
复合农林生境游猎型蜘蛛时空动态规律研究
2019-04-12 05:29闫国增周在豹田作宝
绿色科技 2019年6期
杨 钊,闫国增,周在豹,田作宝,王 朔
(1.天津市河西区绿化管理所,天津 300222;2.北京市林业保护站,北京 100029;3.北京市顺义区林业保护站,北京 101300;4.北京大东流苗圃,北京 102211)
1 引言
近年来,我国由农药引发的食品安全问题引起了社会公众对农产品食品安全的高度重视与关注[1~6]。严峻的农产品食品安全局势反映出我国实施与发展绿色农业的必要性和紧迫性。实施绿色农业的一项重要措施是采用非化学手段缓解虫害问题。大量的研究及例证发现[7~9],天敌是调节和控制害虫种群数量的重要因子之一。在各类农田捕食性天敌总数中,蜘蛛的占有比例平均在70%左右,其种类和发生量居各类捕食性天敌之首,是抑制农业虫害发生和发展的重要力量[10~11]。
目前,国外学者针对蜘蛛类捕食性天敌开展了诸多涉及生境与天敌多样性等旨在构建有利于天敌繁衍生息的生境修复研究与尝试[12~15];国内关于蜘蛛类捕食性天敌的研究大都集中在大规模种植的水稻田或棉田[16~19],而农区小规模、多种类作物轮作的种植模式中,蜘蛛种群动态的研究较少,特别是复合农林生境下,林带生境对于蜘蛛种群的生态调控地位与作用鲜有深入的研究报道。
本文围绕华北农区重要的天敌资源——游猎型蜘蛛展开了多角度研究,就其优势种在“农田—林带”环境中的种群时空分布及林带对蜘蛛种群的生态调控作用进行了分析,提出了针对农作物害虫可持续控制的农林复合环境构建模式。
2 研究区域
2.1 试验区域概况
北京市顺义区地处华北平原北部,该地区耕地面积广阔,主要作物种类为冬小麦、玉米以及瓜果蔬菜等。农田周围多种植有防护林带,具有华北平原典型的农业耕作模式和农田景观。
2.2 选取试验样地
本研究的试验地点选在北京市顺义区南彩镇(116°40′E,40°14′N)。试验区域总面积约10 hm2(东西向宽200 m×南北向长500 m),由轮作种植冬小麦、玉米的农作物田(东西向宽200 m×南北向长450 m)和毗邻农田北边的圆柏林(东西向长200 m×南北向宽50 m)组成;本研究从中选取了一具有代表性的林带与农田复合生境块状样地(如图1),总面积约3072 m2(东西向宽32 m×南北向长96 m),样地具体情况如图1。
林带:为样地内北部块状圆柏林,面积约1120 m2(东西向宽32 m×南北向长35 m),林内株行距5 m×5 m,树体平均高度4 m,平均胸径10 cm;林下地被覆盖物较多,有较多的枯枝落叶;植被种类丰富,主要植物有藜(Chenopodiumalbum)、车前草(PlantagoasiaticaL)、马齿苋(Portulacaoleracea)、马唐(Digitariasanguinalis)、狗尾草(Setariaviridis)、苍耳(XanthiumsibiricumPatrinexWidder)、葎草(Humulusscandens)、泥胡菜(Hemisteptalyrata)、鹅观草(Roegneriakamoji)、荠菜(Capsellabursapastoris)、牛筋草(Eleusineindica)等。
农田:为样地内南部的块状田地,位置位于试验区域内农作物田的北部,面积约1920 m2(东西向宽32 m×南北向长60 m),田内(及试验大田内)冬小麦和玉米轮作种植,冬小麦的种植期为每年的10月至翌年的6月上旬,玉米的种植期为每年的6月中下旬至9月底。
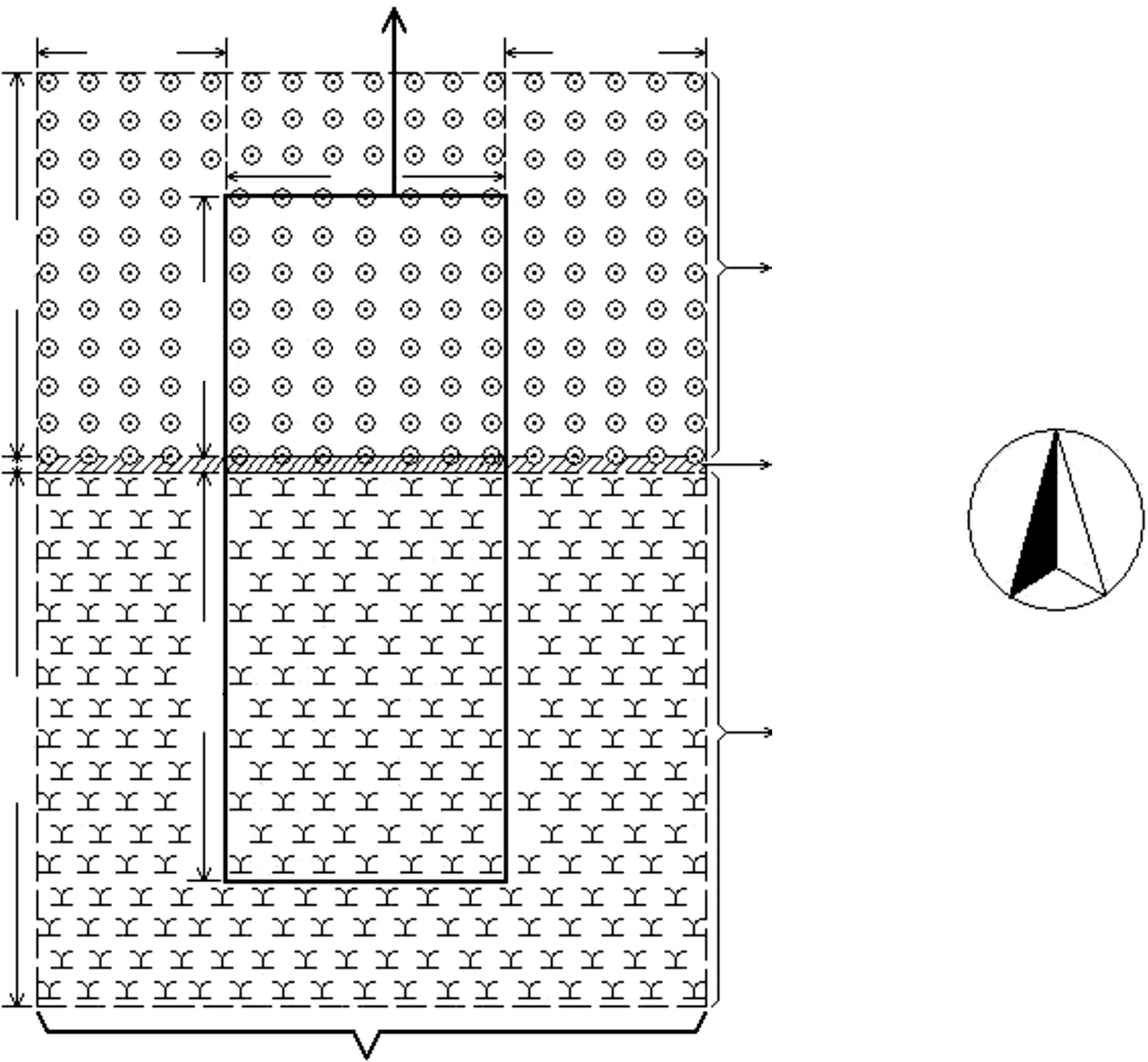
图1 试验区域与样地平面示意
3 研究方法与数据处理
3.1 调查取样方法
调查时间自4月上旬至10月下旬,每10 d调查1次,共调查21次。林带生境采用陷阱法,调查记录生境内样方中地表陷阱内收集到的蜘蛛种类和数量;农田生境采用平行线取样法与陷阱法相结合的方法,记录统计样方内地表陷阱收集到的蜘蛛种类和数量。期间跟踪调查农事活动前、中、后各生境内蜘蛛种群的动态。
3.1.1 陷阱结构
本研究以一次性塑料杯埋入地平面以下作为陷阱诱捕器,塑料杯杯口与地面持平,塑料杯口上方用铁丝固定一个塑料圆盖(阻雨盖)来遮挡雨水(图2A)。塑料杯杯口直径为7 cm,杯底直径5 cm,杯深9 cm,容积约为257 mL。
3.1.2 陷阱溶液
陷阱内溶液为4%的福尔马林和4~6滴50%洗衣粉水组成的混合液。
3.1.3 陷阱样方
陷阱样方由5个单独的陷阱诱捕器按5点排布均匀分布在1m×1m样方内(图2 B)。

图2 陷阱诱捕器与陷阱样方示意
3.1.4 试验样地内样方的分布
林带树间居中位置均匀设置16个1 m×1 m陷阱样方;农田样方设置于距离林带1 m、10 m、20 m、30 m、40 m样线上,每条样线上均匀设置5个1 m×1 m陷阱样方,样方之间间隔6.75 m(图3)。

图3 试验样地内样方分布示意
3.1.5 蜘蛛标本的处理与保存
在野外采集的蜘蛛标本,均使用75%的酒精溶液浸泡,之后带回实验室进行标本整理,分类鉴定到种。
3.2 数据分析方法

4 结果与分析
4.1 样地游猎型蜘蛛多样性
在调查时间段内本研究共调查取样到蜘蛛10科14种(表1)。林带与农田两种生境中游猎型蜘蛛的优势种均为星豹蛛(PardosaastrigeraL. Koch)和白纹舞蛛(AlopecosaalbostriataGrube),且林带生境的种群多样性和均匀度指数均高于农田生境(表2),这反映出林带生境内更为复杂稳定的生态结构与更为丰富的物种。

表1 研究区域内主要蜘蛛种群
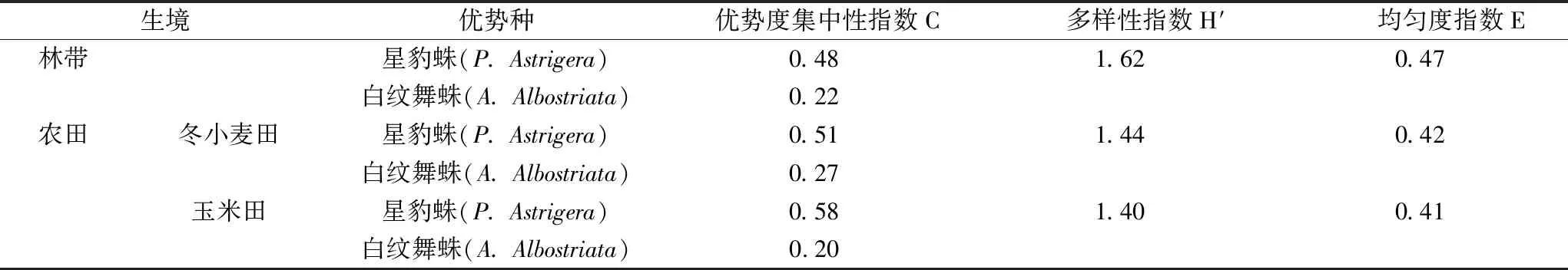
表2 林带与农田两种生境中蜘蛛群落与优势种特征值
4.2 农田游猎型蜘蛛分布规律
本研究调查了自4月上旬至10月下旬农田中游猎型蜘蛛的分布情况,对4~10月内林带与农田中不同距离样线上蜘蛛平均密度的关联性分析(图4)显示:冬小麦—圆柏与玉米—圆柏两类农林系统中,农田不同距离样线上蜘蛛平均密度的变化规律大体相同。冬小麦—圆柏系统中,距林带1m距离样线上蜘蛛种群密度维持在较高的水平范围(453.29±58.06只/100 m2),在背离林带方向1~40 m范围内,蜘蛛密度显著减少(P=0.026,α=0.05),与距林带距离呈负相关;在玉米—圆柏系统中,蜘蛛种群密度在距林带10 m范围内保持高位状态,随后在距林带10~40 m范围内显著减少(P=0.039,α=0.05),并与距林带距离呈负相关。多重比较法(LSD法)进一步显示,在调查时间段内,两类农林系统中1~10m范围内的蜘蛛平均密度均显著高于10m~40 m范围内的蜘蛛平均密度(P冬小麦—圆柏=0.001,P玉米—圆柏=0.000,α=0.01),表明农田蜘蛛的分布集中偏向在近林带范围内。

图4 距林带不同距离样线上蜘蛛密度的变化曲线
4.3 农田施药与蜘蛛种群动态
本研究调查了农事活动中作物喷洒农药前后林带与农田生境内蜘蛛密度的变化,并根据密度与样地面积估算了蜘蛛数量,结果(图5)显示:农田喷洒农药对蜘蛛的杀伤力是巨大的。与喷药前(农田蜘蛛密度为180.23±7.65只/100 m2)相比,喷药后第2 d农田蜘蛛密度急剧下降(20.38±4.84只/100 m2),差异极显著(P农田=0.001,α=0.05);林带生境蜘蛛密度在农田喷药后第2 d亦有所小幅度下降,但并不显著(P林带=0.440,α=0.05)。至喷药后第10 d,林带蜘蛛密度继续下降,农田蜘蛛密度出现回升,两种生境下的蜘蛛密度变化幅度近乎互补(│Δd农田上升│=31.96±3.74只/100 m2)≈│Δd林带下降│=33.37±3.84只/100 m2),对二者进行独立样本t检验,结果(表3)显示:两种生境下蜘蛛密度的变化幅度无显著性差异(F-检验 双样本方差分析:PSig.(1-tailed)=0.492,α=0.05;t-检验 双样本等方差分析:PSig.(2-tailed)=0.817,α=0.05);进一步对二者的变化率(斜率k)进行相关性分析,结果(表4)表明:两种生境下蜘蛛密度的变化率呈极显著负相关(Pearson相关系数:-0.998,PSig.(2-tailed)=0.853,α=0.05)。继续研究至喷药后第20 d林带与农田内蜘蛛密度的变化情况,结果(表5、表6)显示:从喷药后第10 d至喷药后第20 d,两生境中蜘蛛密度的变化近乎拟合(F-检验 双样本方差分析:PSig.(1-tailed)=0.334,α=0.05;t-检验 双样本等方差分析:PSig.(2-tailed)=0.206,α=0.05),二者的密度变化率(斜率k)呈极显著负相关(Pearson相关系数:-0.985,PSig.(2-tailed)=0.776,α=0.05)。这表明在农田施药之后,蜘蛛种群发生了从林带到农田明显的迁移过程。
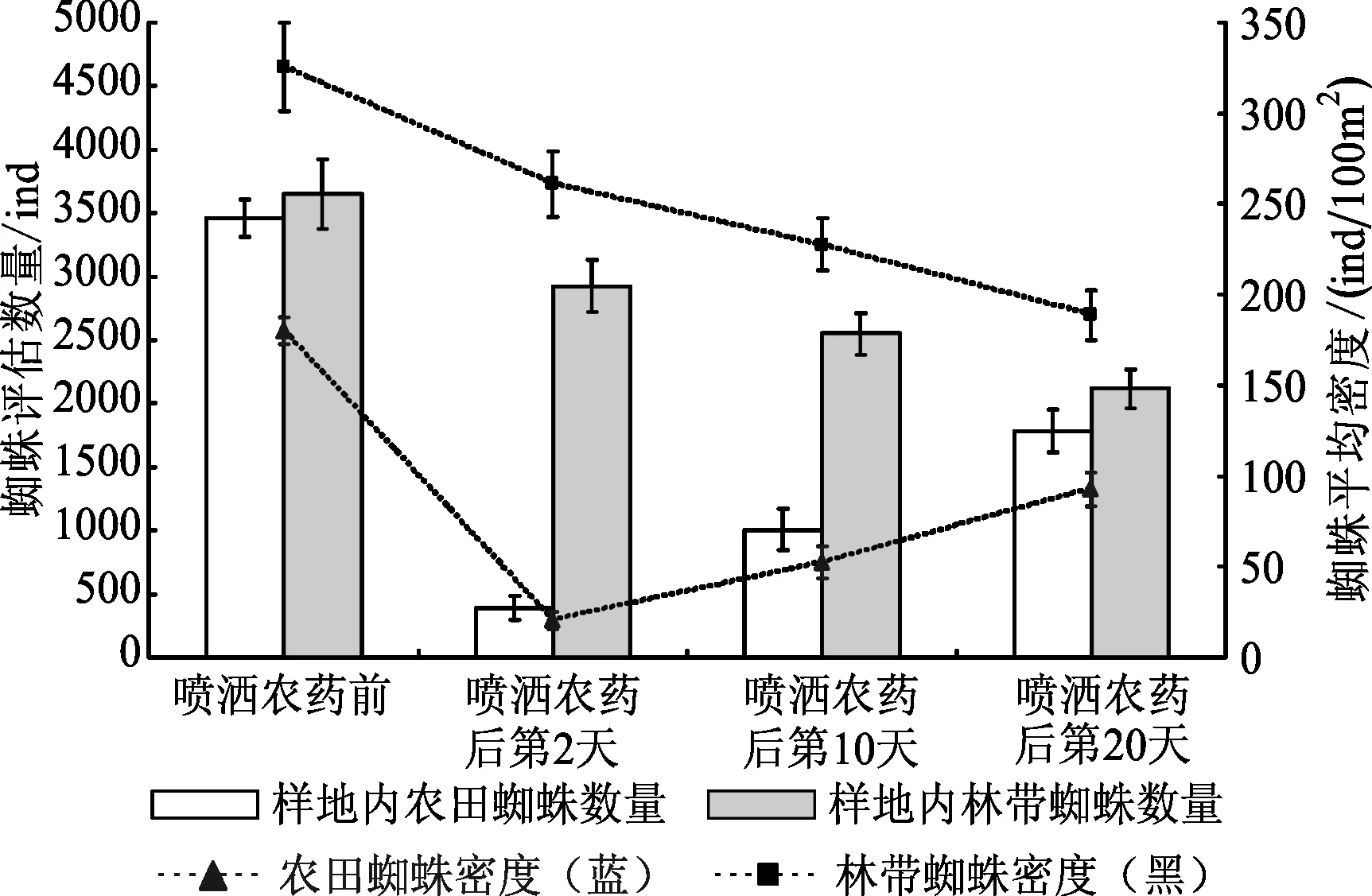
图5 农田喷药前后蜘蛛的分布变化
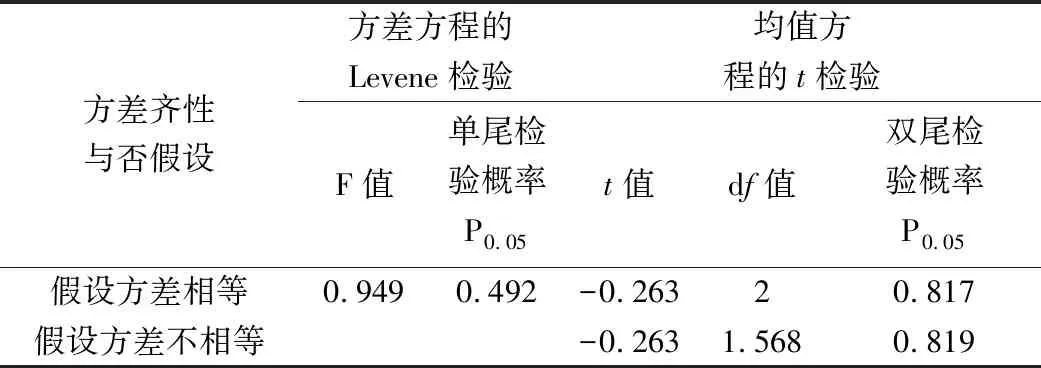
方差齐性与否假设方差方程的Levene检验均值方程的t检验F值单尾检验概率P0.05t值df值双尾检验概率P0.05假设方差相等0.9490.492-0.26320.817假设方差不相等-0.2631.5680.819

表4 喷药后第2 d至喷药后第10 d林带与农田内蜘蛛密度变化率(斜率k)的相关性分析

表5 喷药后第10 d至喷药后第20 d林带与农田内蜘蛛密度变化幅度的独立样本t检验

表6 喷药后第10 d至喷药后第20 d林带与农田内蜘蛛密度变化率(斜率k)的相关性分析
4.4 蜘蛛种群在时间序列上的空间分布
作物的生长期及农事活动影响着作物空间结构、害虫和天敌的种类、数量及其分布。本研究从4~10月对两种优势种蜘蛛在林带与农田内的月平均密度动态进行了调查,结果(图6)显示:在调查时间段内,星豹蛛和白纹舞蛛虽然都为优势种,但前者在林带与农田内的月平均密度明显高于后者(LSD法:P=0.008,α=0.05)。二者的密度月份变动规律相似:在4月份冬小麦拔节期,林带生境和农田生境中的蜘蛛密度差异并不显著;进入5月份春耕农忙期,农田蜘蛛密度显著下降(P星豹蛛农田5月=0.008,P白纹舞蛛农田5月=0.02,α=0.05),林带蜘蛛密度明显上升(P星豹蛛林带5月=0.009,P白纹舞蛛林带5月=0.007,α=0.05),并显著高于农田蜘蛛密度(P5月=0.004,α=0.01);6月中下旬冬小麦的收割期,林带蜘蛛密度与农田蜘蛛密度之间的差异继续扩大(P6月=0.001,α=0.01);7、8月份为玉米的生长期,此前偏重于林带生境分布的蜘蛛密度,在这个时期与农田生境蜘蛛密度的差异水平逐步减小,进入8月份后农田蜘蛛密度反超林带;9月份玉米进入成熟收割期,受农事活动与生物节律的影响,农田蜘蛛密度再次明显下降(P星豹蛛农田9月=0.006,P白纹舞蛛农田9月=0.008,α=0.05),林带蜘蛛密度再次显著上升(P星豹蛛林带9月=0.001,P白纹舞蛛林带9月=0.015,α=0.05),并再次显著高于农田蜘蛛密度(P9月=0.003,α=0.01);10月份农田作物秋收殆尽,游猎蜘蛛即将进入越冬期,林带蜘蛛密度继续上升,农田蜘蛛密度进一步下降,二者之间的差异再次更加显著(P10月=0.000,α=0.01)

图6 林带和农田生境中星豹蛛与白纹舞蛛种群密度变化曲线
5 结语
华北农区农作物的轮作种植及作物季节性的生长,在不断地改变农田景观的同时,也深刻的影响着农田及其周边生境内游猎型蜘蛛的种群及其时空分布。在调查时间段内,园蛛科、卷叶蛛科、肖蛸蛛科和漏斗蛛科的蜘蛛大多在农田区域内被采集到,这类蜘蛛因在作物枝叶间结网而具有农田蜘蛛属性[23];球蛛科的蜘蛛则多出现在林带区域内,或与该类蜘蛛偏好在森林草丛、田间杂草地以及山坡灌木丛等非作物生境栖息觅食有关[23];狼蛛科、跳蛛科、蟹蛛科以及管巢蛛科的蜘蛛由于具有出色的游猎活动能力[23],在农田与林带两种生境内均有被采集到,其中狼蛛科的星豹蛛和白纹舞蛛为研究区域内最为壮大的蜘蛛种群。
农田环境的多变性与不稳定性造就了游猎型蜘蛛出色的应变与适应能力,而林带则以其稳定性甚至唯一性为游猎型蜘蛛提供了重要的越冬、栖息与避难场所。研究显示,在作物种植前与收割后农田环境空旷,农田中游猎型蜘蛛数量显著减少,而林带中游猎型蜘蛛数量显著增加(P6月=0.001,P10月=0.000,α=0.01),林带由此成为游猎型蜘蛛在进入农田前与作物收割后主要的栖息、觅食与越冬场所。
在作物生长期内,频繁的农事活动对农田游猎蜘蛛的种群规模产生了较大的压力,特别是喷洒农药给游猎型蜘蛛种群带来了极大的负面影响(P=0.001,α=0.05),而此时林带又成为游猎型蜘蛛躲避农事活动干扰的天然理想庇护所。林带生境内植被种类丰富,生态结构复杂,食物资源相对稳定,不但能够有效的缓冲喷洒农药等农事活动给游猎型蜘蛛种群所带来的负面杀伤力,其生境内的游猎型蜘蛛又可成为未来农田游猎蜘蛛种群重建的重要来源[24~26]。当农田环境再次趋于稳定,作物害虫逐渐成为新的优质食源选择时,游猎型蜘蛛便发生了从林带到农田明显的迁移过程(林带与农田内蜘蛛密度变化t-检验:PSig.(2-tailed)=0.817,α=0.05;二者蜘蛛密度变化率的Pearson相关系数:-0.998,PSig.(2-tailed)=0.853,α=0.05)。
林带对于游猎型蜘蛛的分布具有积极作用,本研究显示,农田生境内的游猎型蜘蛛多选择在近林带范围内聚集,表现出对林带明显的“依托”关系,其种群密度在背离林带40 m范围内显著下降(P冬小麦—圆柏=0.026,P玉米—圆柏=0.039,α=0.05),呈现出明显的“林带效应”。在复合农林系统中,非作物生境对维持天敌种类和数量发挥着重要的作用与功能[27,28],林带生境内复杂稳定的生态结构以及丰富多样的食物来源,为游猎型蜘蛛种群的发生、增长、繁衍、生息提供了适宜的生存环境,并对农田游猎型蜘蛛的分布具有积极的影响效果,其对于维系农田游猎蜘蛛群落的延续以及生态链的稳定发挥着重要的生态调控功能。
综上所述,复合农林生境中游猎型蜘蛛的时空动态分布受周边生境的影响较为深刻,农田游猎蜘蛛在分布规律上呈现出明显的“林带效应”;在作物栽种前与收割后林带是游猎型蜘蛛种群延续的重要种库[24,25],在农事活动时则是游猎型蜘蛛的重要避难场所,在农田天敌种群过渡和重建中起到关键的桥梁作用[29]。因此,有序营建农田林带,对于农区天敌的有效保护和利用具有积极意义,是实现农区作物害虫可持续防治的一个科学措施。
猜你喜欢
东北林业大学学报(2022年1期)2022-03-28
种子(2022年2期)2022-03-21
安顺学院学报(2021年4期)2021-09-16
落叶果树(2021年6期)2021-02-12
新农业(2019年6期)2019-07-08
农家科技下旬刊(2017年3期)2017-04-26
中国科技纵横(2016年15期)2016-12-29
体育科技(2016年2期)2016-02-28
现代园艺·综合版(2012年16期)2012-07-09
中国青年(1966年5期)1966-08-17
