinese cabbage at the seedling, f lower bud emergence, and f lowering stages. A and B, microscopic images of transverse and longitudinal sections of shoot apices, respectively. a, epidermis; b, endodermis; c, vascular bundle; d, pith cells. C and D, size of pith cells in transverse and longitudinal sections of shoot apices, respectively. CK, room temperature 25°C. **, P<0.01; ***, P<0.001 (Student's t-test). Bars are SE (n=3).
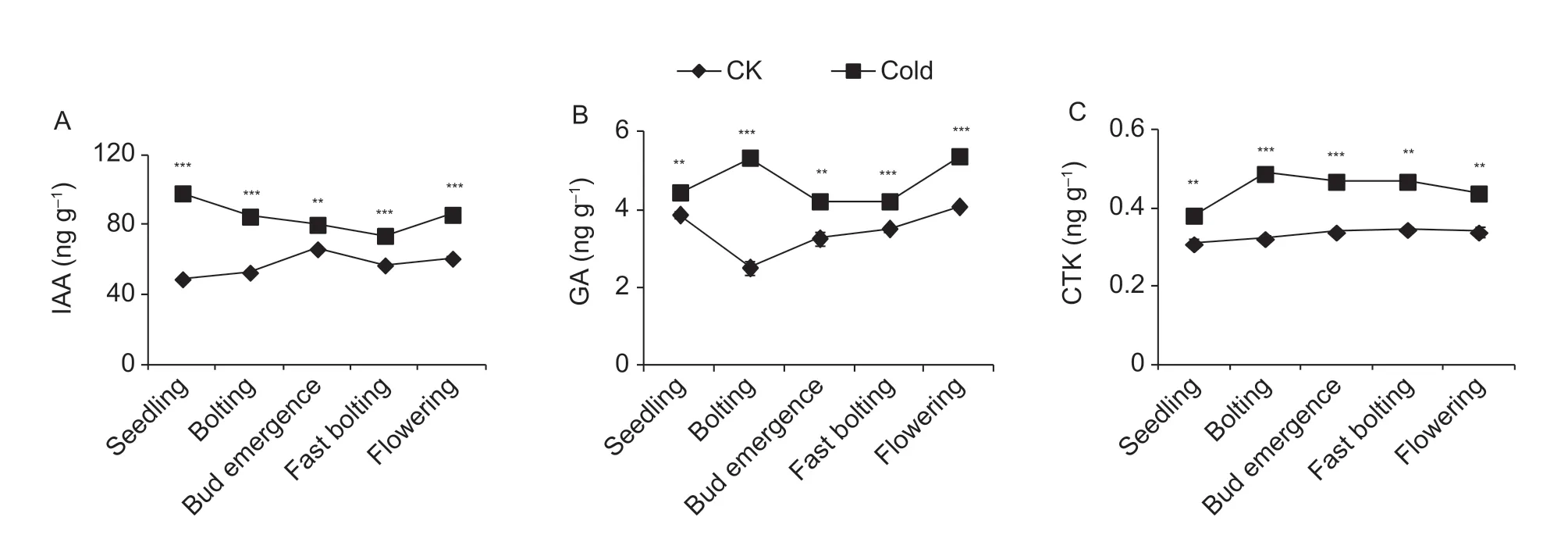
Fig. 4 Hormone contents in shoot apices of f lowering Chinese cabbage under control (CK, room temperature 25°C) and cold treatments. A, indole-3-acetic acid (IAA). B, gibberellin (GA). C, cytokinin (CTK). **, P<0.01; ***, P<0.001 (Student's t-test). Bars are SE (n=3).
GA content in the shoot apices regulates bolting time. Exogenous GA treatment has been shown to induce bolting, whereas the GA synthesis inhibitor uniconazole has the opposite effect. In the present study, similar results were obtained for f lowering Chinese cabbage. At room temperature, all GA-treated plant reached the bolting stage on the 18th day after transplantation, whereas on the same day, less than 40% of the non-treated plants had bolted, and none of the uniconazole-treated plants were found bolting (Fig. 5-A). Under cold treatment, low temperature enhanced the effects of GA, namely, plants started bolting 5 days earlier than at room temperature. All GAtreated plants bolted on the 15th day after transplantation (Fig. 5-D); however, cold treatment did not counteract the effect of uniconazole (Fig. 5-D). In terms of f lower bud emergence, both GA and cold treatments promoted f lower bud emergence, whereas uniconazole treatment inhibited f lower bud emergence only at room temperature (Fig. 5-B and E). However, cold treatment released the uniconazole inhibition of f lower bud emergence (Fig. 5-B and E). Similar results were obtained with respect to f lowering time. Both GA and cold treatments accelerated f lowering time, whereas uniconazole treatment inhibited f lowering only at room temperature (Fig. 5-C and F). The results of GA and uniconazole cotreatment indicated that under both room and low temperature conditions, exogenous GA treatment counteracted the uniconazole inhibition of bolting time, f lower bud emergence, and f lowering time in f lowering Chinese cabbage (Fig. 5).
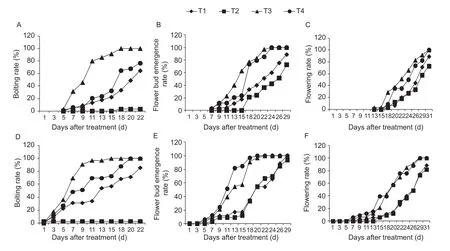
Fig. 5 Effect of cold and exogenous gibberellin (GA) treatments on the growth of f lowering Chinese cabbage. Bolting rate, f lower bud emergence rate and f lowering rate at room temperature (A, B and C, respectively) and cold temperature (D, E and F, respectively; 15°C for 5 days). T1, control (water); T2, uniconazole; T3, GA; T4, GA plus uniconazole.
3.6. Cold treatment and exogenous GA have additive positive effects on f lower stalk growth
Cold treatment induced stem elongation and stem attenuation, and exogenous GA treatment had the same effect on f lower stalks. Flower stalks were longer and thinner after GA treatment, whereas treatment with uniconazole had opposite effects. Under cold treatment, exogenous GA treatment resulted in stems that were longer than those in the control treatment as well as those of GA-treated plants at room temperature (Table 1). These results indicated that low temperature and GA have additive positive effects. With uniconazole treatment under both room and low temperature conditions, the stems were shorter than those in control plants, although there was no difference between cold and room temperature treatments, which indicates that low temperature could not recover the inhibitory effects of uniconazole (Table 1). Similar results were obtained with respect to the stem diameter (Table 1). With regards to fresh weight and leaf number at room temperature or low temperature, GA plus uniconazole treatment induced an effect similar to that observed when using uniconazole alone, i.e., lower fresh weight, whereas GA had no effects on fresh weight and leaf number (Table 1). There was no interaction effect between cold treatment and exogenous GA plus uniconazole treatment on fresh weight and leaf number (Table 1). Compared with room temperature, cold treatment reduced fresh weight and leaf number (Table 1). In addition, similar results were obtained for GA plus uniconazole treatment, with exogenous GA treatment rescuing the uniconazole-induced inhibition of stem length, stem diameter, and fresh weight of f lowering Chinese cabbage under both room and low temperature conditions (Table 1).
4. Discussion
4.1. Cold induces bolting and f lowering in f lowering Chinese cabbage
It is well known that cold affects plant vernalization and bolting, and that different low temperatures have different effects on bolting rates. With an increase in the number of vernalization days, f lower bud differentiation, bolting, and f lowering advance, and bolting rate increases (Wang et al. 2003). In radish and spinach, various low-temperature treatments promoted plant bolting and f lowering in a treatment time-dependent manner (Liu et al. 2010; Li et al. 2014). Similarly, in the present study, we found that all three low-temperature treatments promoted bolting and f lowering in f lowering Chinese cabbage, and that the treatment effect was positively correlated with treatment time. In addition, we detected no difference in bolting and f lowering time among these three low-temperature treatments, in contrast to the f indings of a previous study on Chinese Cabbage that showed that the lower the temperature, the earlier f lower bud differentiation and bolting took place (Hui et al. 2004). These observations indicate that there are no strict low-temperature requirements for bolting and f lowering in f lowering Chinese cabbage, and that a certain degree of cold will promote these processes.
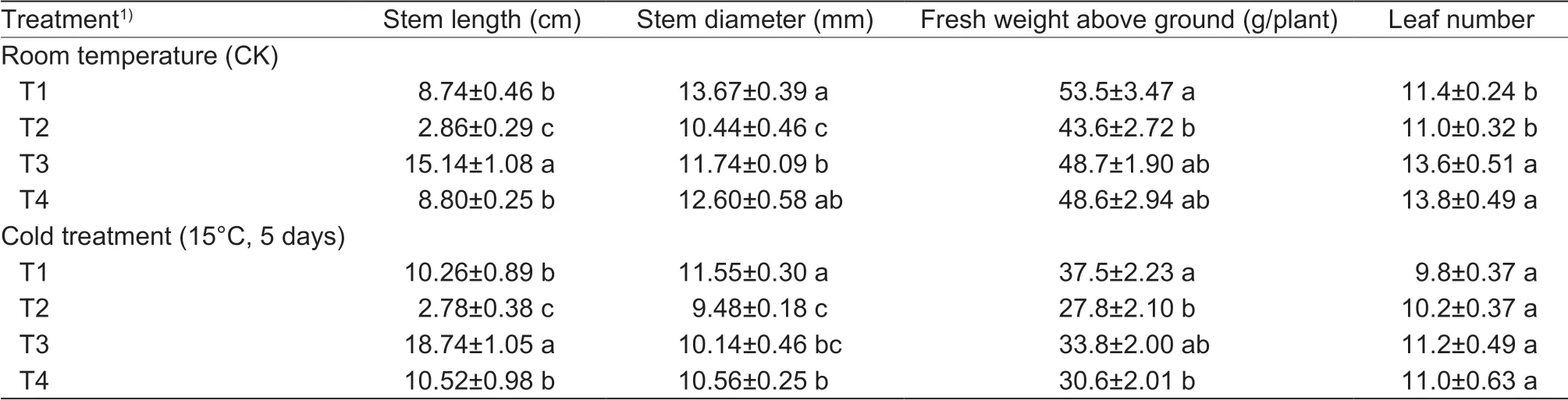
Table 1 Effects of low temperature and gibberellin (GA) treatment on plant growth in f lowering Chinese cabbage
4.2. Cold promotes growth in f lowering Chinese cabbage through mediating plant hormones
Endogenous hormones play an important role in f lower bud differentiation and bolting of vernalized plants (Zanewich and Rood 1995; Dahanayake and Galwey 1999; Tarkowská et al. 2012). Various studies have demonstrated that f lower bud differentiation is under the control of comprehensive regulation mediated by multiple hormones. GA regulates numerous processes during plant development, including seed germination, hypocotyl elongation, and f lower and fruit development (Olszewski et al. 2002; de Jong et al. 2011). In radish, endogenous plant hormone activities are closely related to f lower bud initiation, bolting, and f lowering, and higher GA content can effectively promote f lower bud differentiation and f lowering (Suge and Rappaport 1968; Dahanayake and Galwey 1999; Nie et al. 2016). IAA is also involved in f lower bud initiation and differentiation, and low auxin levels have been detected at the f lower bud initiation stage. Furthermore, auxin inhibits f lower bud differentiation but promotes bolting (Yang et al. 1996; Benková et al. 2003; Goldental-Cohen et al. 2017). In contrast, cytokinins accelerate f lower bud differentiation and bolting time in f lowering Chinese cabbage (Kang et al. 2010). In the present study, we found that cold treatment induced bolting and f lowering in f lowering Chinese cabbage and promoted stem elongation. Furthermore, GA, IAA, and CTK contents in the shoot apices were higher under cold treatment than at room temperature, indicating comprehensive regulation via the actions of these hormones in the control of bolting and f lowering in f lowering Chinese cabbage.
4.3. GA is the main regulator controlling bolting and f lowering in f lowering Chinese cabbage
GA is widely considered as a growth promoter, particularly with respect to stem and cell elongation (Olszewski et al. 2002). The Arabidopsis kao1/kao2 double mutant displays a severe dwarf phenotype and delayed f lowering time, which can be rescued by spraying of exogenous GA (Regnault et al. 2014). Exogenous GA treatment has also been shown to accelerate bolting, f lowering time, and stem elongation in Clivia and Chinese Brassica rapa (Sun et al. 2008; Wang et al. 2009). In the present study, exogenous GA treatment promoted bolting, f lowering, and f lower bud emergence in f lowering Chinese cabbage. GA content in the shoot apices of f lowering Chinese cabbage treated with GA was previously found to be higher than that in control plants receiving only water (Lei et al. 2016). In the current study, cold treatments had effects similar to those observed with GA treatment, with low temperature increasing the GA content in the shoot apices. Accordingly, low temperature and GA have a positively interactive effect on bolting and stem elongation in f lowering Chinese cabbage. Collectively, these results indicate that low temperature promotes bolting and f lowering in f lowering Chinese cabbage via GA induction.
Uniconazole, a gibberellin synthesis inhibitor, is known to inhibit cell elongation, stem elongation, and plant growth (Izumi and Oshio 1991; Kang et al. 2010). Exogenous uniconazole treatment inhibits bud emergence and bolting in radish (Dahanayake and Galwey 1999). Consistent with the uniconazole effect on radish, we found that uniconazole reduced stem elongation and bolting rate in f lowering Chinese cabbage. At both room and low temperature, treatment with uniconazole was observed to reduce the GA content in shoot apices. Moreover, uniconazole inhibited the cold-induced promotion of bolting, whereas cotreatment with GA counteracted the inhibition of bolting, f lowering, and bud emergence. These results indicate that uniconazole inhibits f lowering, bolting, and stem elongation in f lowering Chinese cabbage through reducing GA synthesis, and that this inhibition is stronger than that observed at low temperature.
5. Conclusion
Cold and GA are the two main factors inf luencing bolting and f lowering in f lowering Chinese cabbage. Cold and GA both accelerated plant bolting and f lowering through inducing the GA content in the shoot apices. Moreover, cold and GA co-treated plants displayed additive positive effects. Uniconazole, GA synthesis inhibitor, had a negative effect on plant f lower stalk elongation and f lowering under both room and low temperatures. Meanwhile, GA and cold treatments have different effects on this inhibition implying that cold accelerates bolting and f lowering not only through inducing GA in shoot apices, but also through other ways. Overall, This study provided a fundamental research for revealing the regulatory mechanism of bolting and f lowering in f lowering Chinese cabbage.
Acknowledgements
This work was supported by the earmarked fund for China Agriculture Research System (CARS-25-C-04) and the Natural Science Foundation of Guangdong, China (2016A030313399).
 Journal of Integrative Agriculture2019年5期
Journal of Integrative Agriculture2019年5期
- Journal of Integrative Agriculture的其它文章
- Characterization of TaCOMT genes associated with stem lignin content in common wheat and development of a gene-specif ic marker
- Phenotypic characterization and genetic mapping of the dwarf mutant m34 in maize
- Morphological diversity and correlation analysis of phenotypes and quality traits of proso millet (Panicum miliaceum L.) core collections
- Field identif ication of morphological and physiological traits in two special mutants with strong tolerance and high sensitivity to drought stress in upland rice (Oryza sativa L.)
- Foliar spraying of aqueous garlic bulb extract stimulates growth and antioxidant enzyme activity in eggplant (Solanum melongena L.)
- Seedling and adult plant resistance to leaf rust in 46 Chinese bread wheat landraces and 39 wheat lines with known Lr genes