
转双价基因棉SGK321不同秸秆还田量对土壤线虫群落的影响
2019-04-22 06:40蔡连贺刘瑞华杨殿林赵建宁
农业环境科学学报 2019年4期
李 静,蔡连贺,刘瑞华,风 春,李 刚,杨殿林,赵建宁*
(1.东北农业大学资源与环境学院,哈尔滨 150030;2.农业农村部环境保护科研监测所,农业农村部产地环境污染防控重点实验室/天津市农业环境与农产品安全重点实验室,天津 300191;3.中国农业科学院棉花研究所,河南 安阳 455000;4.呼和浩特第九中学,呼和浩特 010000)
全球转基因作物种植面积不断扩大,已经从1996年的170万hm2上升至2017年的1.898亿hm2[1]。棉花是全球主要经济、纤维作物之一,2018年,我国转基因棉花的种植面积约300万hm2,占全国棉花播种总面积的80%以上,是我国种植面积最大的转基因作物。但转基因作物在为社会带来巨大经济效益的同时,其潜在的生态环境安全风险也被越来越多的人们所关注[2-3]。
土壤线虫是地球上数量丰富的后生动物之一,在维持土壤生态系统稳定、促进物质循环和能量流动等方面发挥着重要作用[4],常被作为监测农业管理措施干扰和评价土壤环境质量的重要指示生物[5-7]。转Bt作物的目的基因可以通过花粉、秸秆还田、根系分泌等方式进入土壤[8-9],会大幅增加土壤中Bt蛋白的含量[10-12],进而可能会对土壤动物如土壤线虫产生影响。段杉[13]研究发现,SGK321与石远321相比在蕾期和吐絮期的线虫数量存在显著差异,但其他一些研究并未发现转Bt棉花对土壤线虫群落有显著影响[14-16]。对转Bt基因玉米MON810的研究表明,其土壤线虫密度显著高于常规玉米[17]。Gu等[18]研究发现秸秆还田对总丰度无显著影响,但随着秸秆还田量的增加,食细菌性线虫丰度逐渐增高,杂食性线虫丰度逐渐减少。有研究发现,秸秆还田后土壤食细菌性线虫、食真菌性线虫和有较高c-p值的杂食/捕食性线虫数量增加,而植食性线虫数量降低[19-20]。牟文雅等[21]研究发现秸秆对线虫数量及物种数有一定影响,可改变线虫群落组成。国内外已有很多文献报道了转基因作物对土壤线虫群落结构的影响和传统作物秸秆还田对土壤线虫群落结构的影响。秸秆还田是现代农业生产过程中的一项重要技术,还田后的秸秆腐解对于促进作物生长、改善和提高土壤质量具有重要作用。因此研究转基因棉花秸秆还田腐解后对土壤线虫群落结构的影响具有十分重要的意义。
本研究以转双价(Bt+CpTI)基因棉SGK321及其常规棉石远321为对象,比较不同棉花秸秆还田量下、不同生育期中两种棉花土壤线虫密度及其群落的动态变化情况,探讨转双价(Bt+CpTI)基因棉秸秆还田对土壤线虫群落结构的影响,以期深入了解转基因棉对土壤生态环境的影响和构建转基因棉生态安全监测指标,为转基因棉种植的土壤生态安全性评价提供科学依据。
1 材料与方法
1.1 试验材料
供试棉花品种为转双价(Bt+CpTI)基因棉SGK321及其常规棉石远321(简称SGK321和石远321),均由中国农业科学院植物保护研究所提供。
供试花盆上口直径20 cm、底部直径16 cm、高23 cm。供试土壤为潮土,取自天津市武清区常规农田土壤,其基本理化性质:全磷含量0.79 g·kg-1,全氮含量 0.63 g·kg-1,有机质含量 18 g·kg-1,pH 8.24。每盆装过2 mm筛后充分混匀的干基土5.6 kg,为模拟正常土壤温度,将花盆埋入土壤中,埋深15 cm。
1.2 试验设计
试验在农业农村部环境保护科研监测所(天津市南开区,39°5′8.28″N,117°8′39.12″E)网室内进行。试验区属温带大陆性季风气候,年均气温和年均降水量分别为15℃和600 mm。该试验是不完全随机区组实验,设置5个还田梯度,按照土层厚度换算而来。每公顷0~20 cm耕层土壤约有2.25×106kg,还田梯度分别为 0、2250、4500、6750 kg·hm-2和 9000 kg·hm-2,分别代表不还田、半量还田、全量还田、1.5倍还田量和2倍还田量。换算成盆栽试验的还田梯度为0(对照)、1.0、2.0、3.0 g·kg-1和 4.0 g·kg-1。试验所用秸秆为SGK321和石远321的秸秆,用粉碎机粉碎后过20目筛,按照试验设置分别与土壤均匀混合,用于转基因棉花SGK321及其常规棉石远321棉花的种植。每个还田处理设有4个重复,每个重复5盆,共200盆盆栽。于2015年4月24日播种,每盆放置6粒棉花种子,待出苗后定苗至每盆2株,日常管理过程中不喷洒农药。分别于播种后60 d(蕾期)和90 d(花铃期)两个时期进行土壤样品采集。
1.3 土壤样品的采集
去除花盆表面杂草和落叶,用土钻取距离棉花主茎2 cm的土壤,深度15 cm,每盆取2钻,5盆混为一个样品。样品混匀后除去植物根系、残渣等,放入自封袋带回实验室,于4℃保存,并在两周内完成线虫分离。
1.4 线虫的分离与鉴定
土壤线虫分离采用Baermann浅盘法[22]:从采集的混匀土壤中取出50 g鲜土,在25℃的环境下静止分离48 h,分离后的土壤浸出液用500目筛过滤,将分离出的线虫经60℃水浴杀死后,保存于4%的福尔马林溶液中[23]。根据土壤的含水量,折合成每100 g干土土壤线虫密度,然后每个样品根据镜检法[24]随机选取100条线虫,使用Nikon公司生产的TS100荧光侧置显微镜进行鉴定。如果样品中线虫不足100条,则所有样品都要鉴定。根据土壤线虫的头部形态学特征,将线虫划分为植食性线虫(Plant-parasites)、食细菌性线虫(Bacterivores)、食真菌性线虫(Fungivores)和杂食/捕食性线虫(Omnivores-predators)等4个营养类群;并根据线虫的食性和生活策略(r-策略和k-策略)将不同线虫类群赋予1~5的c-p值[25]。
1.5 线虫生态指标分析
研究采用生态学评价指数:
(1)Shannon-Wiener多样性指数:H′=-∑PilnPi,Pi=ni/N。式中Pi为样品中属于第i种的个体的比例;ni为第i类群的个体数;N为所有类群的个体总数[26]。
(2)Pielou 均匀度指数:J=H′/lnS。式中H′为Shannon-Wiener多样性指数;S为鉴定分类单元的数目[27]。
(3)Margalef丰富度指数:SR=(S-1)/lnN。式中S为鉴定分类单元的数目;N为鉴定的线虫个体数目[28]。
(4)线虫通路指数:NCR=NB/(NB+NF)。式中NB指食细菌性线虫的土壤线虫密度;NF指食真菌性线虫的土壤线虫密度[29]。
(5)瓦斯乐斯卡指数:WI=(NF+NB)/NPP。式中NF和NB同上;NPP为植食性线虫密度[30]。
(6)自由生活线虫成熟度指数:MI=∑(vi×fi)。式中vi为自由生活线虫的c-p值;fi为第i种线虫的个体数占自由生活线虫的比例[31]。
1.6 数据分析
数据分析采用SPSS软件(16.0版,SPSS Inc.),采用T检验分析转基因棉与常规棉之间的差异,不同还田处理与不同生育期生态指数差异采用单因子方差分析法(One-way ANOVA),品种、秸秆还田量和生育期之间的交互作用采用多因子方差分析法(Threeway ANOVA),处理组间显著检验采用Duncan法。
2 结果与分析
2.1 土壤线虫密度
SGK321土壤线虫密度仅在蕾期3.0 g·kg-1还田处理时显著高于石远321(P<0.05),其他处理差异不显著(图1)。不还田时,两个时期SGK321土壤线虫密度均低于石远321,但无显著差异。
如图1所示,蕾期,随着秸秆还田量的增加,石远321土壤线虫密度逐渐增加,在4.0 g·kg-1还田处理时密度达到最大值。SGK321土壤线虫密度在3.0 g·kg-1还田处理时达到最大值,且秸秆还田后土壤线虫密度均高于不还田处理。花铃期,随着秸秆还田量的增加,石远321和SGK321土壤线虫密度均逐渐增加,在4.0 g·kg-1还田处理时密度达到最大值,同时SGK321的线虫密度均高于石远321,但无显著差异。
花铃期土壤线虫密度相比于蕾期,石远321土壤线虫密度在0、2.0 g·kg-1处理中减少,其他处理中增加;SGK321土壤线虫密度在3.0 g·kg-1还田处理中减少,其他处理中增加。
2.2 土壤线虫群落组成
如表1和表2所示,SGK321两个时期共鉴定出土壤线虫19科34属,其中食细菌性线虫13属,食真菌性线虫3属,植食性线虫11属,杂食/捕食性线虫7属。石远321两个时期共鉴定出土壤线虫19科37属,其中食细菌性线虫13属,食真菌性线虫3属,植食性线虫12属,杂食/捕食性线虫9属。SGK321和石远321的优势属均是螺旋属Helicotylenchus,其中SGK321土壤中螺旋属在蕾期2.0 g·kg-1与1.0、3.0、4.0 g·kg-1还田处理之间存在显著差异(P<0.05)。石远321土壤中螺旋属在花铃期4.0 g·kg-1与1.0、2.0 g·kg-1还田处理之间存在显著差异(P<0.05)。
蕾期,拟丽突属Acrobeloides、原杆属Protorhabditis等6属在两个棉花品种各还田处理之间均存在显著差异(P<0.05);SGK321土壤线虫中茎属Ditylenchus、垫刃属Tylenchus、异皮属Heterodera、锉齿属Mylonchulus等10属在各还田处理之间存在显著差异(P<0.05),石远321土壤线虫中孔咽属Aporcelaimus、真矛线属Eudorylaimus等7属在各还田处理之间存在显著差异(P<0.05)。花铃期,鹿角唇属Cervidellus、茎属等5属在两个棉花品种各还田处理之间均存在显著差异(P<0.05);SGK321土壤线虫中丝尾垫刃属Filenchus和原杆属在各还田处理之间存在显著差异(P<0.05);石远321土壤线虫中棱咽属Prismatolaimus、矮化属Tylenchorhynchus等10属在各还田处理之间存在显著差异(P<0.05)。石远321土壤线虫中拟丽突属、棱咽属和矮化属等5属在两个生育期各还田处理中均存在显著差异(P<0.05);SGK321土壤线虫中丝尾垫刃属、原杆属、鹿角唇属、垫刃属和茎属在两个生育期各还田处理中均存在显著差异(P<0.05)。
与不还田相比,三等齿属Pucephalobus、丽突属Acrobeles等11属只出现在秸秆还田处理中。其中剑属Xiphinema、三等齿属Pucephalobus、微矛线属Microdorylaimus、前矛线属Prodorylaimus仅出现在石远321的还田处理中(表1),丽突属仅出现在SGK321的4.0 g·kg-1还田处理中(表2),其他属分布在两个时期的不同还田处理中。
2.3 土壤线虫的营养类群结构
SGK321和石远321土壤线虫在采样的两个生育期均以食细菌性线虫(蕾期:27.07%~39.93%;花铃期:32.73%~52.7%)和植食性线虫(蕾期:25.63%~55.03%;花铃期:14.98%~42.89%)为主要营养类群(图2)。不还田时,蕾期SGK321中食细菌性和食真菌性线虫丰度高于石远321,杂食/捕食性和植食性线虫丰度低于石远321;花铃期SGK321中杂食/捕食性线虫丰度显著高于石远321(P<0.05)。
蕾期,在2.0 g·kg-1还田处理时,除石远321土壤中食细菌线虫丰度显著高于SGK321(P<0.05),食真菌线虫丰度高于SGK321,但无显著差异外,其他处理SGK321土壤中食细菌性和食真菌性线虫丰度均高于石远321,但无显著差异;两种棉花土壤中食细菌性和食真菌性线虫在不同秸秆还田处理间均无显著差异。SGK321的植食性线虫丰度在2.0 g·kg-1还田处理时显著高于石远321(P<0.05);石远321土壤中植食性线虫丰度在不还田时显著高于3.0 g·kg-1和4.0 g·kg-1还田处理(P<0.05);SGK321土壤中植食性线虫丰度在秸秆还田后各处理间均存在显著差异(P<0.05)。石远321中杂食/捕食性线虫丰度均高于SGK321,但无显著差异,两种棉花不同秸秆还田处理间也均无显著差异。
花铃期,除2.0 g·kg-1还田处理外,石远321的食细菌性线虫丰度均高于SGK321,且在4.0 g·kg-1还田处理时存在显著差异(P<0.05);石远321土壤中食细菌性线虫丰度在4.0 g·kg-1还田处理时显著高于2.0 g·kg-1和不还田处理(P<0.05);SGK321土壤中食细菌性线虫丰度在3.0 g·kg-1还田处理时显著高于1.0 g·kg-1和不还田处理(P<0.05)。石远321土壤中食真菌性线虫丰度均高于SGK321,在3.0 g·kg-1还田处理时存在显著差异(P<0.05),且在3.0 g·kg-1还田处理时显著高于其他还田处理(P<0.05);SGK321土壤中食真菌性线虫丰度在各处理间无显著差异。除2.0 g·kg-1还田处理,SGK321土壤中植食性线虫丰度均高于石远321,且在3.0、4.0 g·kg-1还田处理时存在显著差异(P<0.05);石远321土壤中植食性线虫丰度在2.0 g·kg-1还田处理时显著高于4.0 g·kg-1还田处理(P<0.05);SGK321土壤中植食性线虫丰度在1.0 g·kg-1还田处理时显著高于其他处理(P<0.05)。石远321土
壤中杂食/捕食性线虫丰度在1.0、3.0 g·kg-1还田处理时低于SGK321,但差异不显著;石远321土壤中杂食/捕食性线虫丰度在2.0 g·kg-1还田处理时显著高于3.0 g·kg-1还田处理(P<0.05);SGK321土壤中杂食/捕食性线虫丰度在不还田处理时显著高于3.0、4.0 g·kg-1还田处理(P<0.05)。
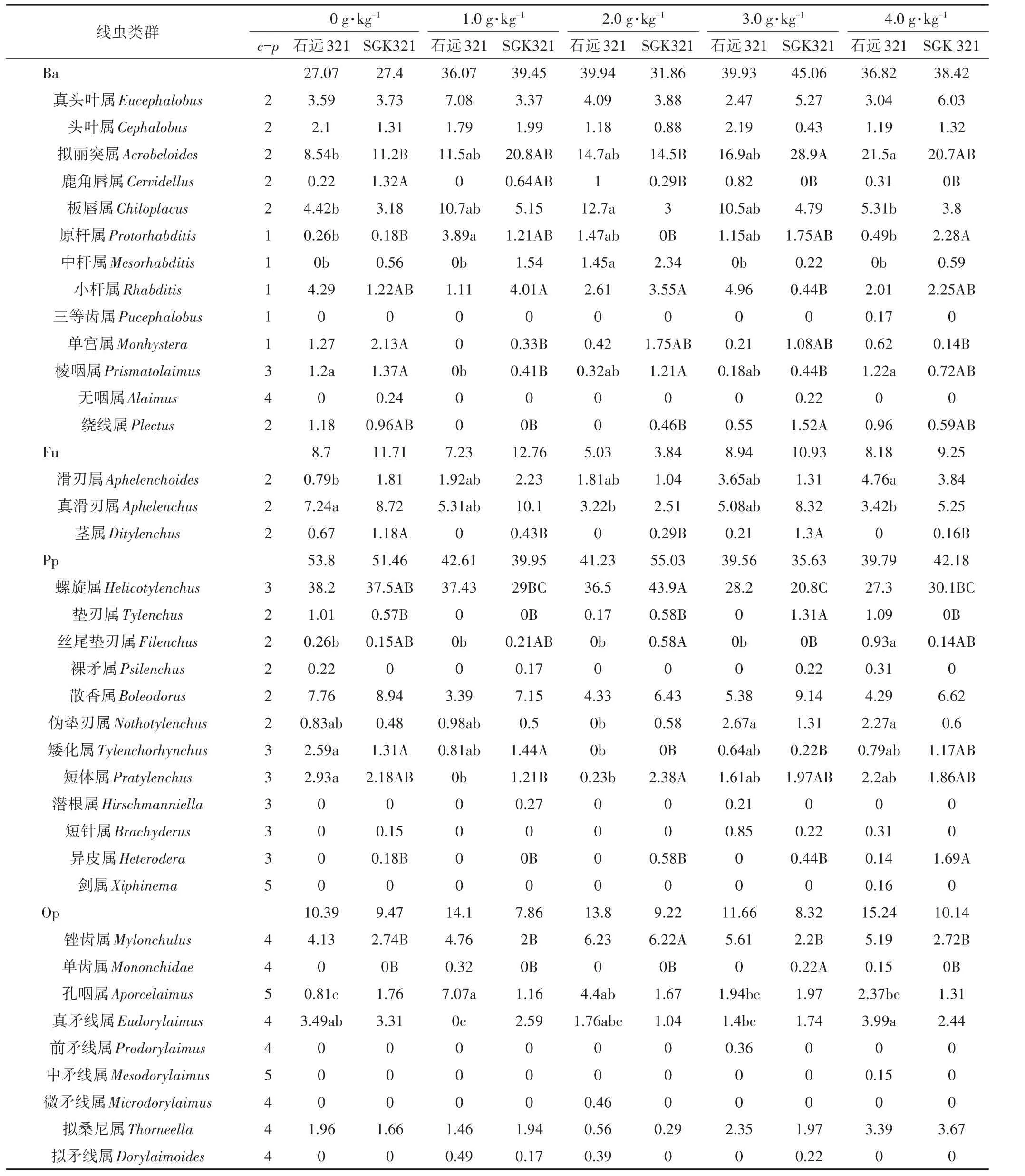
表1 蕾期不同还田处理土壤线虫的优势度Table 1 The dominance of soil nematodes under different treatments at budding stage
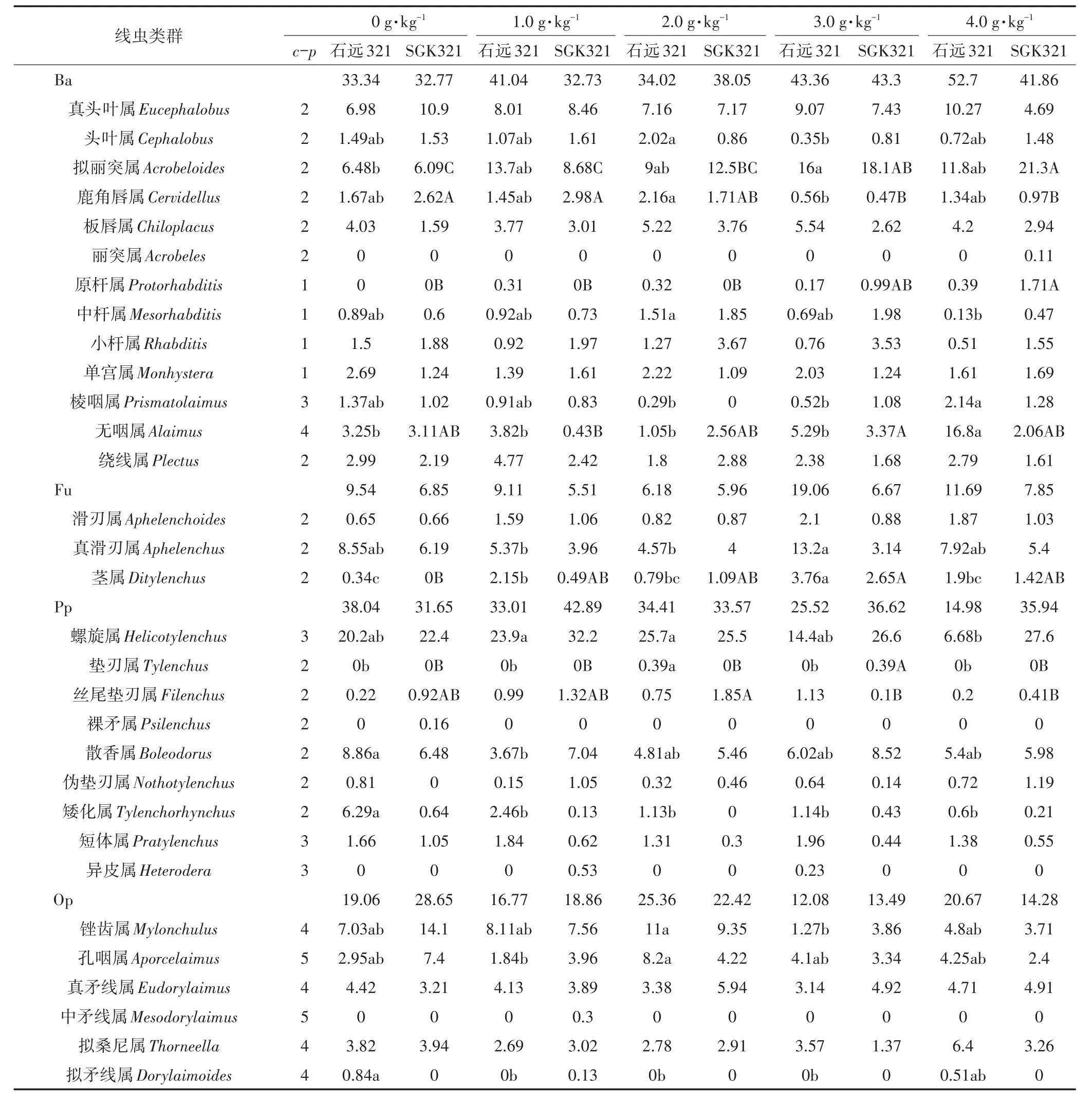
表2 花铃期不同还田处理土壤线虫的优势度Table 2 The dominance of soil nematodes under different treatments at flowering and boll-forming stage
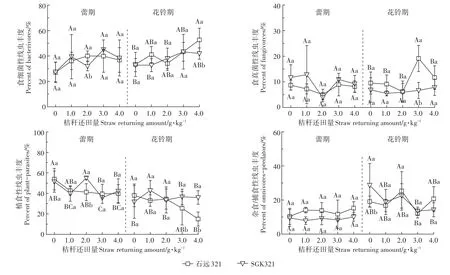
图2 蕾期和花铃期不同秸秆还田处理下各营养类群丰度Figure 2 Abundance of different nutrient groups in different returning amounts at budding stage and flowering and boll-forming stage
2.4 土壤线虫群落生态指数
SGK321的SR指数在蕾期不还田处理时显著高于石远321(P<0.05)(表3)。石远321的J指数在蕾期3.0 g·kg-1还田处理和花铃期1.0 g·kg-1还田处理时显著高于SGK321(P<0.05)。H′指数两品种间不存在显著性差异。石远321不同还田处理中的SR、H′指数在蕾期4.0 g·kg-1还田处理时显著高于1.0 g·kg-1还田处理(P<0.05),J指数在各处理间均无显著差异。SGK321不同还田处理中的H′指数在蕾期3.0 g·kg-1还田处理时显著高于2.0 g·kg-1还田处理(P<0.05),SR、J指数在各处理间均无显著差异。
MI指数[25]是根据线虫的群落结构特征评价人类活动对线虫群落结构的影响,进而可以表明土壤环境所受的干扰程度。石远321的MI指数在蕾期2.0 g·kg-1还田处理时显著高于SGK321,在花铃期1.0、4.0 g·kg-1还田处理时显著高于 SGK321(P<0.05)。蕾期不同还田处理中SGK321土壤线虫的MI指数在1.0、3.0 g·kg-1还田处理时显著高于不还田、2.0 g·kg-1还田处理(P<0.05);花铃期石远321不同还田处理的MI指数在4.0 g·kg-1还田处理时显著高于不还田、1.0、2.0 g·kg-1还田处理(P<0.05)。线虫的营养类群结构与土壤碎屑食物网密切相关,常用食细菌和食真菌线虫的比值(即NCR)探测土壤有机质的分解途径[32]。SGK321的NCR指数在花铃期3.0 g·kg-1还田处理时显著高于石远321(P<0.05)。不同还田处理中石远321的NCR指数在蕾期2.0 g·kg-1还田处理时显著高于不还田处理(P<0.05);在花铃期2.0 g·kg-1还田处理时显著高于3.0 g·kg-1还田处理(P<0.05)。不同还田处理中SGK321土壤线虫的NCR指数在蕾期和花铃期各秸秆还田处理间均无显著差异。WI指数用于反映土壤线虫群落组成和土壤的健康程度[21],石远321的WI指数在蕾期2.0 g·kg-1还田处理和花铃期1.0、4.0 g·kg-1还田处理时显著高于 SGK321(P<0.05)。不同还田处理中石远321的WI指数在花铃期4.0 g·kg-1还田处理时显著高于0、1.0、2.0 g·kg-1还田处理(P<0.05)。SGK321的WI指数在蕾期3.0 g·kg-1还田处理时显著高于不还田和2.0 g·kg-1还田处理(P<0.05)。
花铃期相比于蕾期,石远321中除SR、NCR指数外,其他生态指数均升高;SGK321中除H′、J指数外,其他生态指数均降低(表3)。
2.5 棉花品种、秸秆还田量和生育期对线虫数量及生态指标的交互效应
由表4可知,棉花生育期对植食性线虫和杂食/捕食性线虫数量有极显著影响;棉花品种对线虫总量和植食性线虫数量有极显著影响;棉花秸秆还田量对线虫总量、食细菌性线虫及食真菌性线虫数量有极显著影响,对杂食/捕食性线虫数量有显著影响。多因素方差分析表明:棉花品种和生育期对食真菌性和杂食/捕食性线虫数量有显著交互效应;棉花品种和秸秆还田量对线虫总量和植食性线虫数量有极显著交互效应,对食细菌性线虫有显著交互效应。
由表5可知,除NCR外,棉花生育期对SR、H′、J、MI和WI指数均有极显著影响;棉花品种对J指数有极显著影响,对WI指数有显著影响;棉花秸秆还田量对MI、WI指数有极显著影响,对SR、NCR指数有显著影响。多因素方差分析表明:棉花品种和生育期对SR、MI指数有极显著交互效应,对H′、NCR、WI指数有显著交互效应;棉花品种和秸秆还田量对WI指数有显著交互效应;棉花品种、生育期和秸秆还田量对WI指数有显著交互效应。
3 讨论
土壤线虫因其独特的生物特性及其在土壤群落结构中的重要作用,而成为指示土壤生态系统结构和功能变化的有力工具[33]。陈群英等[34]连续两年的转Bt水稻田间种植未对土壤线虫总密度及各营养类群的线虫密度产生显著影响。本研究表明,转双价基因棉和常规棉土壤线虫密度在不还田时无显著差异,这与Yang等[16]的结果一致,说明转双价基因棉花的种植对土壤线虫密度没有显著影响。秸秆还田后,两种棉花的土壤线虫密度随着秸秆还田量的增加而增加,这说明秸秆的施入,为线虫提供了更加充足的营养物质和能量来源,有利于土壤线虫的生长繁殖[21]。
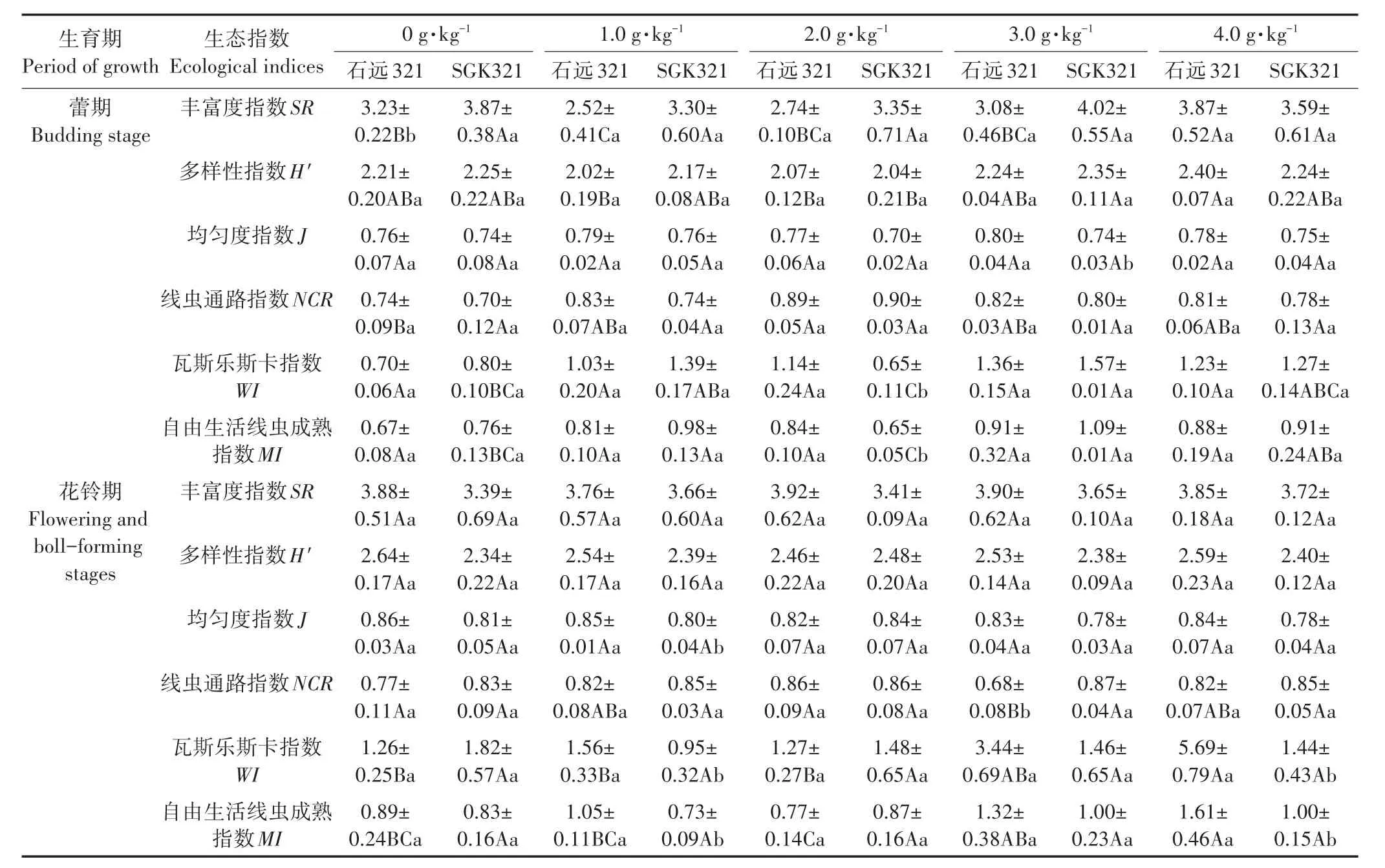
表3 蕾期和花铃期不同还田处理土壤线虫的生态指数Table 3 Ecological indicators of the soil nematode of the different treatments at budding stage and flowering and boll-forming stage

表4 棉花品种、秸秆还田量和生育期对线虫数量的交互效应Table 4 Interactive effects of cotton varieties,straw returning amount and growth period on the number of nematode

表5 棉花品种、秸秆还田量和生育期对线虫生态指标的交互效应Table 5 Interactive effects of cotton varieties,straw returning amount and growth period on nematode ecological indicators
本试验中两个棉花品种,5个还田处理共鉴定出线虫19科38属。秸秆还田对螺旋属和拟丽突属等优势属影响不大,但增加线虫稀有属的数量。说明秸秆还田主要是通过影响稀有类群的数量进而影响线虫群落结构[35]。与不还田处理相比,秸秆还田处理中出现了三等齿属、丽突属等属,表明秸秆还田增加了土壤线虫的丰富度[21],提高了线虫多样性[36]。
土壤线虫群落的营养类群可以有效地反映土壤线虫食物网营养级关系及能量流动途径[29]。戚琳等[37]发现转Bt水稻的田间种植增加植食性线虫丰度。但对转双价基因棉的研究未发现其改变线虫的营养类群[16]。本研究表明,转双价基因棉的种植也没有改变土壤线虫的营养类群结构,各处理间线虫群落组成不存在显著差异,植食性和食细菌性线虫是优势营养类群。食真菌性线虫在两个棉花品种所有处理中均属于最低的营养类群,且秸秆还田后丰度低于不还田,这可能是因为棉花秸秆木质化很强,细菌比较难以利用,真菌在分解棉花秸秆中的作用更加突出。秸秆还田后,常规棉杂食/捕食性线虫丰度增加,这说明秸秆的施用为杂食/捕食性线虫提供食物来源,增加了土壤中原生动物等的密度[38]。
利用线虫的生态指数可以很好地反映不同秸秆还田处理间土壤线虫多样性和群落结构的差异程度[21]。本研究表明,蕾期石远321的SR、H′指数在4.0 g·kg-1与1.0、2.0 g·kg-1还田处理之间存在显著差异,SGK321的H′指数在2.0 g·kg-1与3.0 g·kg-1还田处理之间存在显著差异,这与Zhang等[39]的研究结果相同,说明秸秆还田后,多样性有一定的增加趋势,在一定还田范围内能够提高土壤线虫丰富度。但花铃期两种棉花品种之间土壤线虫H′、SR指数均没有显著差异,表明作物生育期对土壤线虫的影响大于转双价基因棉[34]。MI指数越低,表明土壤环境受到干扰程度越大[21]。本研究中,蕾期SGK321的MI指数在3.0 g·kg-1还田处理与不还田、2.0 g·kg-1还田处理之间均存在显著差异,在花铃期两种棉花的MI指数在3.0、4.0 g·kg-1还田处理显著高于不还田处理,说明较高的秸秆还田量对土壤环境的影响相对更小。蕾期转双价基因棉土壤线虫的MI指数普遍高于常规棉,说明转双价基因棉种植对土壤线虫的干扰较小,这与杨志国等[40]的研究结果相同。NCR指数值为0,代表土壤有机质分解是完全依靠真菌分解途径;若NCR值为1,则表示完全依靠细菌分解途径[29]。WI指数值越大,表明土壤食微线虫丰度越高,土壤健康程度越高。本研究中,NCR值在0.68~0.90间,均大于0.5,表明各秸秆还田处理的盆栽土壤主要依靠细菌分解途径分解有机物,且NCR在秸秆还田条件下显著高于不还田处理,这表明秸秆还田能促进食细菌性线虫繁殖并抑制植食性线虫的生长。除蕾期0、2.0 g·kg-1还田处理和花铃期1.0 g·kg-1还田处理外,WI值均大于1,表明盆栽所用土壤健康状况普遍良好。
多因素方差分析表明,生育期对植食性线虫和杂食/捕食性线虫数量有极显著影响,说明生育期会引起土壤线虫密度的变化,这与李修强等[41]结论一致,对线虫SR、H′、J、MI和WI指数有极显著影响,说明生育期对线虫群落结构组成有影响。品种对线虫总量和植食性线虫数量有极显著影响,对J、WI指数有影响,可能是由于转Bt作物外源基因的插入导致根系分泌物成分发生变化,进而影响线虫群落的数量和组成[8]。秸秆还田量对线虫数量和生态指标均有一定影响,说明秸秆还田增加线虫丰度,丰富线虫种类,有利于土壤生态稳定。生育期与秸秆还田量和生育期、品种与秸秆还田量对线虫群落没有交互作用,说明对线虫数量的影响是相互抵消的。
转基因作物秸秆还田引起的土壤生态系统的变化是一个缓慢的、长期的过程,一年的盆栽试验无法得到全面的反映,需要开展长期的试验研究,以期更全面地评价转基因作物秸秆还田对土壤生态环境的影响。
4 结论
(1)转双价基因棉的种植并未改变土壤线虫的营养类群组成,两种棉花不同还田处理下的多样性指数(H′)、线虫通路指数(NCR)均无显著差异。
(2)秸秆不还田时,转双价基因棉和常规棉土壤线虫密度无显著差异。秸秆还田后随着秸秆还田量的增加,蕾期转双价基因棉土壤线虫数量先升高后降低,常规棉土壤线虫数量逐渐增加。花铃期两种棉花土壤线虫数量逐渐升高。
(3)多因素方差分析表明,生育期、品种和秸秆还田量对线虫数量和生态指数均有一定的影响,生育期与品种和品种与秸秆还田量对线虫数量和生态指数有一定的交互效应,而生育期与秸秆还田量和生育期、品种及秸秆还田量三者之间没有交互作用。
猜你喜欢
今日农业(2022年14期)2022-09-15
新疆农业科学(2022年7期)2022-08-10
少儿科学周刊·儿童版(2022年10期)2022-06-27
农药科学与管理(2021年11期)2022-01-12
西北农业学报(2020年12期)2020-12-14
载人航天(2020年3期)2020-07-02
小读者(2020年4期)2020-06-16
江苏农业科学(2016年2期)2016-04-11
红领巾·萌芽(2015年6期)2015-08-14
小天使·一年级语数英综合(2014年7期)2014-06-26
