
广西会仙湿地不同植物根际细菌群落结构及多样性研究
2019-04-09 05:57凃月李海翔姜磊董堃王敦球
生态环境学报 2019年2期
凃月,李海翔,姜磊,董堃,王敦球
桂林理工大学环境科学与工程学院,广西 桂林 541006
岩溶湿地指主要分布在岩溶地区(包括地表、地下),或以岩溶水为主要补给水源,具有岩溶地区特有的富钙偏碱性水土特征和典型岩溶水土循环演化机制,以喜钙耐碱的湿地生物群落为主或与喜钙耐碱的生物群落相互依存为特征的内陆湿地,包括岩溶湿地的地表或地下的湖泊、沼泽、河流或其他岩溶地下水文系统,但其规模小、数量少、稳定性差、破坏后难以恢复(蔡德所,2012)13。广西会仙湿地作为低中海拔中规模最大的岩溶湿地,其自然资源丰富,是岩溶地区少见的物种库,被称作漓江流域的“肾脏”,对涵养水源、气候和环境改善、水质净化、生物多样性保护以及漓江蓄洪补水等方面发挥重要的生态作用(Ghermandi et al.,2014)。由于周边污水排放、河道占用,会仙湿地逐渐萎缩,近40年来湿地面积减少了27 km2,生态功能严重退化,使生态系统受到严重威胁(林栋材等,2008)。
一般认为,湿地系统的净化功能是利用植物-土壤-微生物的综合作用实现的,水生植物通过与根际微生物的相互作用完成污水净化,其中微生物是对污染物进行吸附和降解的主要生物群体和承担者(Liang et al.,2003;Liang et al.,2017)。研究(Münch et al.,2007)发现,植物根际微生物在污水净化过程中比湿地植物发挥的作用更大。越来越多的研究表明,在湿地系统中微生物种群的多样性、稳定性对维护湿地生态系统的平衡具有关键作用(De-Bashan et al.,2004;武钰坤等,2012)。不同植物根际泌氧能力和根际分泌物性质不同,在一定程度上影响微生物的种类和多样性,而微生物的活性、多样性与污水处理过程紧密相关,表现为根际微生物群落结构多样性越高,湿地系统的净化效果和稳定性越好(魏成等,2008;Zhou et al.,2015)。细菌是微生物的重要组成部分,对不同根系分泌物的利用率和敏感度远高于其他微生物(真菌、放线菌),能够与植物根系并存(李潞滨等,2008),在根际微生物中占主导地位。高通量测序技术从整体水平上研究微生物种群结构,真实、准确地反映环境中细菌群落的多样性和复杂性,具有低成本、高通量、流程自动化等优势,被广泛运用于植物根际细菌群落结构特征研究中(李庆岗等,2012;Ligi et al.,2014;Cao et al.,2017;吾尔恩·阿合别尔迪等,2017)。
目前,大部分研究主要集中在沉积物和土壤细菌群落结构等方面,而对于不同植物根际细菌在低温下多样性和丰富度研究较少。低温条件下,植物地上部分枯萎甚至死亡,使得微生物发挥重要降解作用,而微生物的数量、多样性和群落结构在很大程度上影响污染物的去除。本研究以会仙湿地中分布较广的单一优势植物群落为研究对象,采用Illumina HiSeq 2500高通量测序技术,研究南方低温条件下根际土壤细菌群落组成和变化,以丰富喀斯特湿地植物根际细菌多样性,为低温环境下湿地生态系统的结构和功能研究提供依据。
1 材料与方法
1.1 样品采集和前处理
会 仙 岩 溶 湿 地 ( 25°01′30″- 25°11′15″N ,110°08′15″-110°18′00″E,海拔 118 m)气候属于中亚热带季风气候,全年月平均温度为8.6-28.9 ℃,年平均降雨量为 1890.4 mm,主要集中在 4月、5月,植物群落类型以挺水植物群落(60.45%)和沉水植物群落(9.7%)为主,植物种类较多,且生长茂盛,盖度常可达 80%-95%,以单一优势种群较多,主要群落有芦苇、华克拉莎、长苞香蒲、苦草、五刺金鱼藻等(蔡德所,2012)21,24,133,139-143。研究表明(Gopal,1999),选择当地优势植物,突出生物多样性,是提高湿地净化能力和保护的关键措施。故以会仙湿地中典型的单一优势芦苇、苦草、华克拉莎和美人蕉植物群落为研究对象,于 2017年 10月10日(秋,水温18-23 ℃)和2017年12月27日(冬,水温7-11 ℃)进行采样(图1),采样点选择在该河段植物生长优势且种群单一、覆盖度高的位置。每次采样共15个,为了减少误差,有3组重复样,取水样后于同一位置取植物根际土样。采集水样时,先润洗1遍采样瓶,然后连同瓶盖一同放在水面10 cm下,打开瓶盖避免水中悬浮物进入瓶中,采集完后确保瓶中无气泡,送回实验室尽快测定水质指标。植物根际土壤取样时,采用“抖根法”,先将整个植物根际部分挖出,抖掉与植物根际松散结合的土壤,放入无菌封口袋中,一部分冷冻保存在冰盒里,带回实验室于无菌操作台上将根系0-4 mm 范围的土壤放入无菌离心管中,-80 ℃保存,用于测定植物根际土壤细菌种群结构(刘波等,2010);另一部分土样自然风干,磨细过60目和100目筛,用于土样理化指标的测定;底泥取样时,使用底泥采样器采集0-10 cm的土壤作为底泥样。

图1 采样点位置Fig. 1 Sampling location
测定指标与方法:水质总氮测定方法为碱性过硫酸钾消解紫外分光光度法,总磷测定方法为钼酸铵分光光度法。根际土壤总磷测定方法为碱熔-钼锑抗分光光度法,全氮测定方法为凯氏法,有机碳测定方法为重铬酸钾氧化-分光光度法,pH测定方法为电位法。
1.2 植物根际土壤DNA的提取和PCR扩增
采用CTAB法对样本的基因组DNA进行提取,然后通过1%的琼脂糖凝胶电泳检测DNA的质量和浓度,用无菌水稀释样品浓度至1 ng·μL-1。以稀释后的基因组DNA为模板,采用细菌16S rDNA基因 V4+V5 扩增区域,引物 515F(5′-GTGCCAGCMG CCGCGGTAA-3′)/907R(5′-CCGTCAATTCCTTTG AGTTT-3′)。反应条件是:98 ℃预变性 1 min;98 ℃变性10 s,50 ℃退火0.5 min,72 ℃延伸0.5 min,循环30次;然后72 ℃延伸5 min。使用2%的琼脂糖凝胶电泳检测扩增产物。扩增过程使用高效、高保真酶以保证扩增过程的准确性(DNA提取、PCR扩增、测序由北京诺禾致源科技股份有限公司完成)。
1.3 文库及系统发育
PCR扩增产物经过纯化回收后,使用 TruSeq®DNA PCR-Free Sample Preparation Kit试剂盒进行文库构建,然后采用Illumina HiSeq 2500进行高通量测序。所得序列去除 Barcode和引物序列后,使用FLASH(Magoc et al.,2011)和 MUSCLE3.8.31(Edgar,2004)对样品进行拼接、过滤、多序列对比。
1.4 数据分析
利用Uparse软件(Edgar,2013),在97%相似性下,对样品有效数据进行 OTU序列聚类。采用Mothur法(阈值0.8-1.0)和SILVA(Wang et al.,2007)的SSUrRNA数据库(Quast et al.,2013)对OTU代表序列进行物种注释分析,得到样品在各分类水平上的群落组成。使用Qiime 1.9.1计算测序深度 ACE指数、丰富度指数(Chao1)、多样性指数(Shannon)。不同分类水平上物种组成柱状图用Origin.Pro 9.0绘制,可以直观了解群落组成情况。数据分析于SPSS 19.0中进行,采样图用Arcgis 10.2绘制。
2 结果与分析
2.1 采样点水质及土壤的理化性质
湿地中污染物来源主要是生活废水的排放,导致水中N、P含量偏高。由表1可知,土壤pH为6.95-7.86,水质 DO 为 4.13-12.90 mg·L-1。不同植物间的理化指标差异显著(P<0.05),同种植物根际土壤的理化指标呈现相同的变化趋势,例如,冬季各植物根际土壤中TN、TP、TOC大多高于秋季。水质TN平均质量浓度表现为冬季(2.27 mg·L-1)高于秋季(2.09mg·L-1),超出地表水Ⅴ类标准(GB 3838—2002);而水质 TP 表现为秋季(0.19 mg·L-1)高于冬季(0.16 mg·L-1),超过Ⅱ类标准。沿着水流方向,水中N、P有逐渐降低的趋势,说明湿地系统对N、P有净化作用。
2.2 群落结构分析
2.2.1 在门分类水平上
采用 97%相似水平作为标准界定 OTU(张健等,2010),对OTU代表序列进行聚类和物种注释,共检测出细菌有65门、160纲、219目、391科、677属和 246种,细菌群落具有较高的多样性。由图 2可知,所检出的细菌由10个主要门类组成,包括变形菌门(Proteobacteria)、绿弯菌门(Chloroflexi)、酸杆菌门(Acidobacteria)、浮霉菌门(Planctomycetes)、拟杆菌门(Bacteroidetes)、硝化螺旋菌门(Nitrospirae)、蓝藻门(Cyanobacteria)、放线菌门(Actinobacteria)、厚壁菌门(Firmicutes)和迷踪菌门(Nitrospinae)等。在秋、冬两季不同植物根际土壤中,变形菌均为第一优势类群,所占范围是18.6%-49.5%,主要包括4个亚类:γ-变形菌( Gammaproteobacteria ) 、 β- 变 形 菌(Betaproteobacteria)、δ-变形菌(Deltaproteobacteria)、α-变形菌(Alphaproteobacteria),其所占的比例范围分别为5.4%-12.9%、1.9%-19.2%、1.8%-18.1%、1.9%-8.8%,其中,以β-变形菌和δ-变形菌为优势菌纲。绿弯菌门占 2.8%-21.6%,为群落中第二大优势类群,且大部分属于厌氧绳菌纲(Anaerolineae)。酸杆菌门占 1.8%-12.4%,为群落中第三大优势类群,其主要是由全噬菌纲(Holophagae)和Subgroup_6组成。相比而言,秋季美人蕉中主要的细菌群落有较大区别,主要是由厚壁菌门(29.0%)、放线菌门(26.1%)、变形菌门(18.6%)以及蓝藻菌门(15.0%)组成,且大部分属于杆菌纲。

表1 采样点水质及植物根际土壤理化性质Table 1 Physicochemical parameters in water and plant rhizosphere soil

图2 门分类水平上细菌群落组成Fig. 2 Classification of bacterial sequencing at level of phylum
从季节变化上看,占优势的群落无较大区别,还是以变形菌门、绿弯菌门为主,但在组成数量上有所区别。变形菌在冬季所占的平均比例(45.1%)高于秋季(38.0%),其中美人蕉中变形菌组成差异最大,秋季为18.6%,冬季为48.7%;而绿弯菌门在华克拉莎中区别较大,秋冬季分别为 17.2%、11.9%。同一植物不同季节有新微生物出现,但其所占比例很小,如,冬季美人蕉中出现的深古菌门(Bathyarchaeota、0.3%)、Parcubacteria门(0.03%)等在秋季美人蕉中均未检出。柔膜菌门(Tenericutes)仅不存在于美人蕉根际环境,而在其他样中均可检测到。与植物根际土壤相比,秋、冬底泥中主要门类细菌群落组成差异最小,说明植物根际对环境中微生物群落分布有影响。总之,不同植物根际土壤细菌中占主导的种群无较大差别,但在数量上有区别。
2.2.2 在属分类水平上
为了解属水平物种进化关系,将丰度前100属的代表序列构建系统发育树(图3)。结果表明,组成前100属的菌有15门,主要是变形菌(51.0%)、放线菌(12.0%)、厚壁菌(7.0%)。其中,丰度较高且存在于大多数样品中的有硫杆菌属(Thiobacillus)、厌氧粘菌属(Anaeromyxobacter)、H16属、脱硫酸盐橡菌属(Desulfatiglans)、地杆菌属(Geobacter)、Candididatus_competibacter属(聚磷微生物)、Syntrophorhabdus属、芽孢杆菌属(Bacillus)等。从图可知,芽孢杆菌属(占所有属的 28.0%)、红球菌属(Rhodococcus,18.0%)、unidentified_Chloroplast属(15.0%)在秋季美人蕉中的丰度显著高于其他植物,冬季美人蕉中占优势菌没有秋季明显,属水平上分布较均匀,占总属的0.01%-1.2%。不同植物(除秋季美人蕉)共有的优势菌属为芽孢杆菌属(0.3%-28.0%),其次为厌氧粘菌属(0.8%-2.2%)。总之,除秋季美人蕉中优势菌群明显外,其他9个样各菌在属水平丰度较低,分布较均匀,秋季美人蕉中Unidentified bacteria占20.2%,说明该植物根系土壤中还存在大量未知细菌,有待进一步研究。
2.3 细菌群落多样性和环境因子相关性
运用 ACE、Chao1、Shannon指数对不同植物根际土壤细菌多样性进行分析(表2)。结果表明,各样本中物种覆盖率在95.4%-97.8%之间,能够反映植物根际细菌群落的真实情况。通过分析发现,不同植物、不同季节根际土壤细菌多样性存在显著差异(P<0.05),综合而言,冬季苦草指数最高、秋季美人蕉最低,表明冬季苦草细菌丰富度和多样性最高,秋季美人蕉的最低。从季节上看,冬季ACE、Chao1、Shannon指数分别为5892、6614.3、10.1;秋季各指数为4711.8、4637、8.68,冬季平均指数高于秋季,说明冬季土壤细菌的多样性和丰富度高于秋季。从植物种类上看,冬季Shannon指数以苦草最大,为 10.33,其次为美人蕉和芦苇,华克拉莎最低,为9.83;秋季则不同,华克拉莎、芦苇指数较高,分别为10.04和10.17,美人蕉最低为5.13。总之,不同植物根际土壤细菌群落分布存在差异,根际细菌多样性随季节而变化,且群落变化规律有差异。

图3 前100属代表序列系统发育树状图Fig. 3 Phylogenetic tree of top 100 genus represent sequence

表2 秋、冬植物根际土壤中细菌的丰富度和多样性指数Table 2 Diversity index of bacteria in plants rhizosphere soil of autumn and winter
为了解环境因子对细菌群落结构的影响,对环境因子和多样性指数进行 Spearman相关分析(表3),结果表明,根际土壤中TP、TOC、TN与覆盖率呈负相关,与Ace、Chao1、Shannon指数呈正相关,水质TP与多样性指数关系则表现相反。运用典范对应分析(图4)和Spearman分析(图5),进一步筛选影响细菌群落的驱动因子,由图可知,前两轴共解释了 95.25%的变异,第一主轴上的主要影响因子是土壤TOC、TN、TP,其相关系数分别是0.84、0.74、0.54,第二主轴上的影响因子是水质TP、TN、DO,其相关系数分别是0.71、0.61、0.66。其中水质 TP、TN、DO等对底泥中细菌群落有较大影响,土壤 TOC、TN、TN对植物根际土壤细菌群落有较大影响;而秋季美人蕉不同,主要受水质TP、TN、土壤pH的影响。土壤TOC、TN与变形菌门、迷踪菌门丰度呈显著正相关,与厚壁菌门呈显著负相关,与蓝藻菌门、放线菌门相关性不明显;水质TP、TN、土壤pH与厚壁菌门呈极显著正相关,与硝化螺旋菌门、酸杆菌门、浮游菌门呈显著负相关,与绿弯菌门、拟杆菌门的相关性不大。

表3 环境因子与多样性指数的Spearman相关分析Table3 Spearman correlation analysis of environmental factors and diversity index

图4 样品与环境因子的CCA分析Fig. 4 CCA analysis between samples and environmental factors
2.4 细菌群落相似性和差异性分析
根据OTU聚类结果对样品进行均一化处理后,绘制Venn图(图6),比较不同季节、不同植物细菌群落的相似性,10个样品共有的OTU数为935,占总OTU数(46844)的2.0%。从季节上看,秋季共有OTU为1042(4.8%),而冬季为1182个(4.7%),这表明季节对植物根际土壤细菌的种类和数量有影响,但占主要比重的细菌种类几乎无变化,还是以变形菌、绿弯菌为主。从种类上看,秋、冬季华克拉莎相似性最高,为35.5%,说明华克拉莎细菌群落季节变化最小;芦苇次之,相似性为34.4%;然后是苦草为31.9%;美人蕉细菌群落季节变化最大,相似性最低为21.3%,对环境的响应更敏感。两季底泥相似性最高(36.7%),说明底泥中细菌群落受温度变化较小以及植物种类对细菌群落存在一定影响。聚类结果(图 7)显示,不同植物根际细菌群落之间有差异,分为4个分支,以同一个季节聚类在一起,说明温度对植物根际土壤细菌群落的分布有一定影响。其中秋季美人蕉独立成一个分支,主要原因是其土壤中营养物质含量明显低于其他土样,导致群落分布不同。

图6 OTU venn分析Fig. 6 Analysis of OTU venn
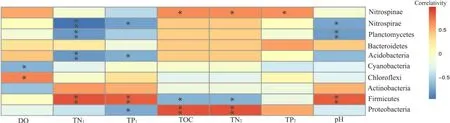
图5 前10门细菌与环境因子相关分析Fig. 5 Correlation analysis between the top 10 bacteria and environmental factors

图7 10门主要细菌聚类分析Fig. 7 Cluster analysis of 10 dominant bacteria
3 讨论
3.1 细菌群落结构
本研究区植物根际土壤中细菌群落具有较高的多样性,发现 10门类主要细菌:变形菌门、绿弯菌门、酸杆菌门、浮霉菌门、拟杆菌门、硝化螺旋菌门等,说明这些菌类分布范围广,并以变形菌门、绿弯菌门、酸杆菌门为主,三者约占总测序序列的57.0%-83.5%。而阮楚晋等(2018)对会仙湿地底泥可培养原核微生物多样性的研究中,仅分离得到厚壁菌门、变形菌门、放线菌门、拟杆菌门这4门细菌,这说明采用现代分子学技术更能灵敏和准确地揭示细菌群落结构多样性。
会仙湿地不同植物根际土壤中优势菌均以变形菌、酸杆菌、绿弯菌为主,这一结果与国内外有关湿地土壤中微生物多样性的研究结果一致(房昀昊等,2018)。刘兴等(2015)对贵州喀斯特地区植被土壤群落结构进行研究,共发现 33门细菌,其优势菌群与本文一致,但发现的细菌种类比会仙喀斯特湿地的低,说明会仙湿地中细菌种类丰富,对维持湿地系统的平衡有重要作用。研究发现,变形菌在会仙湿地秋、冬植物根际土壤中均为优势菌,其中以 β-、δ-、γ-变形菌纲为优势亚群,具有丰富的代谢多样性,在 N、P、C等元素循环及其他污染物的降解过程中起关键作用(Hoefel et al.,2005;Ansola et al.,2014);通过分析发现秋季美人蕉中变形菌最低,水中N、P含量最高,而其他植物中变形菌较高,环境中N、P较低,同样证明变形菌有较好的脱氮除磷作用。苦草中变形菌含量较高,表明苦草在脱氮除磷方面可能发挥重要作用。绿弯菌作为第二优势菌,主要分布在有机物丰富的沉积物中,通常参与COD和萘的去除(Grégoire et al.,2011;杨小丽等,2013)。酸杆菌主要存在于湿地、活性淤泥中,能够参与枯枝烂叶分解的碳循环(Chen et al.,2015),在芦苇和华克拉莎中丰富度较高,说明芦苇和华克拉莎的根际环境有利于碳的循环转化。与其他植物相比,秋季美人蕉根际细菌群落结构明显不同,以放线菌、厚壁菌和蓝藻菌为主,而柔膜菌却仅存在于其他植物根际土壤中,主要原因是土壤理化性质、植物群落类型对土壤细菌群落结构有较大影响(Zhao et al.,2010)。除此以外,秋季美人蕉根系土壤中酸杆菌、硝化螺旋菌明显低于其他样,可能原因是酸杆菌为嗜酸菌,而植物根际土壤pH值以秋季美人蕉最高(7.63),不利于其生长,同样说明酸杆菌可以在非酸性条件下生存。硝化螺旋菌在好氧条件下可以将亚硝酸盐氧化成硝酸盐,在氮循环中起着重要作用,而美人蕉根际环境DO最低为4.13,抑制硝化螺旋菌的繁殖,导致硝化螺旋菌丰度最低;浮霉菌作为兼性厌氧菌和严格厌氧自养菌,在秋季美人蕉中最低,仅有0.9%,说明除DO外,还有其他因素影响植物根际土壤中细菌分布。
地杆菌属、厌氧粘菌属、硫杆菌属、H16属等在大多数植物根际土壤中皆存在,属于革兰氏阴性菌,可能是湿地植物根际环境有利于革兰氏阴性菌生长,亦有利于环境中污染物的降解。研究表明(周文杰等,2015),假单胞菌属(Pseudomonas)是根际有益微生物,对植物的生长有显著的促进作用,但其对环境的适应能力较差,相比于其他根际微生物竞争较弱,营养物浓度是其主要影响因素。本研究得到了类似结果,相比于其他植物而言,美人蕉生长环境中的营养物浓度有明显差异,从而导致其假单胞菌属丰度最高。海源菌属(Idiomarina)、嗜冷杆菌属(Psychrobacter)只在冬季出现,可能原因是不同细菌对温度等条件要求不同,有些细菌却适宜在冬季环境下生存。韦荣球菌属(Veillonella)只存在底泥中,说明植物根际对细菌群落分布有影响(Chaudhary et al.,2012)。此外,在植物根际环境中还发现了有益微生物(王建宁等,2012;林莹莹等,2016),如红球菌属和芽孢杆菌属,它对碳源的利用率强,在促进植物生长和污染物降解中起重要作用,在秋季美人蕉中含量最高,可能是环境中碳源丰富促进其繁殖和秋季温度更适应红球菌的生长。
综上所述,不同植物根际土壤中细菌群落结构有一定差别,但差别不大,占主导的细菌群落相同,相同植物根际土壤中细菌群落结构相似度较高(商迎迎,2017),营养物浓度对细菌分布有较大影响。
3.2 细菌群落结构多样性和环境因子
秋、冬植物根际土壤细菌多样性指数均较大,说明供微生物生长的营养物质较丰富(雷旭等,2015)。总体而言,冬季各植物根际多样性高于秋季,主要是因为冬季土壤中营养物浓度高于秋季,除此以外,会仙湿地植物种类丰富、面积大,植物根际细菌对秋、冬季温度变化具有较好的适应能力。综合各指数可知,植物根际土壤细菌多样性表现为芦苇>华克拉莎>苦草>美人蕉,与赵庆节(2011)研究相似。本研究发现,芦苇根际土壤细菌多样性和丰富度最高,原因可能在低温情况下,芦苇根际的好氧菌仍保持较高比例以及芦苇根系较其他植株更发达,有利于细菌的繁殖。值得注意的是,由于美人蕉是人工引入到湿地中的,在短时间内植物根际吸收作用不明显,使得秋季美人蕉根际土壤中各指标(表 1)远低于其他植物根际环境中的,故其细菌多样性、丰富度远低于其他样。与植物相比,底泥中各多样性指数最大,说明植物根际对某些细菌产生抑制作用,Zhou et al.(2009)研究表明,湿地表层中细菌的数量显著高于其下层的数量,并且出现明显的分层效应,形成好氧和厌氧层交替现象,种植不同植物对细菌群落分布的影响主要发生在0-10 cm处。
已有研究证明(Córdova-Kreylos et al.,2006;金笑等,2017),土壤TN、TOC是细菌群落结构影响的显著因子,与微生物群落数量、生物量等呈显著正相关。通过对环境因子与样品的 CCA分析发现,影响植物根际土壤细菌群落结构的主要环境因子同样是土壤 TOC、TN,其与变形菌门、迷踪菌门呈显著正相关,与厚壁菌门呈显著负相关,而水质环境因子主要对底泥中细菌群落分布有影响。所有样品分成3个相对独立的群,其中秋季美人蕉独立成一个群,其细菌群落结构与其他样本有明显差异,影响其细菌群落的主要环境因子也不同,可能原因是在土壤营养物较低的情况下,水中含量较高的营养物对其产生了主要影响。大多数不同植物形成一个群(图4),对环境因子的响应趋势相似,可能是不同植物对细菌群落结构有影响,但是没有营养物浓度对细菌群落影响大。国内外相关研究表明(Buée et al.,2010),在土壤细菌群落研究中,pH作为指示因子之一,对细菌群落的分布有较大影响。本次研究同样证明,pH对细菌群落有显著影响,不同细菌对pH的响应不同,厚壁菌门丰度与pH呈极显著正相关,硝化螺旋菌门、酸杆菌门、浮游菌门丰度与pH呈负相关,研究结果与刘兴等(2015)相似。
3.3 细菌群落结构相似性和差异性
秋、冬植物根际土壤细菌群落相似性分析表明,不同植物根际细菌群落具有一定的相似性,但相似性较低,说明湿地土壤环境中形成了某些稳定的细菌群落,这些细菌适应湿地环境的变化,对底泥和植物根际环境中细菌群落结构有重要作用。秋季样的整体相似性略高于冬季,说明冬季各植物细菌群落随季节变化更大,主要原因可能是冬季植被生长、耐寒能力不同,各植物根际活动差异较大。美人蕉根际细菌群落结构在秋、冬两季相似性最低,说明其受季节变化影响最大,其次是苦草和芦苇,华克拉莎变化最小。原因可能是美人蕉相对于其他植物根际范围小,没有生长优势,根际在秋、冬两季活动差异相对更高,而芦苇、华克拉莎在秋、冬两季都为优势植物,所以更为稳定。底泥细菌群落的相似性最高,为36.7%,说明湿地土壤细菌群落结构受到植物根际活动的影响,但并不能改变组成土壤细菌群落的主要种类,因为这是由温度、植物种类、污水和土壤性质共同决定的。
4 结论
会仙湿地中植物根际土壤细菌有较高的多样性,发现10门主要细菌,变形菌为第一优势菌群,所占比例为42.4%;优势菌纲占比最高的菌群为β-变形菌(13.5%);优势菌属占比最高的菌群为芽孢杆菌属(3.8%)。其中,秋季美人蕉根际中细菌种类与其他样本明显不同,主要是由于其生长土壤环境中营养物浓度低导致。从季节上看,秋、冬植物根际土壤细菌多样性和丰富度具有差异,冬季高于秋季;从植物种类上看,芦苇多样性和丰富度最高,其次为华克拉莎和苦草,美人蕉最低。相似性分析表明,同一植物不同季节其细菌群落结构相似性较高,美人蕉细菌群落结构随季节变化最大,其次为苦草和芦苇,华克拉莎变化最小。植物根际土壤细菌群落结构和多样性受环境因子的影响较大,其中土壤TOC、TN对细菌群落影响最为显著。从植物根际土壤微生物多样性、丰富度、优势菌种来看,在会仙湿地中增加优势植物芦苇和华克拉莎的种植,维持湿地系统中微生物多样性,利于湿地的修复和保护。
猜你喜欢
中国土壤与肥料(2021年5期)2021-12-02
中国比较医学杂志(2020年4期)2020-05-26
中国沼气(2019年1期)2019-04-13
生物安全学报(2019年3期)2019-02-15
天然产物研究与开发(2018年3期)2018-05-07
吉林农业(2018年23期)2018-01-17
中国蔬菜(2016年8期)2017-01-15
小学生作文选刊·低年级版(2016年6期)2016-06-24
阅读与作文(小学高年级版)(2014年2期)2014-02-14
植物营养与肥料学报(2010年6期)2010-10-26
