
新疆冬小麦过氧化物酶基因 TaPod-D1和 TaPod-A1的等位变异及分布
2019-04-12 08:32战帅帅王丽丽耿洪伟
麦类作物学报 2019年3期
谢 磊,战帅帅,王丽丽,任 毅,时 佳,耿洪伟
(新疆农业大学农学院/新疆农业大学生物技术重点实验室,新疆乌鲁木齐 830052)
面粉色泽是决定小麦品质的重要因素[1],小麦籽粒过氧化物酶(peroxidase, POD)活性是影响面粉及面制品色泽的重要指标之一。籽粒POD活性与面团及面制品褐变程度高度正相关[2-3],高POD活性可以提高小麦面粉及其产品的白度从而提高其商品价值。POD不仅能催化阿魏酸等主要酚酸的氧化,产生发色基团(如醌式结构),使面粉中的无色前体物质形成有色物质,也能氧化降解面粉中的色素类物质(如类胡萝卜素),是面粉和面制品在加工和储藏过程中发生褐变和黄色被漂白的主要原因[4-6]。同时,在面制品加工的面团混合过程中,POD催化所形成的产物在面制品烘烤过程中部分被破坏形成挥发物(如正己醛),增加面制品香气。籽粒中的POD是面包、馒头和面条等面制品白度的重要影响因素[7],与脂肪氧化酶(lipoxygenase, LOX)相比,POD能较少的破坏面制品风味,从而取代LOX作为主要的食品添加剂[8-9]。
环境与基因型均对POD活性有影响,但主要受遗传因素影响[10-11]。前人根据水稻、玉米等禾本科作物基因共线性原理,对普通小麦POD基因进行QTL定位,结果表明,普通小麦POD基因位于第3和第7同源染色体上[12-13]。时 佳等[14]利用90K iSelect 芯片对黄淮麦区151份及北部冬麦区82份小麦品种的籽粒POD基因进行全基因组关联分析(genome-wide association study,GWAS),检测到多个稳定遗传的位点,分别位于2A、2D、5A、6A、7B和7D等染色体上。在检测到的多环境稳定遗传的位点中,2D染色体上的位点(97~100 cM)在3个环境中均能检测到,是比较稳定的遗传位点;7D染色体上位点的贡献率最大,介于18.0%~21.4%之间,并针对7D染色体的POD基因开发了能应用于育种实践的显性互补标记POD-7D1与POD-7D6。POD-7D1 能在具有等位基因TaPod-D1a(与高POD活性相关)的材料中扩增出 540 bp 的片段,而在具有等位基因TaPod-D1b(与低POD活性相关)的材料中无扩增产物;POD-7D6 能在具有等位基因TaPod-D1b的材料中扩增出 640 bp 的片段,而在具有等位基因TaPod-D1a的材料中无扩增产物。魏景欣等[15]利用豆麦/石4185重组自交系(recombinant inbred lines,RIL)群体的214个株系和7 391个SNP标记及一个新开发的STS标记对普通小麦POD活性进行了QTL分析,共检测到3个QTL;并针对3A染色体的POD基因开发了显性互补标记POD-3A1与POD-3A2。POD-3A1能在具有等位基因TaPod-A1a(与低POD活性相关)的材料中扩增出 291 bp 的片段,而在具有等位基因TaPod-A1b(与高POD活性相关)的材料中无扩增条带;POD-3A2能在具有等位基因TaPod-A1b的材料中扩增出 766 bp 的片段,在具有等位基因TaPod-A1a的材料中无扩增条带。时 佳等[14]分别利用TaPod-D1和TaPod-A1位点开发的四对互补显性功能标记,检测两个不同麦区的224份小麦材料的POD活性,并通过对比分析四个不同基因型组合平均POD活性间的显著性差异,发现四个不同基因型组合的品种POD活性高低排列顺序为TaPod-D1a/TaPod-A1b>TaPod-D1b/TaPod-A1b>TaPod-D1a/TaPod-A1a>TaPod-D1b/TaPod-A1a;并且含有两个高POD活性等位基因TaPod-D1a/TaPod-A1b的品种(系)的POD活性显著高于含有单个高POD活性等位基因TaPod-D1a或TaPod-A1b的品种(系)。
尽管目前已有许多和新疆小麦色泽与其他性状关系(如多酚氧化酶、LOX活性等)及调控籽粒色泽有关基因的等位变异检测及其在地方品种的分布规律的相关研究报道[16-20],但迄今为止,对新疆冬小麦品种的POD基因TaPod-D1位点等位变异检测以及两个位点组合基因型检测及分布规律的研究尚未见报道。本研究利用与POD活性相关的功能标记POD-7D1和POD-7D6及POD-3A1和POD-3A2,对98份新疆冬小麦品种进行分子标记检测及分布规律的研究,以期为改良新疆小麦面粉色泽、选育高POD活性品种及分子育种奠定理论和材料基础。
1 材料与方法
1.1 材料及其来源
供试材料为98份新疆冬小麦品种(系),均由新疆农业大学农学院小麦课题组收集保存。其中,地方品种17份,引进品种(系)38份,自育品种(系)43份,这些材料基本反映了不同时期新疆冬小麦的育种现状。
1.2 基因组DNA的提取
为避免因种子混杂而造成错误的检测结果,本研究中每个品种均取3粒大小均匀、籽粒饱满具有较好代表性的种子,提取基因组DNA。
1.3 PCR扩增及检测
1.3.1 检测引物
利用时 佳等[14]开发的显性互补标记POD-7D1和POD-7D6检测小麦7D染色体上TaPod-D1基因的等位变异,利用魏景欣等[15]开发的显性互补标记POD-3A1和POD-3A2检测小麦3A染色体上TaPod-A1基因的等位变异,具体见表1,引物由生工生物工程(上海)股份有限公司合成。

表1 检测POD基因的引物序列、扩增片段长度及其等位变异Table 1 Primer sequences, amplified fragments, and corresponding allelic variations of POD gene
1.3.2 PCR扩增与检测
以小麦基因组DNA为模板,在Eppendorf AG PCR仪(Eppendorf,德国)上进行PCR扩增。PCR体系参考2×Es Taq MasterMix(Dye)说明书配制,除标记POD-3A2外,扩增程序参考2×Es Taq MasterMix(Dye)说明书进行,其中循环数为35,退火温度为64 ℃(标记POD-7D1)或62 ℃(标记POD-7D6)或59 ℃(标记POD-3A1)。标记POD-3A2的扩增程序如下:95 ℃预变性5 min;95 ℃变性30 s,由68 ℃开始,每个循环退火温度均降低0.3 ℃,退火时间30 s,72 ℃延伸1.5 min,共35个循环;最后72 ℃延伸8 min;4 ℃保存。扩增产物用1%的琼脂糖凝胶电泳进行检测,在VILBER LOURMAT凝胶成像系统上拍照保存。
1.4 数据统计与分析
根据每个小麦品种的三粒种子DNA检测结果判断该品种的POD基因TaPod-D1和TaPod-A1位点等位变异类型,选择PCR产物条带清晰、单一且符合目标条带大小的结果进行统计,若结果不一致,需重新提取该品种的其余籽粒DNA,进行检测。
2 结果与分析
2.1 新疆冬小麦品种(系)的 TaPod-D1和 TaPod-A1等位变异类型及分布频率
用显性互补功能标记POD-7D1和POD-7D6及POD-3A1和POD-3A2对98份新疆冬小麦品种(系)进行分子标记检测,结果(图1、图2和表2)表明,在TaPod-D1位点,98份新疆冬小麦品种(系)中,具有TaPod-D1b变异类型的材料有56份(占57.1%),为优势等位变异;在TaPod-A1位点,具有TaPod-A1a变异类型的材料有70份(占71.4%),为优势等位变异。也就是说,在两个位点上,与高POD活性相关的变异类型(TaPOD-D1a和TaPOD-A1b)均为非优势等位变异。
在新疆地方小麦品种、自育小麦品种(系)以及引进小麦品种(系)中,优异等位变异类型TaPod-D1a和TaPod-A1b(与高POD活性相关)的分布频率均低于TaPod-D1b和TaPod-A1a变异类型,但TaPod-D1a变异类型在地方品种、自育品种(系)和引进品种(系)中是渐次升高的,TaPod-A1b变异类型在地方品种、自育品种(系)和引进品种(系)中也有渐次升高的趋势。

a:用标记POD-7D1扩增的结果;b:用标记POD-7D6扩增的结果;M:Marker DL2000;1:邯5316;2:红直头/10;3:奎冬4号;4:新冬29号;5:新冬31号;6:唐山6898;7:奎花2号;8:石冬7号;9:新冬14号;10:奎花1号;11:伊农20;12:新冬23号。
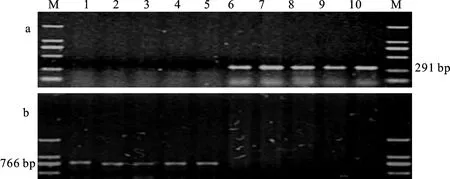
a:用标记POD-3A1扩增的结果;b:用标记POD-3A2扩增的结果;M:Marker DL2000;1:济南4号;2:F49-70;3:小鹅186;4:新冬20号;5:新冬24号;6:碧玛1号;7:碧玛2号;8:工农19;9:新冬21号;10:新冬22号。

表2 新疆冬小麦品种(系)中TaPod-D1和TaPod-A1位点等位基因的分布频率Table 2 Frequencies of TaPod-D1 and TaPod-A1 in different types of winter wheat varieties
2.2 新疆冬小麦品种(系)的 TaPod-D1和 TaPod-A1等位变异组合类型及分布频率
检测结果(表3)表明,在98份新疆冬小麦材料中共出现4种等位变异组合,分别为TaPod-D1a/TaPod-A1a、TaPod-D1a/TaPod-A1b、TaPod-D1b/TaPod-A1a和TaPod-D1b/TaPod-A1b。
在地方品种和自育品种(系)中,优势组合类型均为TaPod-D1b/TaPod-A1a,具有优异组合TaPod-D1a/TaPod-A1b的品种数均最少。在引进品种(系)中,优势组合类型为TaPod-D1a/TaPod-A1a,具有优异组合类型TaPod-D1a/TaPod-A1b的品种数也最少。但优异组合类型TaPod-D1a/TaPod-A1b在地方品种、自育品种(系)和引进品种(系)中有渐次升高的趋势。
在11份具有TaPod-D1a/TaPod-A1b等位变异组合的材料中,6个为自育品种(新冬14号、新冬24号、新冬27号、新冬33号、伊农21和新冬2号),4个为引进品种(F49-70、阿芙乐尔、洛夫林18号和小鹅186),1个为地方品种(麦洛瓦西)。这些品种可以作为高活性POD的优良种质。

表3 新疆冬小麦品种(系)中TaPod-D1和TaPod-A1位点等位变异基因组合类型的分布频率Table 3 Frequencies of different allelelic combination on TaPod-D1 and TaPod-A1 gene loci in different types of winter wheat
3 讨 论
时 佳等[14]利用TaPod-D1位点的显性互补标记POD-7D1与POD-7D6检测243份中国小麦材料,结果表明TaPod-D1a基因型所占比例较低。魏景欣等[15]、耿洪伟等[19]利用TaPod-A1位点的POD-3A1和POD-3A2功能标记检测不同小麦材料,结果显示TaPod-A1b基因型所占比例较低。本研究利用两对功能标记POD-7D1和POD-7D6与POD-3A1和POD-3A2对98份新疆冬小麦品种(系)进行检测,结果表明各位点的等位变异类型的分布与前人的研究结果基本一致,优异等位变异TaPod-D1a和TaPod-A1b类型的分布频率也较低,这可能与新疆乃至全国的育种家在选育优质品种时以面筋强度为主要依据,对酶的特性考虑较少有关;新疆地方小麦品种中的TaPod-D1a和TaPod-A1b的分布频率均为最低,可能与新疆早期育种对面粉及面制品颜色相关的性状研究起步较晚,没有施加足够的选择压力,同时又缺乏对品种(系)POD基因型的认识相关。而新疆自育品种(系)中TaPod-D1a和TaPod-A1b的分布频率介于新疆地方品种和全国品种之间,表明新疆早期小麦育种受地方品种的影响较大,导致高POD活性的TaPod-D1a和TaPod-A1b变异类型较少,近年来随着外引力度加大,使得新疆冬小麦优异等位变异TaPod-D1a/TaPod-A1b得到提升。这一结果与新疆冬小麦早期品种多以地方品种为亲本育成,近期品种则多以自育品种(系)和外引品种为亲本所育的现状相符。表明外来种质的引进改变了新疆小麦的遗传组成,有力地推动了新疆小麦品质的遗传改良。
此外,本研究仅对新疆冬小麦材料中控制POD活性位点7D及3A的等位变异进行了分子标记检测,并未对其POD活性进行测定和关联验证,虽然在一定程度上可通过品种的POD基因型反映新疆冬小麦相应位点不同等位变异的分布和演化规律,但不能排除POD基因型相同而活性差异大的情况。今后,我们将进一步对新疆冬小麦品种(系)的POD活性进行测定,以期通过表型和基因型相结合对新疆小麦的POD活性进行综合评价,为新疆小麦面粉和面制品色泽遗传改良奠定基础。
猜你喜欢
今日农业(2022年4期)2022-06-01
农业灾害研究(2022年1期)2022-05-07
作物学报(2022年6期)2022-04-08
少儿科学周刊·儿童版(2021年21期)2021-12-11
国际医学放射学杂志(2021年5期)2021-10-22
今日农业(2021年4期)2021-06-09
趣味(数学)(2020年4期)2020-07-27
支部建设(2020年15期)2020-07-08
第一财经(2017年36期)2017-09-25
百科知识(2015年18期)2015-09-10
