
环境因子对浮游动物休眠卵萌发的影响
2019-05-10 07:42:38栾磊磊杨宇峰王庆高良敏
暨南大学学报(自然科学与医学版) 2019年2期
栾磊磊, 杨宇峰, 王庆, 高良敏
(1.安徽理工大学 能源与安全学院, 安徽 淮南 232000;2.暨南大学 水生生物研究所∥水体富营养化与赤潮防治广东省教育厅重点实验室, 广东 广州 510632)
休眠是海洋浮游动物度过不良环境的重要生存策略[1-2],水体中的现生种群和沉积物中的休眠种群组成了浮游动物种群.在水生态系统中,当食物短缺、盐度或季节变化等不利因素出现时,物种的种类和种群密度会发生变化,当生存环境好转时,又恢复到原来的水平,这是因为沉积物中存在浮游动物的“卵池”[3-4].河口浮游动物为适应独特的环境,以产休眠卵或迁移等方式度过不良环境[5-6],休眠卵在沉积物中累积形成卵池或卵库,卵库对水体中浮游种群结构、演替和维持种群稳定具有重要意义[7-9].
关于休眠卵分布的研究主要集中在北温带,亚热带的报道较少[2,10].珠江口是我国重要的亚热带河口,桂山岛位于珠江口咸淡水交汇区,环境波动较大.选取桂山岛海域作为研究区域,以探讨沉积物中休眠卵萌发受环境因子的影响,研究结果为亚热带河口浮游动物的潜在补充机制提供了基础数据,对揭示亚热带河口浮游动物群落演替规律具有重要参考价值.
1 材料与方法
1.1 样品的采集及质量控制
用彼得逊采泥器采集0~5 cm表层沉积物于 4 ℃冰箱中冷藏,用于休眠卵萌发实验;水温、盐度和pH等用美国YSI plus多参数水质分析仪现场测定;水体浮游动物采集使用40 μm的浮游生物网由底层至表层垂直拖网,并加甲醛固定,最终含甲醇质量分数为4%.
1.2 实验组设置
实验使用CuCl2、ZnCl2、CdCl2、NaCl、HCl、NaOH优级纯试剂,用去离子水配成母液后备用.实验对重金属和生源要素进行前处理及质量控制,方法同Gu等[11-12]一致,同步采用GBW07437国家标准对标准物质进行了内标和外标的回收率实验,应用于校正分析结果,样品中重金属和总磷(TP)均采用美国Optima 2100DV光谱仪测定,通过对结果的综合,沉积物中TP的回收率介于92%~97%,Cu、Zn和Cd的回收率介于89%~102%.取40 g沉积物置于烧杯中,根据采样点沉积物背景值的质量分数(测定沉积物中Cu、Zn、Cd的背景值分别为29.46、105.20、0.18 mg/kg[13])和沉积物的含水率(57.42%),采用Jiang等[14]方法,计算出样品去除含水率后的质量和Cu、Zn、Cd的含量,并结合样品去除含水率后的体积,据此调节CuCl2、ZnCl2、CdCl2的浓度.
将Cu、Zn、Cd重金属浓度调为背景值的2、4、8、16、32倍,将不同浓度的重金属对沉积物休眠卵分别处理48、72和92 h;取250 g沉积物置于烧杯中,用单因子变量法进行实验,分别将温度调为15、20、25、30、35 ℃,盐度调为0、10、15、20、25、30、35、40、50,pH调为6、7、8、9、10,沉积物中休眠卵分别冷藏半个月和10个月后进行萌发实验.实验用海水均采用 0.45 μm 滤膜进行过滤,以排除海水中浮游动物的干扰,每组设3个平行,并设置对照组,将沉积物中休眠卵置于GXZ 300c型智能照明培养箱,温度(25±1)℃,盐度30,光照L ∶D=12 ∶12) 中培养萌发.
此外还在原位海区不同水深进行休眠卵的萌发实验,将沉积物置于透明瓶中,加入原位海水(0.45 μm 滤膜过滤),用双层筛绢网将口封住,分别置于离海面深度0.5、1、2 m,每个深度3个重复试验.
1.3 样品萌发与计数
将样品用20 μm 筛绢网过滤,并将截留样品置于奥林巴斯体视显微镜SZX7下镜检萌发的幼体并计数;更换0.45 μm 滤膜过滤后的新鲜海水于实验组中继续萌发,若连续8 d 以上没有观察到幼体孵化时,即判定沉积物中存活休眠卵已全部孵化.
1.4 数据处理
使用 SPSS 17.0 进行数据分析,Origin 8.5进行绘图.
2 结果与分析
2.1 调查海域基本情况
调查海域水温29.4 ℃,盐度22.68,pH 8.37,溶解氧5.59 mg/L;沉积物中TP质量分数为403.96 mg/kg,Cu、Zn、Cd的质量分数分别为29.46、105.20、0.18 mg/kg[13].采样期间水体中共发现小拟哲水蚤、刺尾纺锤水蚤、锥形宽水蚤、太平洋纺锤水蚤、亚强次真哲水蚤、微刺哲水蚤、左指华哲水蚤7种,其中已报道产休眠卵的种类有刺尾纺锤水蚤、太平洋纺锤水蚤、左指华哲水蚤[15].
2.2 重金属处理对休眠卵萌发的影响
Cu处理48 h组和72 h组增加到背景值的16倍,96h组增加到背景值2倍,沉积物中休眠卵死亡率达到100%;Zn处理48 h组增加到背景值的32倍,72 h组增加到背景值的16倍,96 h组增加到背景值的8倍,沉积物中休眠卵死亡率达到100%;Cd处理48、72、96 h组增加到背景值的32倍,沉积物中休眠卵死亡率都未达到100%;混合处理48 h组增加到背景值的16倍,72 h组增加到背景值8倍,96 h组增加到背景值的2倍,沉积物中休眠卵死亡率达到100%.因此,增加浓度、增加处理时间休眠卵孵化的幼体数量明显减少,休眠卵死亡率呈明显上升趋势(图1).
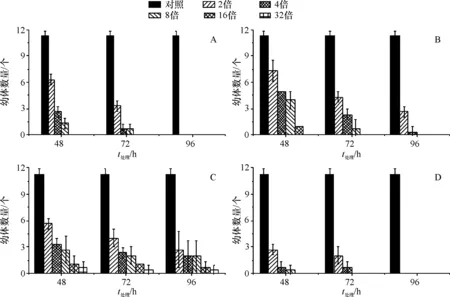
A:铜处理休眠卵的孵化量;B:锌处理休眠卵的孵化量;C:镉处理休眠卵的孵化量;D:铜、锌、镉混合处理休眠卵的孵化量
A:Amount of incubation of resting eggs treated with copper; B:Amount of incubation of resting eggs treated with zinc; C:Amount of incubation of resting eggs treated with cadmium; D:Incubation amount of resting eggs treated with copper, zinc and cadmium
图1 不同重金属处理休眠卵的孵化量
Fig.1 Amount of incubation of resting eggs treated with different heavy metals
2.3 水温对休眠卵萌发的影响
实验水温分别调为15、20、25、30、35 ℃,研究水温对沉积物中休眠卵萌发的影响.结果表明在15 ℃~25 ℃区间,随着水温升高休眠卵孵化数量增多,水温超过25 ℃孵化数量减少.沉积物中休眠卵萌发的最适水温是25 ℃,此温度下萌发的数量最多(78个),水温达到35 ℃时仅萌发2个(图2),不同水温对休眠卵萌发的影响有统计学差异(P<0.05).
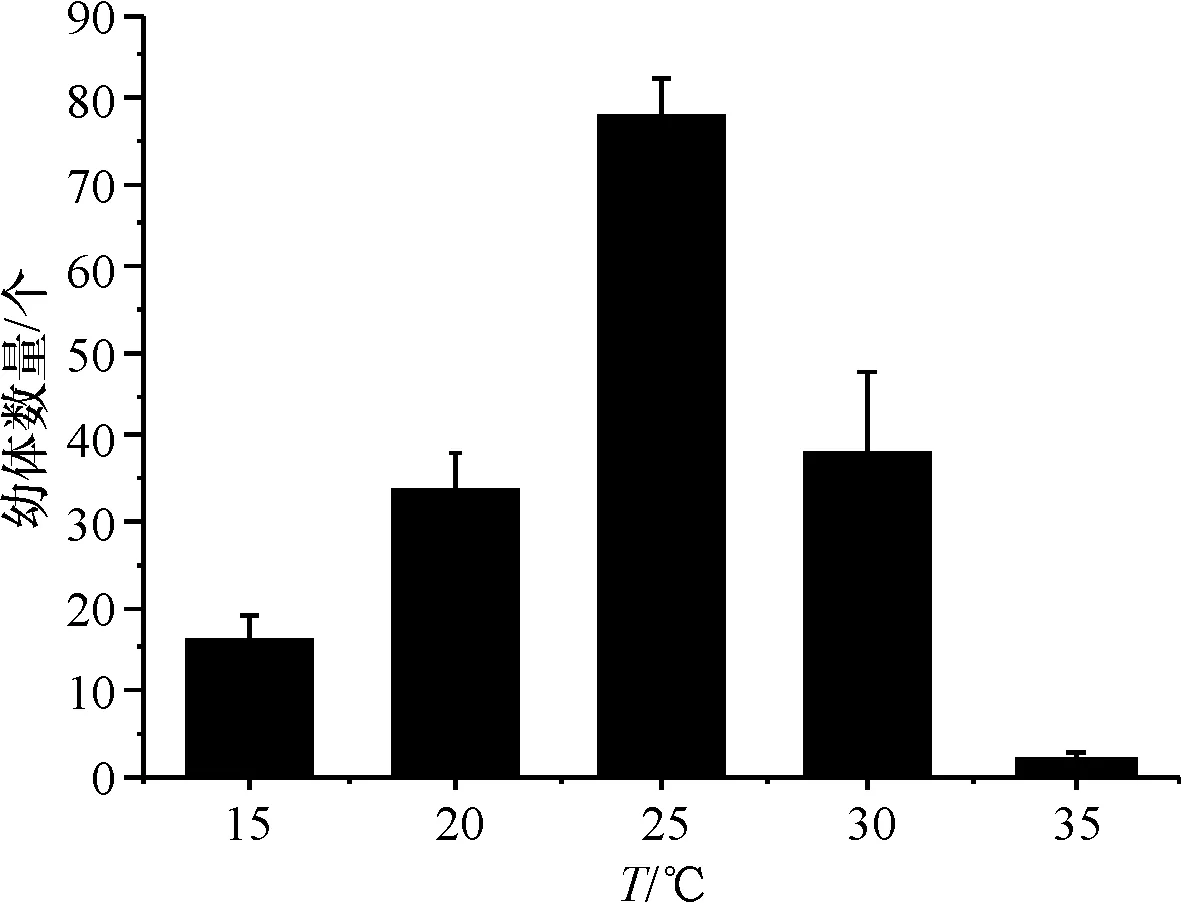
图2 不同水温休眠卵的孵化量
Fig.2 Amount of incubation of resting eggs in different water temperature
2.4 盐度对休眠卵萌发的影响
实验盐度分别调为0、10、15、20、25、30、35、40、50,研究盐度对沉积物中休眠卵萌发的影响,在盐度为0~30区间随着盐度增加,休眠卵萌发的数量增多,盐度超过30萌发数量减少,萌发的最适盐度为30,此盐度下萌发的数量最多(82个),盐度过高或过低都不利于休眠卵萌发(图3),不同盐度对休眠卵萌发的影响有统计学差异(P<0.05).
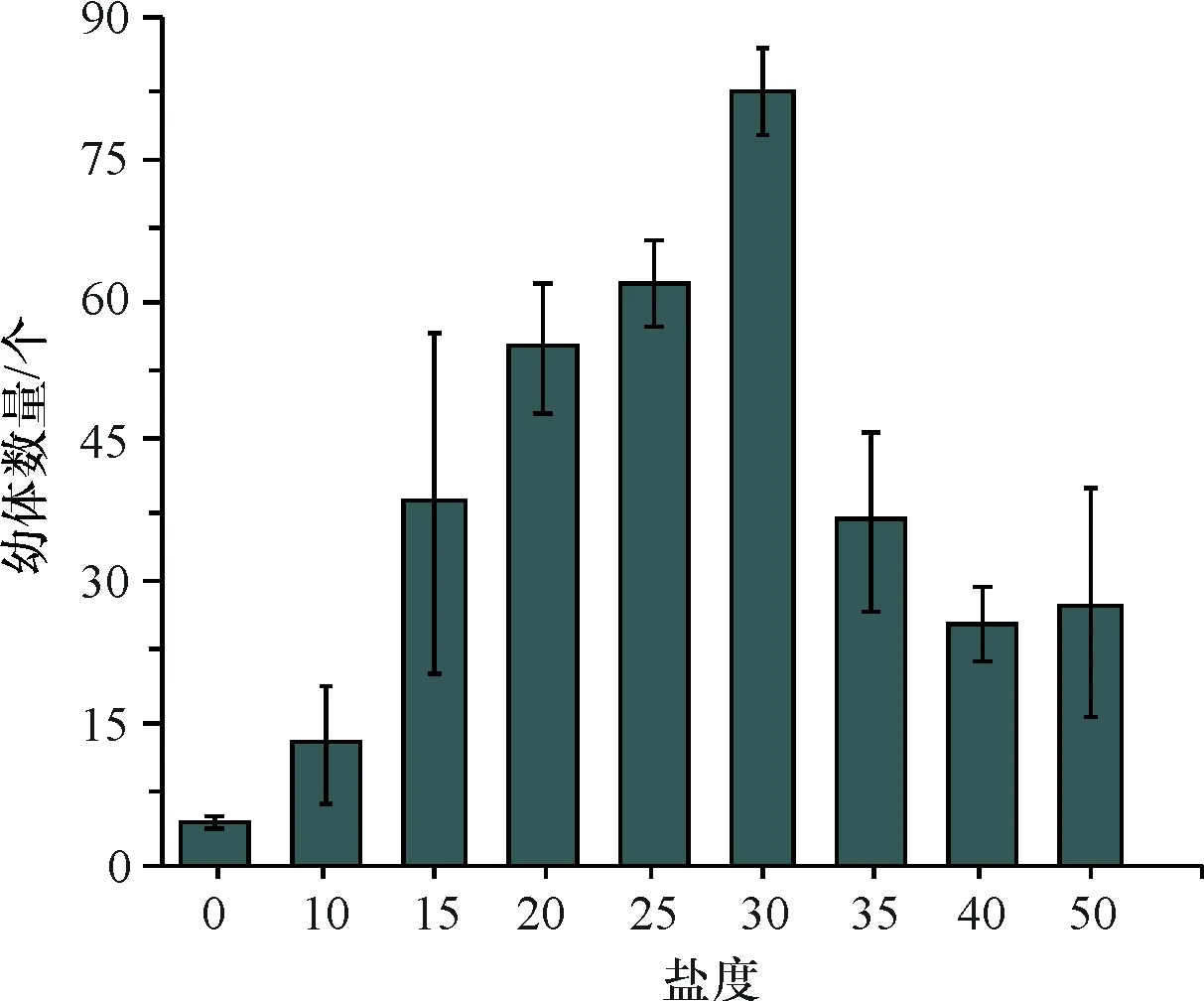
图3 不同盐度休眠卵的孵化量
Fig.3 Amount of incubation of resting eggs in different salinity
2.5 pH对休眠卵萌发的影响
实验pH分别调为6、7、8、9、10,研究pH对沉积物中休眠卵萌发的影响,结果表明pH在6~8区间随着pH增加,休眠卵萌发的数量增多,pH 为8是萌发的最适 pH,在此条件下休眠卵萌发的数量最多(83个),海水pH过高(pH<8)或过低(pH>8)都不利于休眠卵萌发(图4),pH不同的海水对休眠卵萌发的影响有统计学差异(P<0.05).
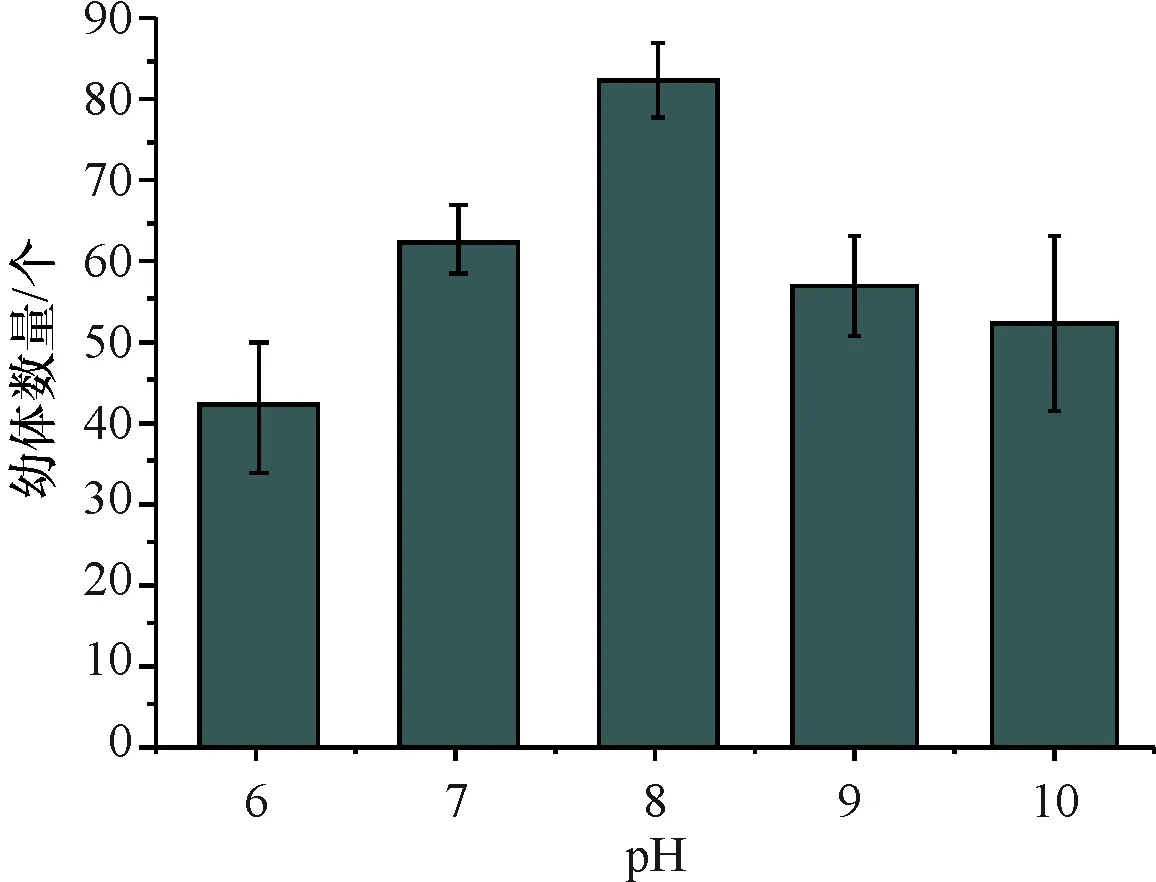
图4 不同pH休眠卵的孵化量
Fig.4 Amount of incubation of resting eggs under different pH
2.6 低温冷藏对沉积物休眠卵萌发的影响
于8月、9月和10月3次采集沉积物样品,将沉积物分别冷藏半个月和10个月后进行实验,发现桡足类休眠卵冷藏10个月后潜在补充量(8月为8.4×105ind./m3,9月为6×105ind./m3,10月为5.3×105ind./m3)显著高于冷藏半个月(8月为0.18×105ind./m3,9月为0.47×105ind./m3,10月为0.56×105ind./m3),轮虫休眠卵冷藏半个月后休眠卵潜在补充量(8月为0.007×105ind./m3,9月为0 ind./m3,10月为0.027×105ind./m3)明显少于桡足类,而冷藏10个月后轮虫休眠卵潜在补充量为0(图5),表明桡足类和轮虫休眠卵对冷藏刺激的响应不一.
2.7 原位海区不同水深对休眠卵萌发的影响
休眠卵放置在采样海域进行孵化,结果显示:放置在离海面0.5 m处休眠卵潜在补充量(4.44×104ind./m3)显著高于2 m处(1.22×104ind./m3).沉积物中休眠卵的孵化过程大致呈锯齿状分布,第3天潜在补充量最多(0.5、1和2 m处分别为:0.089×104、0.056×104、0.022×104ind./m3),孵化持续50天左右,不同水深之间孵化持续时间无明显差异(图6).
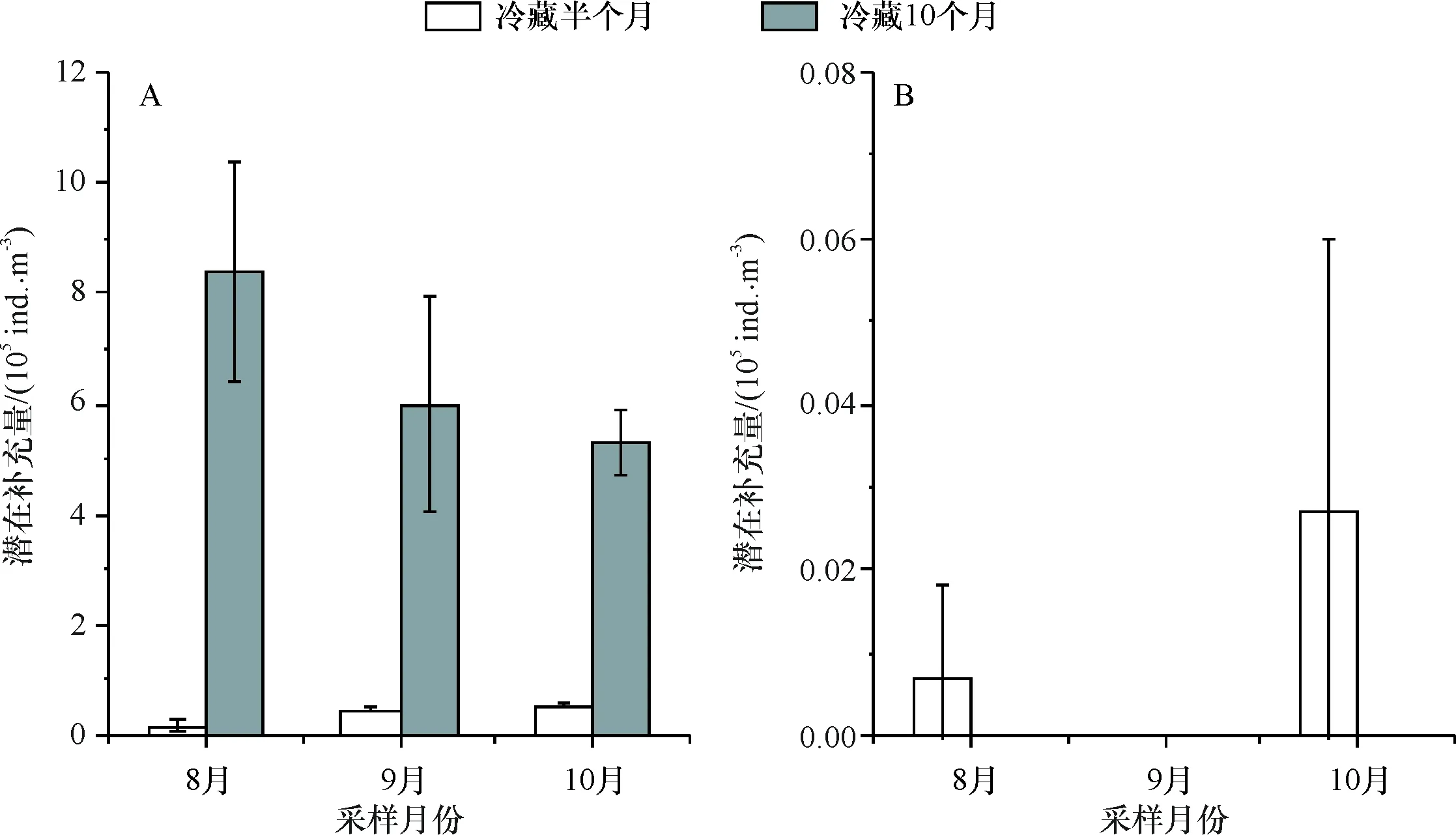
A:不同冷藏时间桡足类休眠卵的潜在补充量;B:不同冷藏时间轮虫休眠卵的潜在补充量
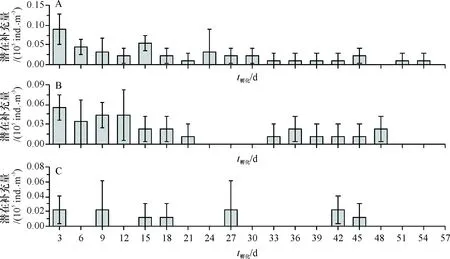
A:0.5 m处休眠卵的潜在补充量;B:1 m处休眠卵的潜在补充量;C:2 m处休眠卵的潜在补充量
A:Potential supplement of resting eggs at 0.5 meter water depth; B:Potential supplement of resting eggs at 1 meter water depth; C:Potential supplement of resting eggs at 2 meter water depth
图6 原位海区不同水深休眠卵的潜在补充量
Fig.6 Potential supplement of resting eggs under different depths of situ sea area
3 讨论
3.1 沉积物中休眠卵对重金属的响应
桂山岛沉积物中Cu潜在生态危害较重,属于严重污染,Zn潜在生态危害较轻,属于中污染、Cd潜在生态危害较轻,属于低污染[13].本研究选取Cu、Zn、Cd 3种重金属对沉积物中休眠卵的毒性影响,结果显示毒性效应由大到小依次为Cd>Cu> Zn.沉积物中休眠卵对Cd非常敏感,在暴露2倍于Cd背景值即可使其死亡率达到50%.
Natalia等[16]研究发现Cu、Cd、Zn对多刺裸腹蚤(Moinamacrocopa)休眠卵的半致死浓度(LC50)(24 h)分别为(0.009 4±0.000 2)、(0.284±0.029),(1.19±0.18)mg/L,LC50 (48 h)分别为(0.009 4±0.000 2)、(0.169±0.020)、(0.34±0.02)mg/L,对Cd、Zn较敏感;Nandini等[17]研究发现Cu、Zn对Moinamacrocopa休眠卵的LC50 (24 h)分别为0.71、1.01 mg/L,徐善良等[18]研究发现Cd对Moinamacrocopa休眠卵的LC50 (48 h)为0.053 mg/L,结果表明同一物种休眠卵在不同区域,环境不同,对重金属的耐受性表现不一致,半致死浓度的差异较大.
厦门湾沉积物中Cu、Cd两种重金属对太平洋纺锤水蚤(Acartiapacifica)休眠卵的孵化影响结果表明,Cu质量分数从34.8 mg/kg增加到348 mg/kg,休眠卵孵化量减少46.6%~100%,Cd质量分数从0.68 mg/kg增加到6.8 mg/kg,休眠卵孵化量减少31.6%~94.7%[14],实验重金属浓度和死亡率均高于桂山岛,可能因为采样区域不同,采样点的污染程度不同,沉积物中休眠卵对重金属的耐受性不一样,且试验物种也不一样,对休眠卵孵化影响差异较大.
此外,重金属对对河口底栖桡足类(Amphiascustenuiremis)成体的毒性影响由大到小依次为Cd>Cu>Ni>Zn>Pb[19],与本研究对休眠卵的毒性影响结果一致,重金属对底栖浮游动物及其休眠卵均有影响.
3.2 沉积物中休眠卵对水温、盐度、pH的响应
水温、光照及 pH 等环境因素对沉积物中休眠卵具有重要影响[2,10,20-22].不同种类浮游动物适应不同的水温、盐度和pH等条件,研究发现,日本海一种浮游动物Tortanusforcipatus休眠卵可在13 ℃~30 ℃温度范围孵化,25 ℃孵化率最高,在盐度为18~54时均有幼体孵化[23];波罗的海的一种纺锤水蚤(Acartiatonsa)休眠卵在温度为5 ℃~23 ℃,盐度为 0~34水体中都可以成功孵化,水温23 ℃时孵化率最高为92.2%,盐度在17~25之间时孵化率最高为81.4%~84.5%[24].
本研究中,沉积物中的休眠卵能在水温为15 ℃~35 ℃,盐度为0~50,pH 为6~10的条件下萌发,表明沉积物中有适应不同水温、盐度和pH种类的休眠卵,能在相应条件下孵化以补充水体中的浮游动物种群,从而在变化的环境中维持种群的延续.有研究表明,浮游动物群落在盐度变化的刺激后,其恢复是由于现存卵库的作用[25-26].
3.3 沉积物中休眠卵对低温冷藏的响应
休眠卵必须度过内滞育期才能孵化,低温刺激有助于缩短其内滞育期并保持可育性,因此低温冷藏对休眠卵萌发具有重要影响[22,27-28].Glippa等[5]在研究休眠卵孵化时,发现冷藏11个月后(孵化率4.5%) 比立即进行孵化(孵化率0.7%)孵化率明显提高,厦门湾中心区域Acartiapacifica休眠卵在10 ℃储存3~60 d用于研究碳氢化物污染对沉积物休眠卵的影响[29],Kasahara等[30]发现5 ℃将休眠卵冷藏14~15个月孵化率明显低于冷藏1~2个月,Vandekerkhove等[31]将休眠卵在黑暗环境下4 ℃冷藏7个月后进行休眠卵分离孵化实验.
本研究结果表明桡足类和轮虫休眠卵在4 ℃冷藏半个月和10个月,孵化量差异显著,桡足类休眠卵和轮虫休眠卵对低温刺激的响应不同,可能是因为桡足类休眠卵不应期较长,需经过较长时间的低温刺激,而轮虫休眠卵内滞育期较短或已渡过内滞育期.研究发现桡足类哲水蚤(Acartiabilobata)在4 ℃冷藏14天孵化率最高,冷藏1年后孵化率下降[32].不同海域沉积物中休眠卵的种类不同,环境不同,导致其内滞育期不同,对低温刺激的响应不同,需要低温刺激的时间不同.
3.4 休眠卵萌发对原位环境的响应
本研究的原位萌发是直接将沉积物置于原位海区中,利用自然海区的水温、光照、海水搅动等,模拟休眠卵在采样区域水体中的萌发.测得深度为0.5、1、2 m处白天的光照强度分别为193~266、108~230、47~70 μmol·m-2·s-1,离海平面越远光照强度越弱,休眠卵孵化量也越少,休眠卵在不同水深孵化数量与光照呈显著正相关(P<0.01),2 m深度处休眠卵孵化数量只有0.5 m深度处的1/4.
4 结论
Cu、Zn和Cd均对沉积物中休眠卵有较大影响,可导致沉积物中休眠卵死亡率增加,Cd对休眠卵的毒性效应最大,且随着三种重金属浓度的增加和处理时间的延长,休眠卵死亡率呈明显上升趋势;休眠卵能在较宽的水温、盐度和pH范围萌发,且随着水温、盐度和pH的增加孵化数量呈上升趋势,到达最适值后呈下降趋势,不同水温、盐度和pH对休眠卵萌发的影响有统计学差异(P<0.05),桂山岛海域沉积物中休眠卵最适孵化条件为:水温 25℃、盐度 30、pH 8;冷藏刺激有利于沉积物中桡足类休眠卵的萌发,桡足类休眠卵冷藏10个月后萌发量显著高于冷藏半个月;放置在采样海域进行孵化,休眠卵的孵化数量受光照影响较大,随着光照强度减弱,孵化数量显著减少,孵化数量与光照强度呈显著正相关(P<0.01).
猜你喜欢
中国蚕业(2022年1期)2022-03-09 02:59:20
今日农业(2021年6期)2021-11-27 08:05:59
生物学通报(2021年9期)2021-07-01 03:24:44
辽宁化工(2021年5期)2021-06-03 05:15:50
——以“不同咖啡因饮料对水蚤心率的影响”为例
中学生物学(2021年2期)2021-04-30 12:04:06
蚕桑通报(2020年3期)2021-01-07 03:05:20
河北渔业(2019年1期)2019-02-13 01:35:34
河南科技(2015年21期)2015-03-26 21:21:21
湖南农业(2015年7期)2015-03-20 01:48:37
海洋与湖沼(2014年4期)2014-03-09 06:57:32
