
Carbon cycle in response to residue management and fertilizer application in a cotton f ield in arid Northwest China
2019-05-10 06:13ZHANGPengpengXUShouzhenZHANGGuojuanPUXiaozhenWANGJinZHANGWangfeng
ZHANG Peng-peng, XU Shou-zhen, ZHANG Guo-juan, PU Xiao-zhen, WANG Jin, ZHANG Wangfeng
1 The Key Laboratory of Oasis Eco-agriculture, Xinjiang Production and Construction Corps, Agricultural College, Shihezi University, Shihezi 832003, P.R.China
2 College of Life Science, Shihezi University, Shihezi 8320003, P.R.China
3 Wulanwusu Agro-meteorological Experiment Station of Xinjiang, Shihezi 832003, P.R.China
Abstract Understanding the inf luence of farming practices on carbon (C) cycling is important for maintaining soil quality and mitigating climate change, especially in arid regions where soil infertility, water def iciency, and climate change had signif ciantly inf ulenced on agroecosystem. A f ield experiment was set up in 2009 to examine the inf luence of residue management and fertilizer application on the C cycle in a cotton f ield in the Xinjiang Uygur Autonomous Region of Northwest China. The study included two residue management practices (residue incorporation (S) and residue removal (NS)) and four fertilizer treatments (no fertilizer (CK), organic manure (OM), chemical fertilizer (NPK), chemical fertilizer plus organic manure (NPK+OM)). Soil organic carbon (SOC) and some of its labile fractions, soil CO2 f lux, and canopy apparent photosynthesis were measured during the cotton growing seasons in 2015 and 2016. The results showed that SOC, labile SOC fractions, canopy apparent photosynthesis, and soil CO2 emission were signif icantly greater in S+NPK+OM (residue incorporation+chemical fertilizer) than in the other treatments. Analysis of all data showed that canopy apparent photosynthesis and soil CO2 emission increased as SOC increased. The S+OM (residue incorporation+organic manure) and S+NPK+OM treatments were greater for soil C sequestration, whereas the other treatments resulted in soil C loss. The S+NPK treatment is currently the standard management practice in Xinjiang. The results of this study indicate that S+NPK cannot offset soil C losses due to organic matter decomposition and autotrophic respiration. Residue return combined with NPK fertilizer and organic manure application is the preferred strategy in arid regions for increasing soil C sequestration.
Keywords: residue, fertilizer, SOC pool, canopy apparent photosynthesis, soil respiration, soil C balance
1. Introduction
Achieving sustainable global food security and mitigating global climate change are serious challenges in the 21st century (Lal 2004, 2013a; West et al. 2014). The common link between these two challenges is the global carbon (C) cycle. The terrestrial C pool, which is an important component of the global C cycle, is greatly inf luenced by anthropogenic activities (Lal 2009; Wu et al. 2015). Increasing soil organic carbon (SOC) stocks for all of agricultural areas by 4 per mille (about 2.5 Gt C yr-1) can offset about 30% of global greenhouse gases emission (Minasny et al. 2017). Greater soil C sequestration could also result in signif icant increases in crop yields (Lal 2013b). Hence, increasing soil C sequestration is a win-win strategy for enhancing crop yield and mitigating climate change.
SOC, which accounts for about 62% of the global soil C pool (Lal 2004), affects soil quality and agronomic productivity by inf luencing soil physical, chemical and biological properties (Zhu et al. 2015). The accumulation of SOC in farmland depends on the balance between inputs (e.g., addition of crop residue and manure) and outputs (i.e., release of CO2due to organic matter decomposition) (Lal 2009; Bhattacharyya et al. 2011; Kou et al. 2012). Human activity greatly inf luences the SOC pool in farmland (Zhang et al. 2014; Abdalla et al. 2015). Soil may be either a C sink or a C source, depending on climate, crop species, initial SOC content, and long-term management practices (e.g., tillage, fertilizer application, residue return) (Feng et al. 2011; Qiao et al. 2015; Saikia et al. 2015; Wu et al. 2015).
Soil respiration (i.e., the release of C from soil as CO2) is controlled by a range of biotic factors (e.g., substrate supply, plant productivity, and microbial properties) and abiotic factors (e.g., soil temperature, moisture, and O2supply) (Han et al. 2007, 2014; Gomez-Casanovas et al. 2012; Huang et al. 2012; Zhang et al. 2014; Gong et al. 2015). As the second largest C f lux between terrestrial ecosystems and the atmosphere, soil respiration annually returns about 80 Pg of C to the atmosphere (Raich et al. 2002). Small changes in soil respiration can have great effect on atmospheric CO2concentrations and soil C sequestration. Potential positive feedback between increasing temperature and enhanced soil respiration may ultimately accelerate global warming (Han et al. 2007; Huang et al. 2012; Gong et al. 2014). Farming practices inf luence soil respiration by altering both autotrophic and heterotrophic respiration. These changes may have positive or negative inf luence on soil C sequestration and climate change (Gomez-Casanovas et al. 2012; Fan et al. 2015). The inf luence of farming practices on soil respiration must be quantif ied in order to estimate soil C emission from farmland.
Photosynthesis is an important component of the global C cycle. Photosynthesis is strongly inf luenced by both anthropogenic activities and climatic factors (Huang et al. 2012; Fan et al. 2015). The f ixation of atmospheric CO2by plants sequesters organic C in above and below ground biomass. The return of crop residue to soil increases soil C input (Lal 2009; Saikia et al. 2015). Lal (2013b) reported that crop biomass production increased as photosynthesis increased, resulting in greater soil C sequestration. Therefore, the adoption of management practices which increase photosynthesis is considered a viable option for enhancing soil C sequestration and mitigating atmospheric CO2. Some studies have shown that higher photosynthetic rates and more C input into the soil do not always increase soil C sequestration (Gomez-Casanovas et al. 2012; Zhang et al. 2014). This, in part, may be the consequence of a recently documented link between concurrent photosynthesis and variations in soil respiration rates (Huang et al. 2012; Han et al. 2014; Gong et al. 2015). Therefore, greater understanding about the relationship between photosynthesis and soil respiration is important for regulating the C cycle in farmland ecosystems.
The Xinjiang Uyghur Autonomous Region is not only an important cotton (Gossypium hirsutum L.) producing area, but also the largest arid area in China. Cotton production in Xinjiang increased signif icantly in the 1990s due to the adoption of agricultural practices such as plastic f ilm mulch, drip irrigation, residue incorporation, NPK fertilizer application, and green manure. Most cotton producers in Xinjiang currently return cotton residue to the soil and apply large amounts of chemical fertilizer (440 kg N ha-1, 420 kg P2O5ha-1, and 270 kg K2O ha-1). The livestock industry in Xinjiang has also grown in recent years, resulting in the production of large amounts of manure. Therefore, manure application to farmland not only makes use of an important agricultural resource but also promotes integration between the crop and livestock industry. Although many researchers have studied the effect of farming practices on C cycling in farmland (Ding et al. 2007; Fan et al. 2015), information is still limited on the systematic study of the C cycle under long-term current farming practices, particularly in grey desert soil in arid regions of Northwest China. The goal of this f ield experiment was to assess the C cycle in a cotton f ield in arid Northwest China after six years of different residue management and fertilization practices. The specif ic objective was to evaluate the inf luence of residue return, fertilizer application and their combination on soil physiochemical properties, plant productivity, soil respiration, and soil C balance.
2. Materials and methods
2.1. Site details
The f ield experiment was set up in 2009 at the Wulanwusu Agrometeorological Experiment Station in the Xinjiang Uygur Autonomous Region of Northwest China (44°17´N, 85°49´E). The site has an arid climate, with mean annual precipitation of 211 mm, mean annual evaporation of 1 664 mm, and mean annual temperature of 7.0°C. The annual frost-free period is 170 days. The soil texture is sandy loam and is classif ied as a Calcaric Fluvisol according to the FAO/WRB (FAO 1998). Table 1 shows selected soil properties before the start of the experiment in 2009. Fig. 1 shows the daily maximum temperature, minimum temperature, and precipitation amount at the experimental site during the 2015 and 2016 growing seasons.
2.2. Experimental design
The experimental design was split-plot with three replications. Each plot was 3 m×8 m. The main plot treatments were: (i) residue incorporation (S) and (ii) no residue incorporation (NS). The split-plot treatments were: (i) no fertilizer (CK); (ii) manure (OM); (iii) N, P, and K fertilizer (NPK); and (iv) N, P, and K fertilizer plus manure (NPK+OM). The same treatment was applied to its respective plot each year.
The chemical fertilizers were urea (46% N), ammonium dihydrogen phosphate (18% N and 46% P2O5), and sulphate-potassium magnesium (22% K2O, 5% Mg and 14% S). According to current agricultural fertilizer application rate in Xinjiang, the chemical fertilizer amounts were 440 kg N ha-1, 420 kg P2O5ha-1, and 270 kg K2O ha-1in this study. Of these amounts, 30% of the N, 70% of the P2O5, and 100% of the K2O were applied 10 days before planting each year. The remainder of the fertilizer (i.e., 70% of the N and 30% of the P2O5) was applied as topdressing via the irrigation system. The manure was production from Xinjiang Tianwu Ecological Technology Co., Ltd. (China), which consisted of composted poultry manure applied at the rate of 30 t ha-1in dry matter on 10 days before sowing. The 30 t manure contained 5 181 kg organic C, 534 kg total N, 411 kg total P, and 654 kg total K. Table 2 shows the mean soil C input differed signif icantly between the mineral and organic manure treatments, especially between residue incorporation and no residue incorporation. Therefore both residue incorporation and organic manure treatments showed higher mean soil C input rates in comparison with no residue incorporation and mineral fertilizer, respectively.
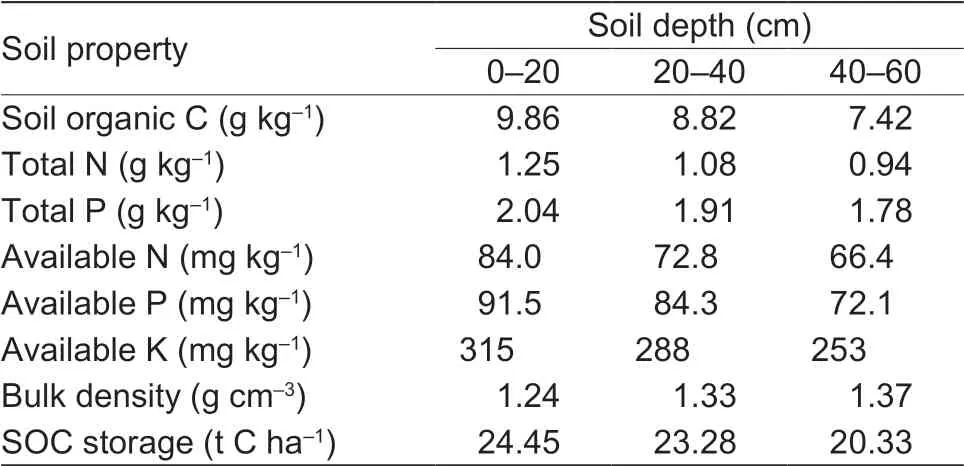
Table 1 Initial soil properties at the experimental site in 2009
Plastic f ilm (60 cm width and 1.57 mm thickness) was completely buried between the plots before planting to prevent the movement of water and fertilizer between plots. The combination of plastic f ilm mulch and drip irrigation is widely used for cotton production in Xinjiang. Therefore, the plots were covered before sowing with 1.1-m-wide sheets of transparent plastic f ilm, with a 0.4-m-wide strip of bare soil between each sheet. There were two sheets of plastic f ilm per plot. Two drip-irrigation tubes were laid on the soil surface beneath the plastic f ilm with a distance of 0.8 m between the tubes. The emitters were 0.4 m apart. Specif ic information about the irrigation dates, irrigation amounts, and topdress fertilizer amounts are shown in Table 3. On 30-40 days after harvesting, the residues are always cut by machine with 10-15 cm height and ploughed to a depth of 20-25 cm. On 10 days before sowing, all plots are always rotary tillage using a single pass of tined stubble cultivator at a depth of 7-10 cm. These tillage practices in this study were typical in Xinjiang, which were used since the beginning of the experiment in 2009.
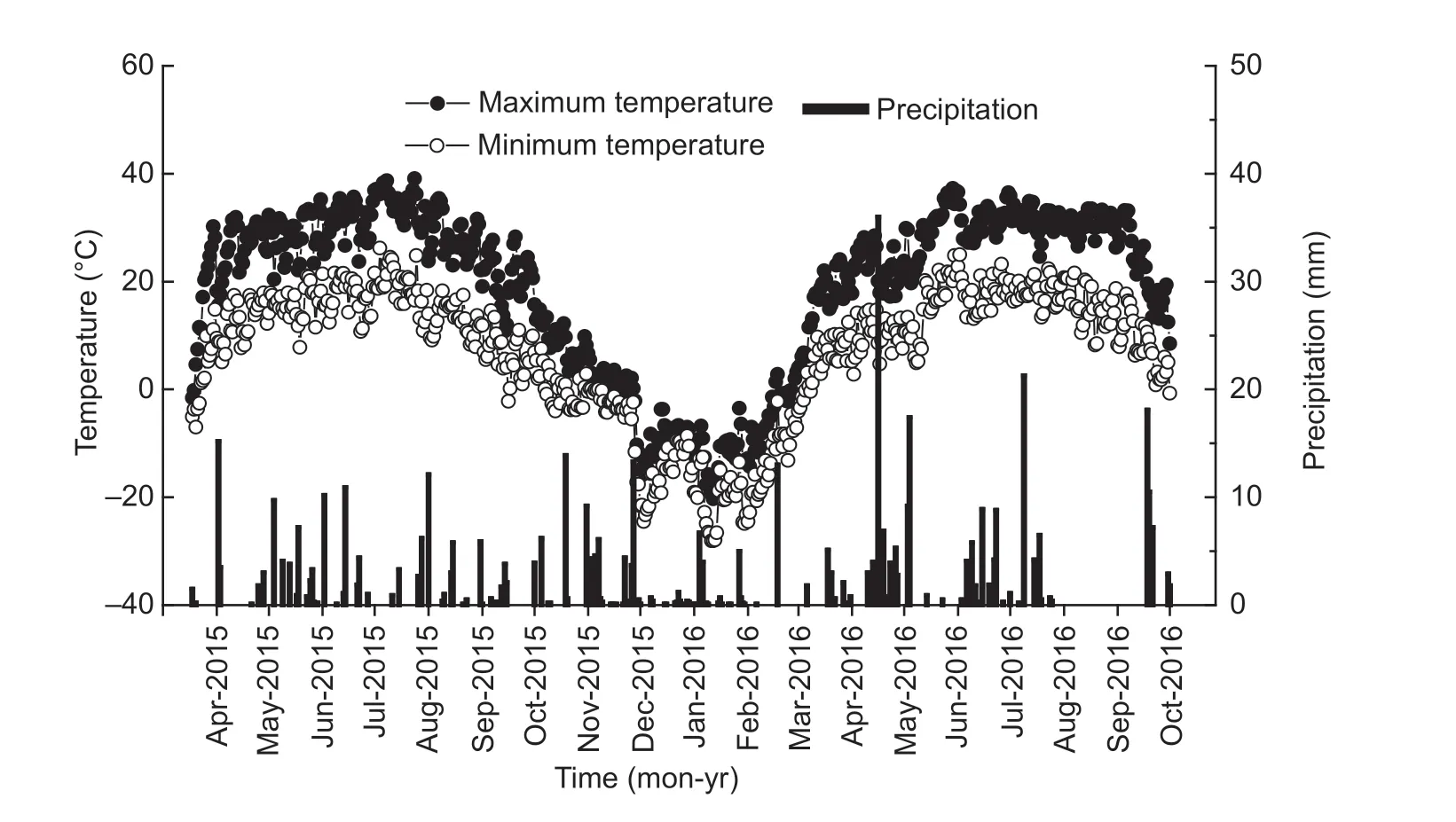
Fig. 1 Minimum and maximum temperature and precipitationat the Wulanwusu Agro-meteorological Experiment Station of China in 2015 and 2016.
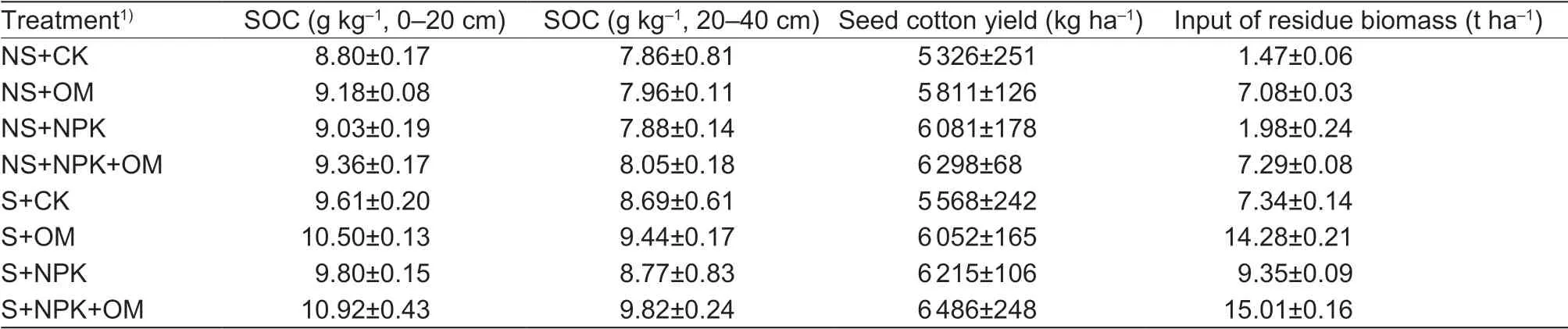
Table 2 Mean soil organic C (SOC), seed cotton yield and soil C input (residue and organic manure) (from 2009 to 2014) under different residue management and fertilizer treatments
Cotton seeds (cultivar Xinluzao 59, a common cultivar in the study area) were sown through holes in the plastic f ilm mulch on 19 April, 2015 and 15 April, 2016, which is the type of crop from 2009 to 2014. The plant density was 2.05×105plants ha-1. Each sheet of plastic f ilm mulch had four rows of cotton. The spacing between the rows was 30, 50 and 30 cm. The distance between plants within a row was 0.15 m. This row spacing is common in the region. The planting pattern was the same in both years. The cotton plants were top pruned at the full f lower stage (12 July in both years). The cotton was harvested on 1 October, 2015 and 30 September, 2016.
2.3. Soil CO2 emission measurements
Soil respiration rates were measured approximately every 15 days in the entire year (May 2015-October 2016) using an LI-8100 Automated Soil CO2Flux System (Li-COR Inc., Lincoln, NE, USA). Measurements were taken between 11:00 to 17:00 on the each day, as the base of taking into account diurnal changes in temperature. To minimize disturbance to the soil surface, the LI-8100 respiration chamber was mounted on polyvinyl chloride (PVC) pipes (11.5 cm height and 20 cm inner diameter) that were installed on the day before the f irst measurement and left in the soil during the experiment. There were three collars which were installed to a depth of 9.5 cm in the bare soil between the two sheets of plastic f ilm mulch in each plot. The soil respiration measurements were conducted in the same plot sequence on each sampling date to avoid systematic errors. Cumulative soil CO2emission was calculated based on the soil respiration rate (1 µmol CO2m-2s-1=10.368 kg C ha-1d-1) using the following equation:

Table 3 Irrigation date, irrigation amount, and fertilizer amount applied during the cotton growing season
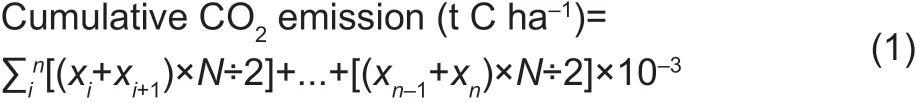
Where, i is the date of the f irst CO2measurement, n is the date of the next CO2measurement, x is the CO2f lux rate (kg ha-1d-1), and N is the number of days between two consecutive soil CO2f lux measurements.
2.4. Canopy apparent photosynthesis
Canopy apparent photosynthesis (CAP) was determined using the assimilation chamber method according to Acock et al. (1978) and Reddy et al. (1995) at the following stages: (i) full square stage (FS, 65-70 days after sowing); (ii) early full f lower stage (EFF, 75-80 days after sowing); (iii) full f lower stage (FF, 85-90 days after sowing); (iv) early full boll stage (EFB, 95-100 days after sowing); (v) late full boll stage (LFB, 105-110 days after sowing); and (vi) boll open stage (BO, 115-120 days after sowing). The assimilation chamber (90 cm long×976 cm wide×110 cm high) was covered with acrylic f ilm, which transmitted more than 95% of the solar radiation. Two fans were installed inside the chamber to mix the air. Air temperatures within the chamber were less than 3°C above ambient.
The CAP measurements were made between 12:30 and 13:30 on clear, windless days. The chamber was placed over two rows in the center of each plot. There was a 30-cmwide space between the two cotton rows. Two assistants held the chamber tightly against the plastic f ilm mulch to prevent air leakage around the bottom of the chamber. The CO2concentration inside the chamber was determined with a LI-840A CO2/H2O Analyzer (LI-COR Inc., Lincoln, NE, USA). The gas exchange rates in each plot were measured during at least three 60-s intervals. We began recording the values when the CO2concentrations inside the chamber began to drop steadily.
After measuring CAP, the plants within the chamber were cut off at ground level and removed. The chamber was returned to its original position and the gas exchange measurements were repeated to determine soil respiration. The CAP values were corrected to account for soil respiration.
2.5. Net primary productivity and yield
The shoot biomass and root biomass of the cotton were measured on the same day as CAP. Shoot biomass was determined using f ive plants (clipped at the cotyledonary node) from each plot. Root biomass was determined using a soil column cylinder auger (Eijkelkamp, Giesbeek, The Netherlands) to excavate f ive soil cores (15 cm diameter) from the 0-20 cm soil depth in each plot, because more than 52.4% of cotton plants are distributed in the soil layer of 0-20 cm, and 80.6% of the root is in the range of 0-15 cm along both sides of cotton plants (Li et al. 2000). The cores were collected directly beneath the plants that had just been removed. The soil was carefully picked apart and the roots were removed by hand.
The shoot and root samples were oven-dried at 105°C for 30 min and then at 80°C for 48 h. The samples were weighed after oven-drying and then the average biomass of each plant was calculated. The total biomass per unit land area was calculated by multiplying the individual plant biomass (maximum aboveground and root biomass each year) by the plant density. Total seed cotton yield was determined by hand harvesting all of the plants in a 24 m2area within each plot. Soil C input (Cinput, t ha-1) was estimated based on the amount of shoot residue (Cshoot, t ha-1), root residue (Croot, t ha-1), and manure (Cmanure, t ha-1) that was incorporated into the soil in each treatment:

Where, Mshoot(t ha-1) and Mroot(t ha-1) are the maximum shoot and root biomass, respectively. The values 0.45 (g g-1) and 172.7 (g kg-1) are the C content of the residue and poultry manure, respectively (Zhang et al. 2014). The value 0.524 and 0.806 are the ratio of root biomass in the 0-20 cm depth and in the range of 0-15 cm along both sides of cotton plants to total root biomass, respectively (Li et al. 2000).
Soil C balance was calculated based on soil C input and cumulative CO2emission:

2.6. Soil sampling and analyses
Previous results showed that SOC and labile C fractions are changed on the whole growing season (Xu et al. 2014; Gong et al. 2015; Saikia et al. 2015). In this study, soil samples and soil bulk density were collected from f ive random points within each plot at the following times in 2015 and 2016: (i) before sowing (BS, 5 days before sowing); (ii) full f lower (FF, 85-90 days after sowing); and (iii) harvest time (HT, 5 days before harvesting). Soil samples were collected from the 0-20 and 20-40 cm soil depths at each sampling point using a soil drill (5 cm diameter×20 cm depth). Samples from the same depth within each plot were thoroughly mixed and divided into two parts. The f irst part of each sample was stored in a refrigerator at 4°C for determination of microbial biomass C (MBC). The second part of each sample was air-dried for chemical analysis.
Soil bulk density (BD) in the 0-20 and 20-40 cm depth was determined by using soil core (7.5 cm diameter×7.5 cm depth). The cores were over dried at 105°C for 48 h and bulk density was calculated by dividing the weight of the dried soil by the volume of the core. The SOC was determined by the Walkley-Black method (Walkley and Black 1934). Brief ly, 10 mL of 1 mol L-1K2Cr2O4+20 mL of concentrated H2SO4were add to 0.1 g soil sample, which were then treated at 178-180°C for 10 min. Finally, the excess dichromate was titrated with 0.5 mol L-1FeSO4. The MBC was determined by the chloroform fumigation extraction method (Jenkinson and Ladd 1981). The soil sample was incubated with chloroform for 24 h. At the end of incubation, soil sample was extracted with 0.5 mol L-1K2SO4(2:5 of soil:extraction ratio (w/v)) in tubes for 1 h on an end-over-end shaker. Then all the supernatant was f iltered into separate vials for MBC analysis. The MBC was measured using the same methods used for SOC. Labile C (LBC) was measured by the KMnO4-oxidation method (Blair et al. 1995). The soil sample (containing 15 mg C) was extracted with 25 mL of 333 mmol L-1KMnO4in tubes for 1 h on an end-over-end shaker. Then the tubes were centrifuged for 5 min and the supernatants diluted 1:250 with distilled water. The LBC was determined by the absorbances of the diluted samples and standards on a split beam spectrophotometer at 565 nm. Water extractable organic C (WSOC) and hot-water extractable organic C (HWSOC) were determined using methods described by Ghani et al. (2003). The soil sample (3 g) was extracted with 30 mL of distilled water for 30 min on an end-over-end shaker, centrifuged for 20 min and all the supernatant from was f iltered through 0.45-µm membrane f ilter into separate vials for WSOC analysis. A further 30 mL distilled water was added in the same tubes and the tubes were capped and left for 16 h in a hot-water bath at 80°C. Then HWSOC was collected by f iltering supernatants.
The WSOC and HWSOC were measured using the same methods used for SOC.
2.7. SOC storage
The SOC storage (0-40 cm depth) was estimated using the following equations (Zhang et al. 2012):
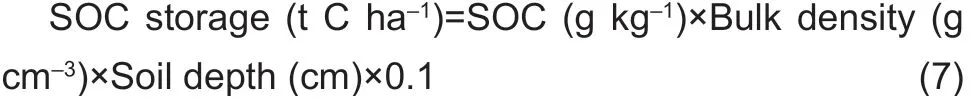
2.8. Statistical analysis
Statistical analysis was completed using the SPSS (22.0) software package for Windows. Two-way ANOVA using general linear model (GLM) procedures was used to test the main and interactive effects of residue management and fertilizer application. The least signif icant difference (LSD) at the 0.05 level of probability was used to compare means. Correlation analyses were used to test the relationship between each pair of variables. Regression analysis was conducted when the correlations were signif icant. All f igures were prepared with Sigmaplot 12.5.

Table 4 Soil bulk density (BD, g cm-3) under different residue management and fertilizer treatments in 2015 and 20161)
3. Results
3.1. Soil bulk density
Soil bulk density increased as soil depth increased in both growing seasons (Table 4). The soil bulk density also increased between sowing and harvest. The soil bulk density ranged between 1.19 and 1.56 g cm-3. Fertilizer application signif icantly reduced soil bulk density. Residue management generally had no signif icant effect on bulk density. The exception was in the 20-40 cm depth at harvest. Residue×fertilizer interaction signif icantly affected soil bulk density in the 20-40 cm depth at full f lowering and at harvest both years. Six years of S+CK and NS+CK treatments signif icantly increased bulk density in the 20-40 cm depth by 0.08 and 0.07 g cm-3(6.02 and 5.26%) (Tables 1 and 4). In c omp arison, NPK+OM treatment had a reducing tread in the 0-20 cm and 20-40 cm depth.
3.2. SOC and organic C fractions
The SOC and organic C fractions decreased as soil depth increased in both growing seasons (Table 5). The SOC and organic C frac tions changed greatly during growing seasons, especially WSOC had the greatest change range, which increased from sowing to f lowering and then declined. Fertilizer application signif icantly affected SOC and its labile fractions in both soil depths in 2015 and 2016. Resid ue management and residue×fertilizer interaction signif icantly affected WSOC in both soil depths. The effects of residue management and residue×fertilizer interaction on SOC, LBC, MBC, and HWSOC were inconsistent.
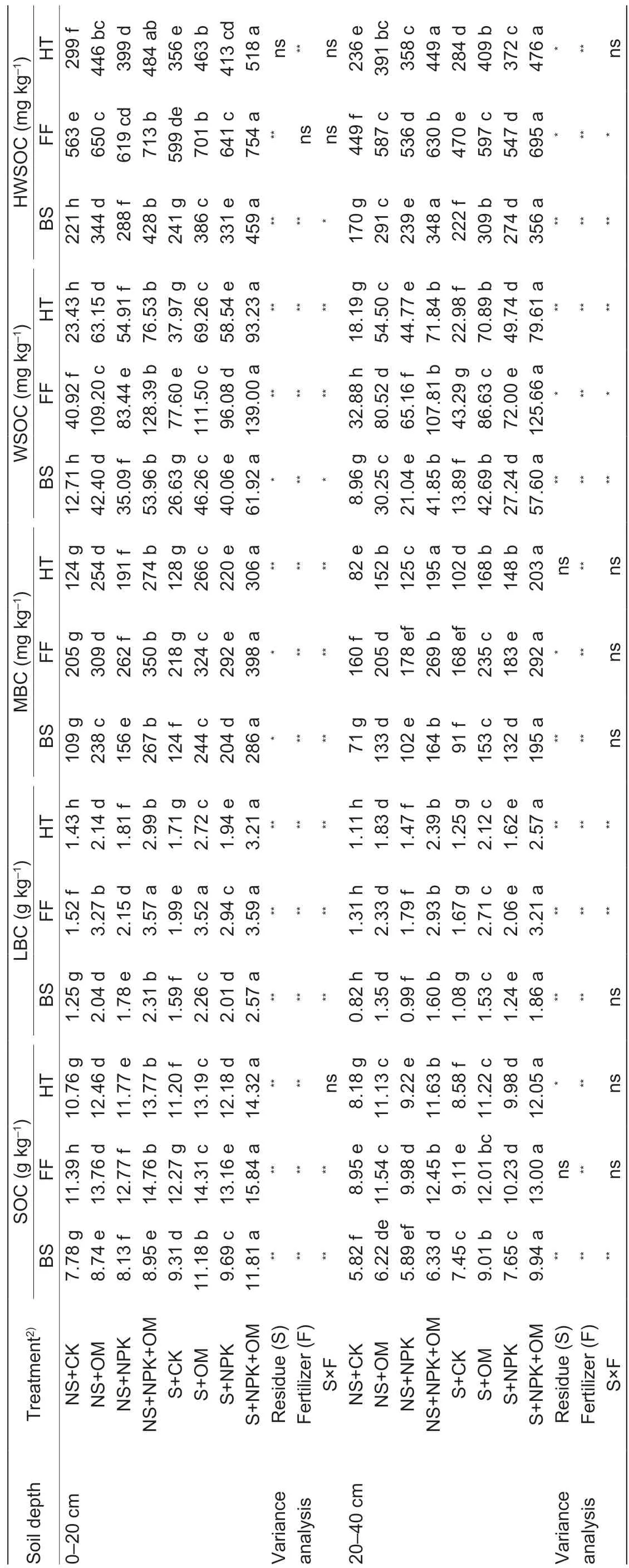
Table 5 Mean soil organic C(SOC), labile C (LBC), microbial biomass C (MBC), water soluble organic C (WSOC), and hot water soluble organic C (HWSOC) as affected by the residue management and fertilizer treatments in 2015 and 20161)
Residue incorporation signif icantly increased SOC by averages of 10.89% compared with no residue incorporation (Table 5). Fertilization also signif icantly increased SOC, and there were signif icant differences among the fertilizer treatments. Regardless of residue management, SOC declined in the order NPK+OM>OM>NPK>CK. The LBC, MBC, WSOC and HWSOC fractions decreased in the same order as SOC. Compared with CK, NPK+OM signif icantly increased SOC by 43.40%.
Six years of S+OM and S+NPK+OM treatments signif icantly increased SOC in the 0-20 cm depth by 1.32 and 1.95 g kg-1(13.35 and 19.82%), and in 20-40 cm depth by 0.19 and 1.12 g kg-1(2.19 and 12.66%), respectively (Tables 1 and 5), compared 2009 with present. In comparison, S+NPK decreased SOC by only 0.17 g kg-1(1.69%) in 0-20 cm depth, and 1.17 g kg-1(13.21%) in 20-40 cm depth.
3.3. SOC storage
Six years of residue return had signif icantly increased SOC storage (0-40 cm) (Table 1 and Fig. 2). All of the fertilized soils had greater SOC storage than unfertilized soil (CK) in the 0-40 cm soil depth. The NPK+OM treatment had the greatest SOC storage (45.80-72.49 t ha-1) among all treatments. The SOC storage in CK was 39.87-58.87 t ha-1. The NPK+OM and OM treatments increased SOC storage 13.04-19.30% and 7.11-12.66% than NPK, respectively.
3.4. Soil CO2 emission
Soil CO2f lux was measured from May 2015 to October 2016. Soil CO2f lux was the greatest during the summer months and the least during the winter (Fig. 3-A). The soil CO2f lux decreased signif icantly in the following order: S+NPK+OM>S+OM>NS+NPK+OM>S+NPK>NS+OM>SCK> NS+NPK>NS+CK (Fig. 3-C). There was signif icantly positive linear correlation between average air temperature and soil CO2f lux (Fig. 3-B). It should be noted that there was a spike in soil CO2f lux on 15 November, 2015.
3.5. Canopy apparent photosynthesis
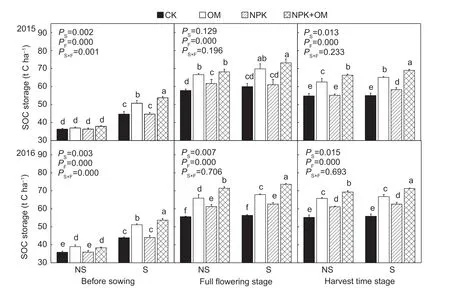
Fig. 2 Soil organic C storage in the 0-40 cm soil depth under different residue management and fertilizer application treatments in 2015 and 2016. NS, no residue incorporation; S, residue incorporation. CK, no fertilizer; OM, manure; NPK, N, P, and K fertilizer; NPK+OM, N, P, and K fertilizer+manure. P S, P F and P S×F stood for the P-value of variance analysis for soil organic carbon storage under the effects of straw, fertilization and their interaction, respectively. Different letters within a crop growth stage represent signif icant differences at P<0.05. Values represent mean±standard deviation (n=3).

Fig. 3 Seasonal changes in soil CO2 f lux (A) (n=6) during the cotton growing and non-growing seasons, the relationship between temperature and CO2 f lux (B) (n=30), average CO2 f lux (C) (n=6) and cumulative CO2 emission (D) (n=6) under different residue management and fertilizer treatments. NS, no residue incorporation; S, residue incorporation. CK, no fertilizer; OM, manure; NPK, N, P, and K fertilizer; NPK+OM, N, P, and K fertilizer+manure. P S, P F and P S×F stood for the P-value of variance analysis for soil organic carbon storage under the effects of straw, fertilization and their interaction, respectively. Values represent mean±SD.
On a ground surface basis, CAP reached a maximum during the full f lower stage and then gradually declined (Fig. 4). The S+NPK+OM treatment had the greatest CAP rate in this study. Residue management signif icantly affected CAP in 2016 but not in 2015. Fertilizer treatment signif icantly affected CAP in both years. The residue×fertilizer interaction was signif icant in 2016 but not in 2015. For the fertilizer treatments, CAP decreased in the order NPK+OM>NPK>OM>CK. The CAP rate in NPK+OM was greater than that in CK, OM and NPK by 55.70-58.98%, 30.96-32.44%, and 13.05-14.93%, respectively.
3.6. Photosynthesis product accumulation and seed cotton yield
Residue management signif icantly affected total cotton biomass (shoot+root) (Fig. 5). Shoot biomass increased rapidly after the full square stage and reached a maximum at the boll open stage. The fertilizer treatments had no signif icant effect on shoot biomass before full f lowering. The differences among treatments steadily increased after full f lowering, when air temperatures and the amount of irrigation water increased. Root biomass reached a maximum at the early full boll stage and then declined in both years. For the fertilizer treatments, maximum shoot biomass and maximum root biomass both decreased in the order NPK+OM>NPK>OM>CK. The fertilizer treatments signif icantly affected seed cotton yield, whereas residue management did not (Table 6). The seed cotton yield in NPK+OM was signif icantly greater than that in CK, OM, and NPK by 19.05-24.95%, 7.57-10.25%, and 3.01-3.73%, respectively.
3.7. Soil C balance
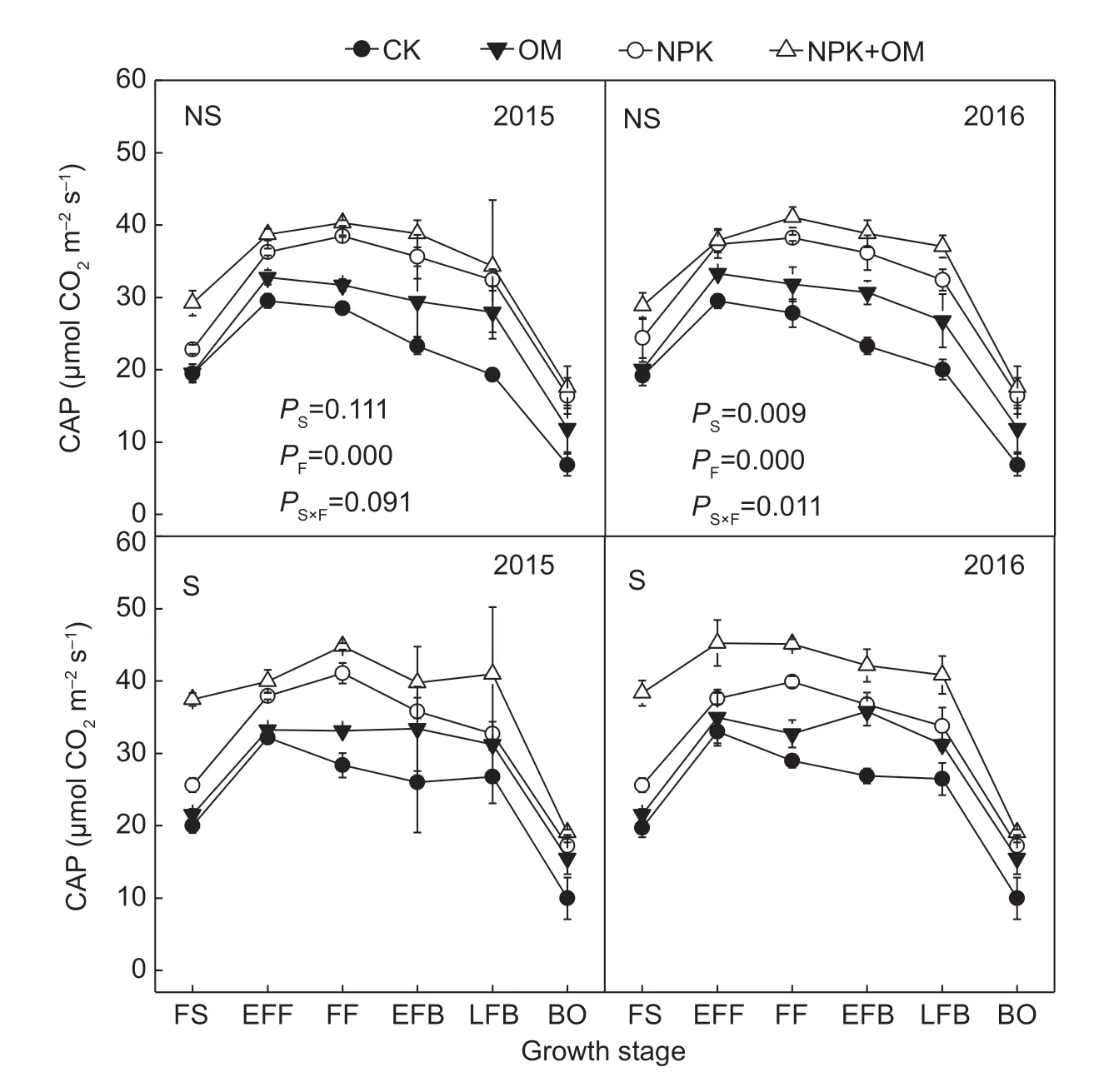
Fig. 4 Canopy apparent photosynthetic rate (CAP) of cotton as affected by residue management and fertilizer treatment during the cotton growing season in 2015 and 2016. NS, no residue incorporation; S, residue incorporation; CK, no fertilizer; OM, manure; NPK, N, P, and K fertilizer; NPK+OM, N, P, and K fertilizer+manure. FS, full square stage; EFF, early full f lower stage; FF, full f lower stage; EFB, early full boll stage; LFB, late full boll stage; BO, boll open stage. P S, P F and P S×F stood for the P-value of variance analysis for soil organic carbon storage under the effects of straw, fertilization and their interaction, respectively. Values represent mean±SD (n=3).
Residue management and fertilizer treatment both signif icantly affected soil C inputs (Table 6). Soil C inputs were 171% greater in the residue treatments than in no residue treatments. Compared with CK, OM, NPK, and NPK+OM increased soil C inputs by 147, 103, and 163%, respectively. Cumulative soil CO2emission between May 1, 2015 and October 1, 2016 decreased in the order S+NPK+OM (24.4 t C ha-1)>S+OM (22.8 t C ha-1)>NS+NPK+OM (21.0 t C ha-1)>S+NPK (19.9 t C ha-1) >NS+OM (19.6 t C ha-1)>S+CK (18.2 t C ha-1)>NS+NPK (17.7 t C ha-1)>NS+CK (16.0 t C ha-1) (Fig. 3-D). Cumulative CO2emission was signif icantly greater in the residue-incorporated treatments than in the no residue treatments. Regardless of residue management, cumulative CO2emission in NPK was signif icantly greater than that in CK, but signif icantly less than that in NPK+OM and OM. Residue management×fertilizer interaction had signif icant inf luence on cumulative CO2emission.
Soil C balance in S+OM and S+NPK+OM were greater than 1 (Fig. 6), indicating that these treatments were C sinks. Soil C balance was greater in the residueincorporated treatments than in the corresponding no residue treatments. The application of OM signif icantly reduced soil C loss compared with CK and NPK. For the fertilizer treatments, soil C balance declined in the order NPK+OM>OM>NPK>CK. Therefore, in this study, residue incorporation plus NPK+OM had the greatest potential C sequestration.
3.8. Correlations among soil C input, SOC, CAP, cumulative CO2 emission and soil C balance
There was a strong, positive linear correlation between soil C input and SOC (R2=0.559, P=0.0330, in 0-20 cm; R2=0.522, P=0.0430, in 20-40 cm) during the growing season (Fig. 7-A). Similarly, there was a signif icantly positive linear correlation between SOC and CAP (R2=0.726, P=0.0072, in 0-20 cm; R2=0.520, P=0.0436, in 20-40 cm) and cumulative CO2emission (R2=0.855, P=0.0010, in 0-20 cm; R2=0.806, P=0.0025, in 20-40 cm) (Fig. 7-B and C). Moreover, there was signif icant positive linear correlation between SOC storage and cumulative CO2emission (R2=0.977, P<0.0001, in 0-20 cm; R2=0.966, P<0.0001, in 20-40 cm) (Fig. 7-D). Cumulative CO2emission was signif icantly correlated with CAP (R2=0.575, P=0.0292) (Fig. 7-E). Soil C balance was signif icantly correlated with soil C input (R2=0.975, P<0.0001) between 1 May, 2015 and 1 October, 2016 (Fig. 7-F).
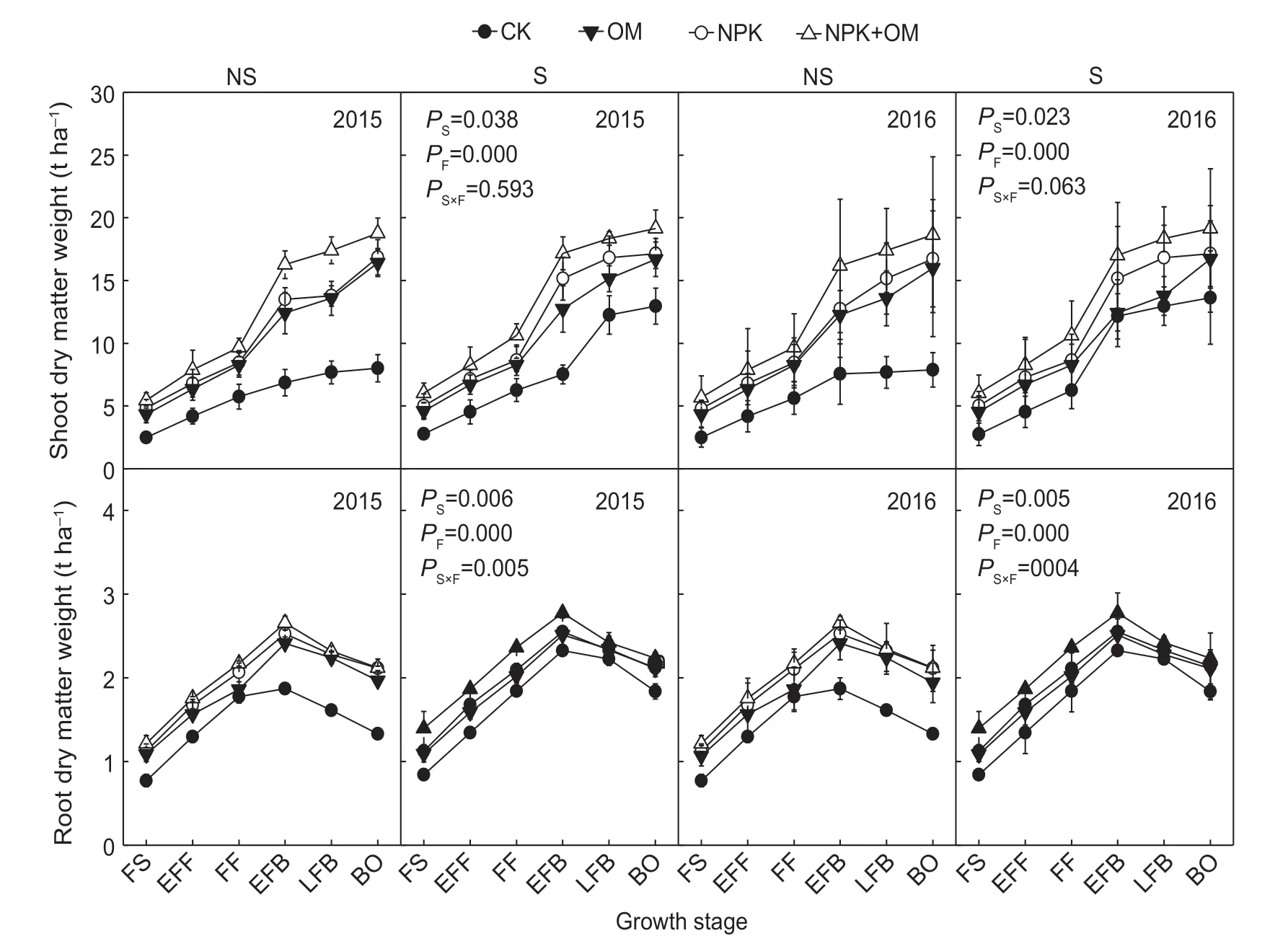
Fig. 5 Cotton biomass production as inf luenced by residue management and fertilizer application. NS, no residue incorporation; S, residue incorporation; CK, no fertilizer; OM, manure; NPK, N, P, and K fertilizer; NPK+OM, N, P, and K fertilizer+manure. FS, full square stage; EFF, early full f lower stage; FF, full f lower stage; EFB, early full boll stage; LFB, late full boll stage; BO, boll open stage. P S, P F and P S×F stood for the P-value of variance analysis for soil organic carbon storage under the effects of straw, fertilization and their interaction, respectively. Values represent mean±SD (n=15).
4. Discussion
4.1. Bulk density
Soil bulk density is soil basic property and signif icantly inf luenced by tillage and irrigation. Obvious seasonal variations in bulk density was observed and increased between sowing and harvest in 2015 and 2016 (Table 4). Based on plough and rotary tillage, soil bulk density was reduced (Abdalla et al. 2015). Therefore, this might be related to soil sinks driven by gravity and decreased in soil porosity caused by the increased irrigation during cotton growing seasons as a result of soil compaction (Gong et al. 2015). As found in India (Lemke et al. 2010) and Ireland (Groenigen et al. 2011), residue incorporation and fertilization signif icantly reduced soil bulk density in a longterm experiment. This is not surprising since bulk density in NPK+OM has a decreased trend in contrast to previous sampling in 2009 (Tables 1 and 4), and this trend can be attributed to the denser superf icial root network (Kou et al. 2012) and macroaggregate content was increased with large amounts of organic matter input (Zhang et al. 2014). In contrast, bulk density in N+CK has been signif icantly increased. There were no signif icant effects of residue management and fertilization on bulk density under other treatments. The reason is that the six-year experiment is short for studying the response of bulk density to residue management and fertilization. Similar result was also found by Groenigen et al. (2011).
4.2. SOC pool
SOC pool is inf luenced by agricultural management, tillage, water regime, soil type and season fractions (i.e., plant growth, temperature and precipitation) during the growing seasons (Lal 2009; Abdalla et al. 2015; Zhu et al. 2015). In the present study, seasonal changes in SOC pool were much more pronounced than the treatment effects, and there was negative linear correlation between seasonal changes in SOC and bulk density (R2=0.1159, P=0.0179) (data no shown). But solar radiation, temperature, precipitation and irrigation (Fig. 1 and Table 3) were the best at full f lower stage as a result of cotton grown stronger (Fig. 4), which signif icantly increased SOC content, especially in arid region with poor soil and signif icant climate change (Xu et al. 2014).
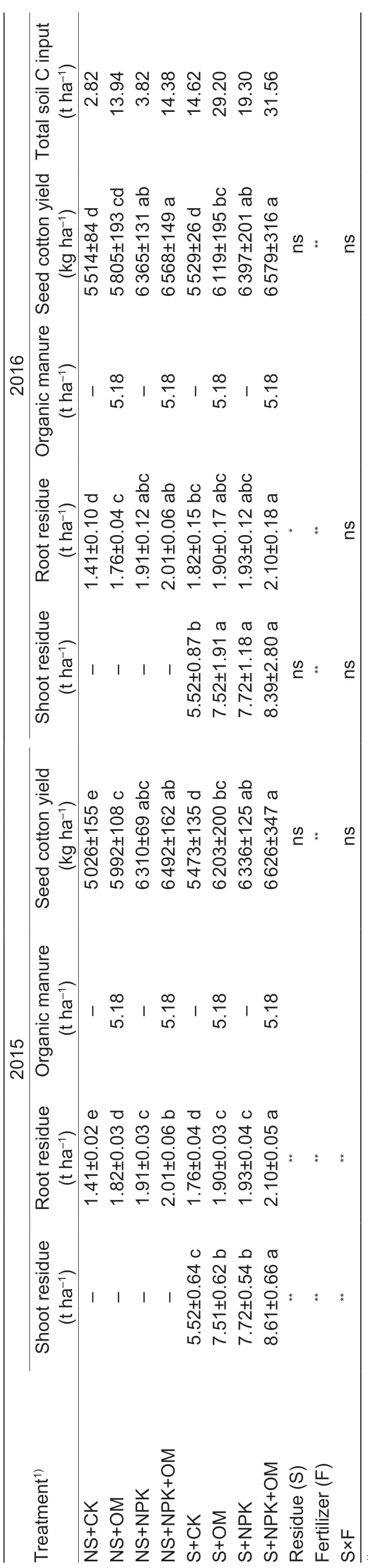
Table 6 Estimated C content of shoot residue, root residue, organic manure and soil C input, and seed cotton yield as affected by residue management and fertilizer treatments
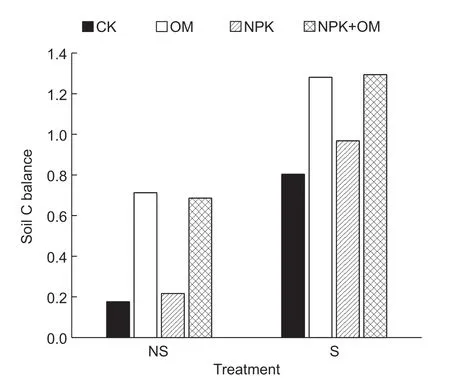
Fig. 6 The effect of residue management and fertilizer application on the soil C balance from May 2015 to October 2016. NS, no residue incorporation; S, residue incorporation. CK, no fertilizer; OM, manure; NPK, N, P, and K fertilizer; NPK+OM, N, P, and K fertilizer+manure.
The SOC mineralization rates are generally stable under long-term land use patterns (Kou et al. 2012). In the present study, SOC increased with increasing soil C input (R2=0.559, P=0.0330, in 0-20 cm; R2=0.522, P=0.0430, in 20-40 cm) (Fig. 7-A). This indicated that changes in SOC were directly related to the amount of C input from crop residues and organic manure. Similar results were reported in a 20-year f ield experiment (Qiao et al. 2015) and a 50-year f ield experiment (Lemke et al. 2010).
Great emphasis has been placed in the past on the importance of residue incorporation in Xinjiang. In this study, SOC and its labile fractions (i.e., LBC, MBC, WSOC and HWSOC) increased signif icantly in the residue-incorporated treatments but decreased in the no residue treatments (Table 5). This may be because soil enzyme activity was enhanced by soil moisture and temperature conditions in the plastic f ilm mulch drip-irrigation system, thus increasing organic matter decomposition (Zhang et al. 2014, 2016). Residue incorporation not only increases soil C input, but also improves soil aggregation and aggregate stability which protect organic matter from decomposition and reduce soil degeneration (Zhang et al. 2014). These f indings, which agree with results of several previous experiments, indicate that long-term residue incorporation is an effective practice for increasing SOC in arid areas (Feng et al. 2011; Groenigen et al. 2011).

Fig. 7 Relationships among soil C input, soil organic carbon (SOC), canopy apparent photosynthetic rate (CAP), cumulative CO2 emission, and soil C balance from May 2015 to October 2016. Values are means of the average of two growing seasons (n=8).
The SOC and its labile fractions were signif icantly greater in NPK+OM than in NPK (P<0.05) (Table 5). One reason is the readily metabolizable C and N in organic manure increased SOC by promoting plant growth, especially root growth and the production of root exudates (Bhattacharyya et al. 2011). Besides, the C/N ratio in NPK+OM is appropriate for organic material decomposing (Zhang et al. 2012), but the C/N in NPK is low as result of decreased the activity of microorganisms (Xu et al. 2014). In our study, NPK signif icantly increased SOC. This f inding contradicted Zhang et al. (2012), who reported that NPK had no effect on SOC in a paddy soil in Jiangxi Province, China. The NPK increased crop production and the return of crop residues and root-related C in both locations. However, SOC concentrations are relatively high in Jiangxi, and the effect of additional C inputs is small. In contrast, SOC concentrations in Xinjiang are relatively low, and additional C input causes SOC to increase. Soil C loss is also reduced by low temperatures during winter in Xinjiang (Fig. 3). Bhattacharyya et al. (2011) and Zheng et al. (2015) observed that climate signif icantly affects the geographic distribution of SOC. The change of organic C fractions were faster than SOC (Table 5) during the growing seasons, indicated that labile organic C fractions were more sensitive to changes in soil quality. This agrees with many studies which indicate that labile SOC fractions are early indicators of changes in soil quality (Blair et al. 1995; Mandal et al. 2013; Zhu et al. 2015).
4.3. Canopy apparent photosynthesis
Soil impoverishment is a serious problem in Xinjiang (Zhang et al. 2014). Soil nutrient supply is the main factor affecting CAP. Therefore, CAP is affected by residue management and fertilizer application (Egodawatta et al. 2012; Saikia et al. 2015). In the present study, CAP was generally greater in residue-incorporated treatments than in the no residue treatments (Fig. 4). For the fertilizer treatments, CAP generally decreased in the order NPK+OM>NPK>OM>CK. Furthermore, CAP increased as SOC increased (R2=0.726, P=0.0072, in 0-20 cm; R2=0.520, P=0.0436, in 20-40 cm) (Fig. 7-B). The most likely reason is that long-term residue incorporation and inorganic fertilizer with organic manure application increased SOC (Fig. 7-A), which improved soil quality by enhancing organic C fractions contents (Table 5) and soil nutrient turnover rate (Lal 2004, 2013b; Wu et al. 2015; Zhu et al. 2015). In addition, residue return and organic manure application signif icantly increase soil CO2emission (Fig. 3-C), which can alter CO2concentrations in the midcanopy and increase plant photosynthesis and net primary production (Gill et al. 2002). In turn, greater CAP enhances the production of above- and below-ground biomass and the release of root exudates, thus increasing organic C inputs to the soil (Bhattacharyya et al. 2011). Moreover, changes in CAP affect the accumulation of photosynthesis product and therefore yield (Gomez-Casanovas et al. 2012; Saikia et al. 2015). Residue incorporation and fertilizer application in this study increased shoot, root biomass and yield, probably due to higher CAP. Mandal et al. (2013) reported that soil which is inherently low in SOC and N can be highly productive if adequate amounts of fertilizer and manure are applied. These f indings suggest that a desirable nutrient management practice in arid areas (i.e., residue incorporation and inorganic fertilizer plus organic manure) would be the one that sustains reasonably high yield levels and simultaneously improves fertility, especially SOC storage.
4.4. Soil CO2 emission
A distinct seasonal variation in soil CO2f lux was observed. Soil CO2f lux was the highest in summer and the lowest in winter (Fig. 3-A). Soil respiration generally increases as soil temperature increases when water is not limiting (Ding et al. 2007; Gong et al. 2015). Farmers in Xinjiang generally use a combination of plastic f ilm mulch and drip irrigation, thus keeping the soil moist during the growing season. In this study, there was signif icant positive correlation between soil CO2f lux and temperature (R2=0.654, P<0.0001) between 1 May, 2015 and 1 October, 2016 (Fig. 3-B). Our results agree with Ding et al. (2007), who reported that (i) temperature was related to soil CO2f lux and (ii) temperature accounted for 49-66% of the seasonal variation in CO2f lux. Our results contradicted Dossouyovo et al. (2016), who reported that temperature had no signif icant effect on soil CO2f lux in northern Benin. Those authors concluded that soil CO2f lux at their experiment site was mainly dependent on soil moisture. There was a sudden increase in soil CO2f lux on 15 November, 2015. This was caused by deep tillage which had occurred the previous week. Tillage can result in an immediate shortterm outburst of CO2due to a decrease in CO2partial pressure and disturbance of soil aggregates and pores (Lal 2013b; Abdalla et al. 2015; Dossouyovo et al. 2016). The results of this study suggested that less tillage could reduce soil CO2emission and increase the SOC pool.
Residue and fertilizer are major factors affecting soil respiration in farmland, directly by inf luencing root and microbial activities and indirectly by inf luencing soil physical and chemical properties, plant growth, and the substrate for microbes (Ding et al. 2007; Huang et al. 2012; Han et al. 2014; Fan et al. 2015; Gong et al. 2015; Mancinelli et al. 2015). In our study, the average and cumulative CO2emissions were signif icantly greater by 14.97-15.45% in the residue-incorporated treatments than in the no residue treatments (Fig. 3-D). This agrees with Abdalla et al. (2015), who also found that the incorporation of crop residue increased soil CO2emission by 18%. Hu et al. (2014) reported that wheat straw incorporation in Edmonton, Canada increased soil CO2emission by 40%. This increase was much larger than the one we observed. One reason may be that Edmonton has a humid continental climate. Temperatures and soil moisture conditions are more favorable for microbial activity in Edmonton than in Xinjiang. Bhattacharyya et al. (2011) and Groenigen et al. (2011) reported that differences in climatic conditions can explain variations in the decomposition rate of crop residue and soil CO2emission. The average and cumulative CO2emissions in NPK were signif icantly greater than those in CK, but less than those in OM and NPK+OM (P<0.05) (Fig. 3-C). This was because C inputs from organic manure increased the SOC turnover rate and microbial activity in OM and NPK+OM (Iovieno et al. 2009; Qiao et al. 2015). Besides, the C source in NPK was limited as result of less soil CO2emission (Jiang et al. 2014).
There was strong positive correlation between cumulative CO2emission and SOC (R2=0.855, P=0.0010, in 0-20 cm; R2=0.806, P=0.0025, in 20-40 cm) and SOC storage (R2=0.977, P<0.0001, in 0-20 cm; R2=0.966, P<0.0001, in 20-40 cm) (Fig. 7-C). This indicated that the effects of residue management and fertilizer application on SOC and bulk density inf luenced cumulative soil CO2emission. Gomez-Casanovas et al. (2012) found that autotrophic and heterotrophic components of soil respiration were correlated with plant photosynthesis and, to a lesser extent, with soil temperature and moisture. We also found that cumulative CO2emission was signif icantly related to CAP (R2=0.575, P=0.0292) (Fig. 7-E). A similar relationship between photosynthesis and soil CO2emission was previously reported in maize and wheat f ields (R2=0.79) (Huang et al. 2012). This suggests that photosynthetic rates could be used to estimate seasonal soil CO2emission. Correlation analysis revealed that SOC and CAP during the cotton season respectively explained 92 and 58% of the difference in cumulative CO2emission among treatments from May 2015 to October 2016. These analyses indicate that SOC was the primary factor inf luencing soil CO2emission. Similar to our results, Ding et al. (2007) reported that SOC and CAP accounted for 96 and 78% of the difference in cumulative CO2emission among treatments. In comparison, CAP and soil temperature together accounted for 53% of the variation in soil CO2emission in a coastal wetland (Han et al. 2014). One reason is that the amount of vegetation in coastal wetland is much less than in cotton f ields (2.05×105plants ha-1).
4.5. Soil C balance
Soil C balance in farmland depends mainly on the balance between soil C input and soil CO2emission (Kou et al. 2012). Residue incorporation and fertilizer application increased both soil C input and cumulative soil CO2emission (Table 6 and Fig. 3-D). The S+OM and S+NPK+OM treatments increased C input more than cumulative soil CO2emission (Fig. 6). These results indicated that residue incorporation and organic manure application are prerequisite measures for increasing soil C sequestration and maintaining soil quality in arid parts of the Xinjiang Region. We estimated that S+NPK (a common agricultural practice in Xinjiang) resulted in a loss of -0.32 t C ha-1yr-1. This meant that a minimum of 0.32 t C ha-1yr-1(approximately 1.9 t ha-1yr-1poultry manure) must be added to the soil to offset C losses and maintain the soil C pool. Linear analysis showed that soil C balance was signif icantly correlated with soil C input (R2=0.975, P<0.0001) (Fig. 7-F). This suggests that the soil in this study was not C saturated. According to the conceptual soil C sequestration model proposed by Lal (2004), the soil in this study has a substantial saturation def icit and therefore has potential to sequester C.
5. Conclusion
The SOC pool responded strongly to residue management and fertilizer application in this arid region. The current residue management and fertilizer practices (i.e., S+NPK) are suff icient for plant photosynthesis and yield development. However, they cannot enhance SOC storage and soil quality. The combination of residue incorporation and inorganic fertilizer plus organic manure application signif icantly enhanced SOC and its labile fractions contents, thus increasing the ability of the soil to supply nutrients for crop growth. Moreover, soil CO2emission was signif icantly correlated with SOC. This meant that organic amendment simultaneously increased SOC, thereby enhancing soil CO2emission. The calculation of soil C balance demonstrated that the soils in Xinjiang are a source of CO2, except when residue is incorporated and organic manure is applied. If the residue is incorporated and both inorganic fertilizer and organic manure are applied, then the C sequestration potential of this soil is 3.56 t C ha-1yr-1. Hence, the combination of residue incorporation and inorganic fertilizer plus organic manure application is a strategy to achieve food security and mitigate climate change by improving soil C sequestration and soil quality in arid regions.
Acknowledgements
This study was supported by the National Natural Science Foundation of China (31471450). The authors are also grateful to Dr. William J. Gale (Agricultural College, Shihezi University, China) for his helpful revision of the paper especially in English language. We would also like to thank the reviewers for helping us to improve our original manuscript.
 Journal of Integrative Agriculture2019年5期
Journal of Integrative Agriculture2019年5期
- Journal of Integrative Agriculture的其它文章
- Characterization of TaCOMT genes associated with stem lignin content in common wheat and development of a gene-specif ic marker
- Phenotypic characterization and genetic mapping of the dwarf mutant m34 in maize
- Morphological diversity and correlation analysis of phenotypes and quality traits of proso millet (Panicum miliaceum L.) core collections
- Field identif ication of morphological and physiological traits in two special mutants with strong tolerance and high sensitivity to drought stress in upland rice (Oryza sativa L.)
- Crosstalk of cold and gibberellin effects on bolting and f lowering in f lowering Chinese cabbage
- Foliar spraying of aqueous garlic bulb extract stimulates growth and antioxidant enzyme activity in eggplant (Solanum melongena L.)