
桉树生物量估算模型及与IPCC法的对比分析
2019-04-29 07:12揭凡杜阿朋竹万宽
桉树科技 2019年1期
揭凡,杜阿朋,竹万宽
桉树生物量估算模型及与IPCC法的对比分析
揭凡1,杜阿朋2,竹万宽2*
(1.中林集团雷州林业局有限公司石岭林场分公司,广东 廉江 524456; 2.国家林业和草原局桉树研究开发中心,广东 湛江 524022)
为构建适用于区域桉树人工林生物量异速生长方程,收集整理桉树林的生物量文献数据,拟合测树因子(胸径和树高)与地上、地下和单株生物量间的回归关系。结果表明:单自变量模型中,基于胸径因子的方程拟合优度高于树高因子。双自变量模型中,树高因子的添加仅对单株生物量拟合优度提高了0.7% ~ 1.5%。模型的预测值与实测值的比较及相容性分析表明,方程ln=-2.833+2.301ln+0.352 1ln对地上生物量预测效果最优,精度达94.6%;方程ln=-5.175+0.939lnDH对地下生物量预测效果最优,精度达66.8%;方程ln=-2.960+0.896lnDH对单株生物量预估效果最优,精度达95.5%。分量模型与单株模型相容性较好。桉树人工林和的平均值分别为0.634 1(n=65,SD=0.132)和0.205 6(n=76,SD=0.089)。IPCC法对林分生物量估算精度高于异速生长方程,达到95.3%。因此,建议采用IPCC生物量估算参数法进行区域尺度桉树人工林生物量估算。
桉树人工林;生物量;异速生长方程;区域尺度
森林是陆地生态系统的主体,作为陆地生态系统中最大的碳库[1],其在区域及全球碳循环中具有重要作用[2]。基于全球气候变化的背景,准确估算森林生物量对于了解森林在全球碳循环中的角色定位具有重要意义[3]。森林生物量占陆地植被总生物量的85% ~ 90%[2, 4],其总初级生产力约占全球陆地生态系统的一半[5]。森林生物量是研究森林生态系统生产力的基础,也是度量植被碳库、研究整个生态系统结构、功能、能量流动和物质循环的重要参数[6]。目前,森林生物量可通过直接测量和间接估算2种方法获得[7]:前者为收获法,破坏性大,不适用于乔木;后者是利用生物量模型、生物量估算参数和3S技术等方法进行估算[8]。生物量模型中的异速生长方程法通过构建相对易测的植株结构和功能特征指标(如生物量等)与易测的植株形态学变量(如胸径、树高等)的数量关系模型[9-10],从而估测林木或林分的生物量。因其精确度较高,而且能有效降低对森林和植被的破坏性取样[11],因而在森林生态系统生物量和生产力估算中是较为广泛使用的方法。生物量估算参数法是利用立木蓄积量,通过生物量估算参数估算区域尺度森林生物量的方法[12],常用的参数包括生物量转化与扩展系数(Biomass Conversion and Expansion Factor)、生物量扩展系数(Biomass and Expansion Factor)和根茎比(Root:shoot Ratio)等。
目前,国内外已有不少学者汇编了不同尺度下各树种的异速生长方程,如陈传国[13]、冯宗炜[14]、胥辉[15]、罗云建等[16]及WANG等[17]综合了中国主要树种的异速生长方程,JENKINS等[18]和TER-MIKAELIAN等[19]对美国主要树种的异速生长方程进行了归纳,ZIANIS等[20]汇编了美国和澳大利亚等地区主要树种的异速生长方程。但查阅汇编的方程较为繁琐,且只适用于某一地点和特定林分进行生物量估算,并不能应用到大尺度范围上森林生物量的估算。MONTAGU等[21]和WILLIAMS等[22]证实了可以构建不同种和不同区域的桉树生物量异速生长方程用于生物量估算,但也仅针对澳大利亚桉树种植区。政府间气候变化委员会(IPCC,Intergovernmental Panel on Climate Change)发布的《土地利用、土地利用变化和林业的优良做法指南》和《2006年IPCC国家温室气体清单编制指南》建议采用生物量估算参数法(IPCC法)用于区域尺度森林生物量估算[23-24]。
桉树()是我国南方三大速生树种之一,其因适应性广、生产力高等优势被大面积引种栽培,已成为重要的造林树种和木制纸原料之一。截至2015年,桉树人工林面积占世界人工林面积的15%,在木材供应方面占有重要地位[25]。目前,中国桉树人工林面积约为4.5 × 106hm2,年木材产量占全国的26.9%,不仅具有可观的经济价值,同时也是我国森林碳汇的重要组成部分。因此,本研究利用已公开出版或发表的桉树人工林生物量相关数据,以种植面积较广的尾巨桉()和巨尾桉()为研究对象,通过汇总、筛选和整合分析,探索桉树人工林生物量估算方法,以期为区域尺度森林生物量估算提供理论基础。
1 研究方法
1.1 文献搜集及数据筛选条件
通过检索国内1978―2017年间已公开发表或出版的桉树人工林生物量文献汇总相关数据,包括研究地点、经纬度、平均胸径、平均树高、树种、林龄、林分密度、林木单株总生物量和各组分(树干、树枝、叶片和地上部分)生物量等。数据筛选严格遵从以下条件:(1)生物量数据为使用平均标准木法测得的实测数据,不包括推算数据;(2)文献中研究对象为桉树人工林种植苗纯林,不包括混交林和萌芽林;(3)未受到病虫害、台风及间伐等严重干扰;(4)林龄在1 ~ 7 a生之内;(5)非超高密度能源林;(6)除常规的基肥和追肥外无特殊的施肥措施。最终,共筛选出127组生物量数据,数据来源情况见表1。

表1 数据来源概况
1.2 生物量异速生长方程
将收集整理后的数据按照各项指标建立数据库,其中,(胸径)和(树高)作为2个基本变量,是最常见的用于估算林木生物量的林分结构指标。为了满足方差齐性,将最常见的幂函数关系作对数转换,对于转换过程中引入的系统偏差,提供修正因子=EXP(2/2)[52],以消除系统偏差。主要使用以下方程进行模型拟合[22]:
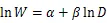
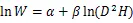
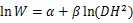
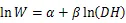
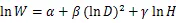


式中:为组分或单株生物量(kg);为胸径(cm);为树高(m)。随机抽取105组数据进行异速生长方程的构建,根据拟合方程的决定系数2和估计值标准误差来选择最佳模型。
1.3 生物量估算参数
目前,IPCC法利用生物量估算参数估算森林生物量主要使用以下公式:
=××(1+) (9)
=×××(1+) (10)
式中:为林分生物量(t×hm-2);为生物量转化与扩展系数,即地上生物量与立木蓄积量之比(t×m-3);为生物量扩展系数,即地上生物量与树干生物量之比,无量纲;为木材密度(t×m-3);为根茎比,即地下生物量与地上生物量之比,无量纲。为林分蓄积量(m3×hm-2)。对于缺失林分蓄积量但提供林林分平均胸径(,cm)、平均树高(,m)和林分密度(,株×hm-2)的文献,通过公式=1/3π(/2)2进行推算,指利用、和推算蓄积量与文献中实测蓄积量的换算系数。由于本研究未涉及木材密度,故仅使用随机抽取的105组数据计算使用公式(9)估算生物量。
使用未参与建模的27组数据对以上估算方法进行检验对比。
1.4 数据处理
数据整理建库采用Excel 2016完成,生物量异速生长方程拟合采用SigmaPlot 14.0和SPSS 19.0软件完成。
2 结果与分析
2.1 桉树生物量异速生长方程
通过使用胸径和树高因子作为单一自变量或组合自变量进行生物量异速方程拟合(表2),拟合方程均达到极显著水平(<0.001)。对于单因子模型,胸径因子对地上部分、地下部分和单株生物量的拟合优度均高于树高因子。对于双因子模型,树高因子的添加并未提高地上和地下部分生物量的拟合优度,对单株生物量的拟合优度提高了0.7%~1.5%

表2 桉树生物量异速生长方程构建
2.2 模型指标的综合评价
2和是回归模型的最常用指标,可以反映模型的拟合优度,也可以反映自变量的贡献率和因变量的离差状况。由表2可知,各生物量模型的评价指标和2数值相近,难以直观确定模型的优劣程度,故需对模型进行综合评价[53]。首先,对逆向指标进行一致化处理(即绝对值取倒数),再对一致化处理后的数据和正向指标2进行Z-score标准化处理。根据各指标的标准差占所有指标标准差的权重计算各指标的权重系数,模型的综合评价结果见表3。模型(7)对地上部分生物量预估效果最好,模型(2)对地下部分生物量预估效果最好,模型(3)对单株生物量预估效果最好。
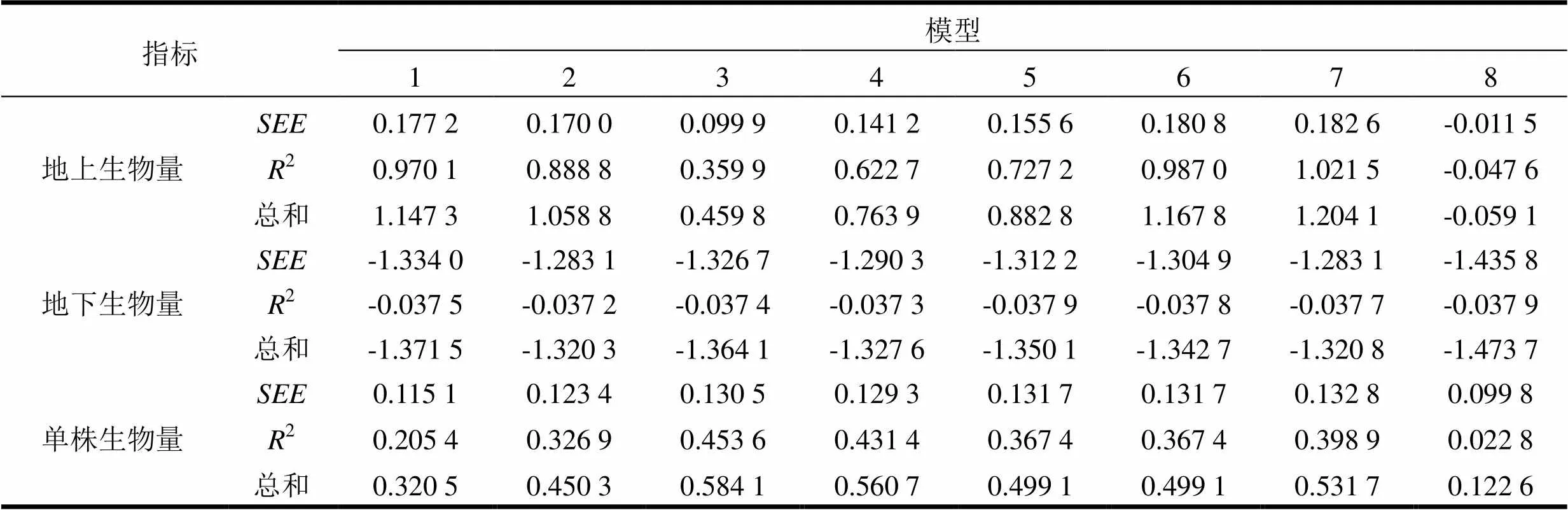
表3 模型综合评价结果
2.3 桉树生物量模型检验及生物量估算相容性
基于27组未参与建模数据,使用方程(7)、(2)和(3)分别估算桉树地上生物量、地下生物量和单株生物量,再对校正后的数据与实测值进行比较。结果发现,地上和单株生物量估算值与实测值的决定系数2分别为0.990和0.968,接近1:1直线。地下生物量估算值与实测值决定系数较低,为0.905。地上、地下和单株生物量估算值随胸径的增加相比于实际值会出现偏小的趋势,在地下和单株生物量表现尤为明显。通过不同模型估算的地上和地下生物量之和,与单株生物量模型估算值进行比较。分量之和与单株生物量估算值间的线性关系决定系数为0.982,接近1:1直线。单株生物量估算值随胸径的增加相比于分量之和会出现偏小的趋势。
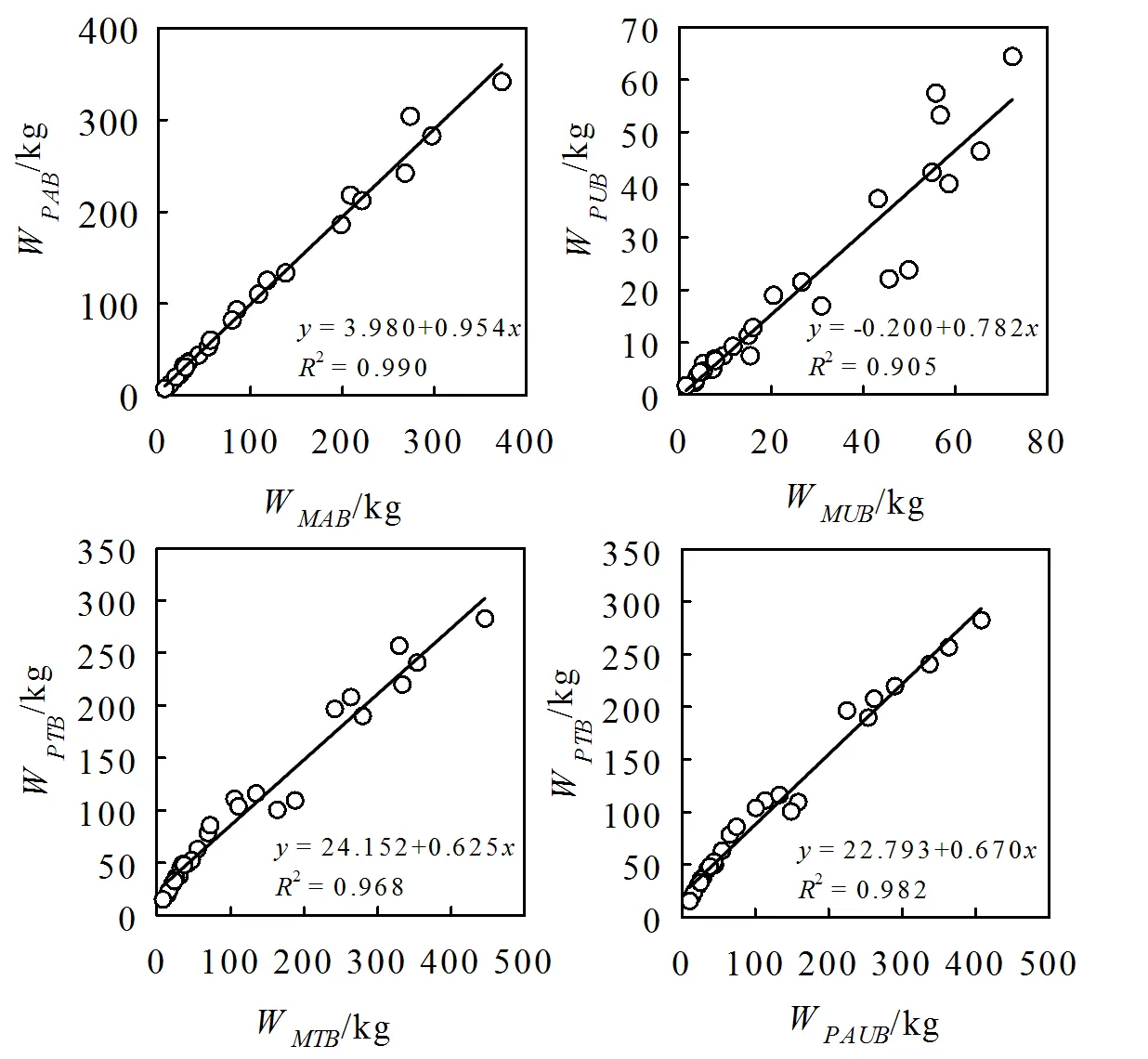
图1 估算生物量相容性及模型检验
注:W,地上生物量实测值;W,地上生物量估算值;W,地下生物量实测值;W,地下生物量估算值;W,单株生物量实测值;W,单株生物量估算值;W,地上和地下生物量估算值之和。
2.4 桉树生物量估算参数
桉树人工林和的平均值分别为0.634 1 (n=65,=0.132)和0.205 6(n=76,=0.089)。生物量参数法是根据公式(9)推算出验证数据组的林分生物量,生物量异速生长方程法是通过模型(3)推算的单株生物量与林分密度乘积得到林分生物量,将两者同时与林分生物量实测值进行比较,结果见图2。IPCC法和生物量异速生长方程法估算值与实测值的线性关系决定系数分别为0.953和0.874,IPCC法更接近1:1直线。

图2 2种生物量估算方法的对比
3 讨论与结论
异速生长方程是最常用的林木生物量估算方法之一。在已构建的异速生长模型中,自变量除了常见的胸径和树高[22, 54]之外还有其他测树因子。自变量的添加可以在一定程度提高构建模型的精度[55],但同时也会增加获取测树因子的难度和引入较多误差。本研究中,单自变量模型的拟合效果均表现为胸径高于树高。在胸径和树高双自变量模型中,树高因子的添加未能提高地上和地下部分生物量的拟合优度,对单株生物量的拟合优度提高了0.7%~1.5%。因此,在建立高精度模型的自变量选择时应该兼顾实用性原则[8]。各方程对地上和单株生物量的拟合优度较高,分别在0.838~0.946和0.929~0.957之间,对地下生物量的拟合优度较低,仅0.647~0.668。造成的原因可能是地下部分生物量(即根生物量)测定过程中,由于不同试验区的土壤条件、根系挖掘方式等破坏了根系完整性,加之取样和测定标准不同引起地下生物量测定的较大误差。因此,在对林木根生物量进行测定的过程中,应该适当选择根系生物量及结构调查方法[56],尽量减少测定误差。
通过对8种模型的评价指标的比较分析及模型的综合评价,发现模型(7)、 (2)和(3)的评价指标的值较大,说明其具有更好的预估效果[53]。通过对比地上、地下和单株生物量预测值与实测值的偏离程度来进一步验证模型的优劣。本研究中,地上、地下和单株生物量预测值与实测值线性关系的决定系数均在90%以上,说明模型(7)、(2)和(3)分别对地上、地下和单株生物量的预测效果较好。模型(7)和(2)估算的地上和地下生物量之和与模型(3)估算的单株生物量的一致性可以反映模型的相容性。本研究中,地上和地下生物量估算值之和与单株生物量估算值线性关系的决定系数达到0.982,说明模型间具有较高的相容性。
2种方法的生物量估算值随胸径的增大出现比实测值偏大的趋势,这可能是因为样本数据中中林龄和幼龄林数据较多,拟合方程数据中有大部分平均木数据,而桉树生长达到一定阶段后生长较为缓慢,生物量不会显著增加。对异速生长方程拟合的单株生物量结合林分密度进行换算后与IPCC法对比发现,IPCC法估算得到的生物量估算值较异速生长方程法更接近实测值,更适合区域尺度估算桉树人工林生物量。
[1] 徐耀粘,江明喜.森林碳库特征及驱动因子分析研究进展[J].生态学报,2015,35(3):926-933.
[2] DIXON R K, SOLOMON A M, BROWN S, et al. Carbon pools and flux of global forest ecosystems.[J]. Science, 1994,263(5144):185-190.
[3] 张宇,岳祥华,漆良华,等.利用异速生长关系和地统计方法估算武夷山南麓毛竹林生物量[J].生态学杂志,2016,35(7):1957-1962.
[4] CANADELL J G, QUERE C L, RAUPACH M R, et al. Contributions to Accelerating Atmospheric CO₂ Growth from Economic Activity, Carbon Intensity, and Efficiency of Natural Sinks[J]. Proceedings of the National Academy of Sciences of the United States of America, 2007,104(47):18866-18870.
[5] PAN Y, BIRDSEY R A, FANG J, et al. A Large and Persistent Carbon Sink in the World's Forests[J]. Science, 2011,333(6045):988-993.
[6] 汪珍川,杜虎,宋同清,等.广西主要树种(组)异速生长模型及森林生物量特征[J].生态学报,2015,35(13):4462-4472.
[7] WEST P W. Tree and Forest Measurement[M]. Berlin Heidelberg :Springer, 2009.
[8] 罗云建,张小全,王效科,等.森林生物量的估算方法及其研究进展[J].林业科学,2009,45(8):129-134.
[9] SALIS S M, ASSIS M A, MATTOS P P, et al. Estimating the aboveground biomass and wood volume of savanna woodlands in Brazil's Pantanal wetlands based on allometric correlations[J]. Forest Ecology and Management, 2006, 228(1/3): 61-68.
[10] NIKLAS K J. Plant allometry: The scaling of form and process[M]. Chicago: The University of Chicago Press, 1994.
[11] CLAIR J B S T. Family differences in equations for predicting biomass and leaf area in Douglas-Fir (var)[J]. Forest Science, 1993,39(4):743-755.
[12] 左舒翟,任引,王效科,等.中国杉木林生物量估算参数及其影响因素[J].林业科学,2014,50(11):1-12.
[13] 陈传国.东北主要林木生物量手册[M].北京:中国林业出版社,1989.
[14] 冯宗炜.中国森林生态系统的生物量和生产力[M].北京:科学出版社,1999.
[15] 胥辉.林木生物量模型研究评述[M].昆明:云南科技出版社,2002.
[16] 罗云建,王效科,逯非.中国主要林木生物量模型手册[M]. 北京:中国林业出版社,2015.
[17] WANG C. Biomass allometric equations for 10 co-occurring tree species in Chinese temperate forests[J]. Forest Ecology and Management, 2006,222(1):9-16.
[18] JENKINS J C, CHOJNACKY D C, HEATH L S, et al. National-scale biomass estimators for United States tree species[J]. Forest Science, 2003,49(1):12-35.
[19] TER-MIKAELIAN M T, KORZUKHIN M D. Biomass equations for sixty-five North American tree species[J]. Forest Ecology and Management, 1997,97(1):1-24.
[20] ZIANIS D, MENCUCCINI M. On simplifying allometric analyses of forest biomass[J]. Forest Ecology and Management, 2004, 187(2):311-332.
[21] MONTAGU K D, DUTTMER K, BARTON C V M, et al. Developing general allometric relationships for regional estimates of carbon sequestration—an example using Eucalyptus pilularis from seven contrasting sites[J]. Forest Ecology and Management, 2005,204(1):115-129.
[22] WILLIAMS R J, ZERIHUN A, MONTAGU K D, et al. Allometry for estimating aboveground tree biomass in tropical and subtropical eucalypt woodlands: towards general predictive equations[J].Australian Journal of Botany, 2005,53(7):607-619.
[23] IPCC. Good practice gudiance for land use, land-use change and forestry[R].Japan: Institute of Global Environment Strategies, 2003.
[24] IPCC. IPCC guidelines for national greenhouse gas inventories: agriculture, forestry and other land use[R]. Japan: Institute of Global Environment Strategies, 2006.
[25] 潘嘉雯,林娜,何茜,等.我国3个桉树人工林种植区生产力影响因素[J].生态学报,2018,28(19):6932-6940.
[26] 张利丽,王志超,陈少雄,等.不同林龄尾巨桉人工林的生物量分配格局[J].西北农林科技大学学报(自然科学版),2017,45(6):61-68.
[27] 韩斐扬,周群英,陈少雄,等.2种桉树不同林龄生物量与能量的研究[J].林业科学研究,2010,23(5):690-696.
[28] 周建辉,郑磊,张婧,等.不同林龄尾巨桉林木碳贮量研究[J].桉树科技,2013,30(4):11-14.
[29] 周群英,陈少雄,韩斐扬,等.短周期尾巨桉能源林生物量与能量特征研究[J].热带亚热带植物学报,2013,21(1):45-51.
[30] 郑海水,曾杰,周再知,等.胶园防护林更新树种的选择[J].广东林业科技,1997,13(1),15-21.
[31] 韩斐扬,周群英,陈少雄.6年生11种桉树无性系生物量与炭化研究[J].桉树科技,2012,29(3):1-8.
[32] 杨卫星,何斌,卢开成,等.桂西南连续年龄序列尾巨桉人工林的生物生产力[J].农业研究与应用,2016(3):6-11.
[33] 叶绍明,郑小贤,谢伟东,等.萌芽更新与植苗更新对尾巨桉人工林收获的影响[J].南京林业大学学报(自然科学版),2007,31(3):43-46.
[34] 陈婷,温远光,孙永萍,等.连栽桉树人工林生物量和生产力的初步研究[J].广西林业科学,2005,34(1):8-12.
[35] 朱宇林,温远光,曹福亮,等.短周期尾巨桉连栽林分生产力的研究[J].江西农业大学学报, 2006,28(1):90-94.
[36] 李春宁,韦建宏,付军,等.2种更新方式尾巨桉中龄林地上部分生物生产力比较分析[J].农业研究与应用,2017(6):17-20.
[37] 吴华静,田丰,桂凌健,等.南宁七坡林场尾巨桉人工林生物量的初步研究[J].广西科学院学报,2014,30(4):233-237.
[38] 邓龙江.连栽对尾巨桉萌芽林生物量和生产力的影响[D].南宁:广西大学,2015.
[39] 梁宏温,温远光,吴国喜,等.连栽对尾巨桉短轮伐期人工林生长量和生产力动态的影响[J].福建林业科技,2008,35(3):14-18.
[40] 张琼.桉树工业原料林生态系统生物量和碳贮量初步研究[D].福州:福建农林大学,2005.
[41] 朱宾良,李志辉,陈少雄.不同林分密度对尾巨桉生物产量及生产力的影响研究[J].湖南生态科学学报,2007,13(4):11-14.
[42] 李志辉,陈少雄,谢耀坚,等.林分密度对尾巨桉生物量及生产力的影响[J].中南林业科技大学学报,2008,28(4):49-54.
[43] 李况.不同年龄桉树人工林生态系统碳氮储量分配特征[D].南宁:广西大学,2013.
[44] 刘正富.巨尾桉人工林生长、生物量及植物多样性的动态变化[D]. 南宁:广西大学,2012.
[45] 陶玉华,隆卫革,马麟英,等.柳州市马尾松、杉木、桉树人工林碳储量及其分配[J].广东农业科学,2011,38(22):42-45.
[46] 骆栋卿.高峰林场巨尾桉密植林生物量生产力及土壤水文功能研究[D].南宁:广西大学,2016.
[47] 韦宇静,梁士楚,黄雅丽,等.巨尾桉与几种阔叶树和针叶树碳储量的比较研究[J].广西科学院学报,2014,30(4):229-232.
[48] 左花.不同经营周期巨尾桉人工林的生物量和碳储量[D].南宁:广西大学,2015.
[49] 卢婵江.广西东门不同林龄巨尾桉人工林的生物生产力及经济效益分析[D].南宁:广西大学,2017.
[50] 李跃林,李志辉,谢耀坚.巨尾桉人工林养分循环研究[J].生态学报,2001,21(10):1734-1740.
[51] 张胜伟.巨尾桉工业原料林生物量研究[D].昆明:昆明理工大学,2008.
[52] SPRUGEL D G. Correcting for Bias in Log-Transformed Allometric Equations[J]. Ecology, 1983,64(1):209-210.
[53] 金贞珍.关于多指标综合评价方法及其权数问题的讨论[D].延边:延边大学,2007.
[54] PILLI R, ANFODILLO T, CARRER M. Towards a functional and simplified allometry for estimating forest biomass[J]. Forest Ecology and Management, 2006,237(1):583-593.
[55] SAINT-ANDRE L, M Bou A T, Mabiala A, et al. Age-related equations for above- and below-ground biomass of ahybrid in Congo[J]. Forest Ecology and Management, 2005,205(1/3):199-214.
[56] 张小全,吴可红.树木细根生产与周转研究方法评述[J].生态学报,2000,20(5):875-883.
Allometry Equations for EstimatingTree Biomass and Comparison with IPCC Method
JIE Fan1, DU Apeng2, ZHU Wankuan2
In order to construct allometric equations for large scaleplantations, biomass data were obtained from published papers and subject to meta-analysis to obtain regression relationships of various factors with aboveground, belowground and total-tree biomass. The results showed that a single independent variable equation based on DBH performed better than one based on tree height. In double independent variable models, the use of tree height in combination with DBH only improved theRof total-tree biomass by 0.7% to 1.5%, but did not improve correlations obtained with other component equations. Verification analysis of predicted and measured values of the models showed that the equation ln= -2.833 + (2.301*ln) + 0.3521*lnhas the best predictive ability for aboveground biomass with an accuracy of 94.6%, while ln=-5.175 + 0.939*lnDHhas the best predictive ability for underground biomass with a correlation of 66.8%, and ln= -2.960 + 0.896*lnDHhas the best predictive ability for total-tree biomass with a correlation of 95.5%. The derived component model was comparable with total-tree biomass model. The means of(Biomass Conversion and Expansion Factor)and(Root:shoot Ratio) were 0.634 (n=65, SD=0.132) and 0.206 (n=76, SD=0.089), respectively. The accuracy of the forest biomass estimation by the IPCC method was higher than that of the allometric equation, which was 95.3%. Therefore, it is recommended to use the IPCC(Intergovernmental Panel on Climate Change) method for estimating the biomass ofplantations on regional scales.
plantation; biomass; allometry equations; regional scale
S714.5
A
广东省林业科技创新项目(2018KJCX014);2016年省级生态公益林激励性补助资金项目(2016-03);广西科技重大专项(桂科AA172004087-9);广东湛江桉树林生态系统国家定位观测研究站运行补助
揭凡(1986― ),男,助理工程师,主要从事桉树人工林培育研究,E-mail: 51256163@qq.com
竹万宽(1989― ),男,硕士,研究实习员,主要从事桉树人工林生态定位监测研究,E-mail: zwk_2015@163.com
猜你喜欢
环境保护与循环经济(2021年7期)2021-11-02
哈尔滨轴承(2020年1期)2020-11-03
中国奶牛(2019年10期)2019-10-28
儿童故事画报·发现号趣味百科(2019年9期)2019-02-02
环球时报(2019-01-03)2019-01-03
电子制作(2018年23期)2018-12-26
现代园艺(2017年19期)2018-01-19
现代园艺(2017年21期)2018-01-03
广西林业科学(2016年2期)2016-03-20
广西林业科学(2016年2期)2016-03-20
