
云杉与阔叶树种新鲜凋落叶混合分解特征
2019-04-09 05:57张晓曦刘慧王博雅李佳佳雷航宇刘增文
生态环境学报 2019年2期
张晓曦 ,刘慧,王博雅,李佳佳,雷航宇,刘增文
1. 延安大学生命科学学院/陕西省区域生物资源保育与利用工程技术研究中心,陕西 延安 716000;2. 西北农林科技大学水土保持研究所,陕西 杨凌 712100;3. 西北农林科技大学资源环境学院,陕西 杨凌 712100
近数十年来,对被过度采伐的秦岭地区的天然林的保护工作使其形成了大面积的天然次生林,这些次生林在防治土壤侵蚀和涵养水源等方面发挥了重要生态作用。然而,与原始天然林相比,次生林物种组成和群落结构相对单一,物种多样性较低,尚需长时间的自然演替以使受损生态系统逐渐恢复,增强其稳定性(迪玮峙,2012)。云杉(Picea asperata)是秦岭地区重要的天然次生林树种之一,相较于当地其他次生林树种,如油松(Pinus tabuliformis)和锐齿栎(Quercus aliena),云杉林自我萌蘖更新能力较强,其林下光照条件较弱,其他树种很难在其林下生存(康冰等,2011;迪玮峙,2012)。这一更新特征在保证云杉次生林更新的连续性的同时,由于单一树种长期生长对于土壤养分的吸收利用和归还(主要是凋落物分解)具有选择性,也将不可避免地导致其林地土壤出现养分失衡、pH降低和阳离子交换量下降等其他极化现象,从而降低其林分稳定性(Mi et al.,2013;刘增文等,2008)。因此,有必要人为引入其他树种对其进行混交改造,以保证云杉天然次生林长期稳定发挥其生态服务功能(Zhang et al.,2016)。
用于混交造林的树种间应具有和谐的种间关系(Zhang et al.,2016)。从调节次生林地土壤养分平衡、维持地力的角度出发,除平衡混交种对光照、水分和土壤养分的竞争外,种间凋落物混合分解是否受到抑制也是需要加以考虑的因素。大量研究表明,凋落物混合分解中往往存在明显的非加和效应(Santos et al.,2018;Song et al.,2010;李宜浓等,2016),如果混合分解出现彼此抑制,则必将妨碍凋落物向土壤归还养分,而当某一树种凋落物分解受到促进而另一树种凋落物分解受到抑制时,则可能加剧或缓解林地土壤养分失衡现象。鉴于不同种间混合分解结果多样,且难以通过现有单种分解结果加以预测,有必要对特定的拟混交造林树种的凋落物混合分解效应进行试验验证。
目前,国内外已有大量关于凋落物混合分解的研究,研究内容也从单纯检测凋落物组成和种间基质质量差异(C、N、P、Mn、木质素和纤维素含量及其比值等)对凋落物分解速率、养分释放的影响深化至探究凋落物间养分和次生代谢物转移以及微生物生境和凋落物化学多样性的改变等混合分解的产生机理(Berger et al.,2014;Gessner et al.,2010;陈法霖等,2011;李宜浓等,2016)。然而,现有多数研究是将混合凋落物作为一个整体研究其混合效应,而对混合物中不同凋落物间的互相影响考虑尚显不足(Leppert et al.,2017;肖玲艳,2015)。在混合物中一种凋落物分解加速而另一种分解受抑的情况下,其混合分解效应可能难以被检测,导致研究结果无法反映混合凋落物中每种组分所受到的真实影响,难以用于指导营林实践。因此,本研究以云杉及5种拟用于对其次生林进行混交改造的阔叶树种的凋落物为研究对象,在室温(20-25 ℃)、恒湿条件下进行为期180 d的早期分解培养试验,研究云杉和阔叶凋落物混合对彼此分解速率的影响。此外,鉴于凋落物分解实质上是一个酶解过程(肖文娅等,2018),而混合分解往往对参与凋落物不同阶段分解的水解酶和氧化还原酶的活性也产生非加和性作用,如Li et al.(2013;2016)等研究表明,白榆(Ulmus pumila)和油松(Pinus tabuliformis)凋落物混合分解导致土壤多酚氧化酶活性较单独分解显著提高,而袁亚玲等(2018)发现针阔凋落物混合显著提高C、N循环相关水解酶的活性。本研究选取主要在凋落物分解前期(蔗糖酶)、中期(纤维素酶)以及前期和后期(多酚氧化酶)分解产生作用的3种酶(张瑞清等,2008)为代表,分析混合分解对上述酶活性的动态影响,以辅助分析混合分解效应产生的机理。本研究结果有助于进一步探讨混合分解中凋落物相互影响的机制,为秦岭地区云杉次生林混交改造提供理论依据。
1 材料与方法
1.1 研究区概况
研究区位于陕西省周至县厚畛子林场(东经108°44′-108°57′,北纬 33°46′-33°57′)。当地处于秦岭北麓中段,海拔1300-2650 m,属暖温带湿润大陆性季风气候,多年平均气温6.4 ℃,年均降水量1001.7 mm,土壤属山地棕壤和暗棕壤。当地分布有大面积的云杉、油松、日本落叶松(Larix kaempferi)、红桦(Betula albo-sinensis)和锐齿栎等天然次生林和人工林。
1.2 凋落物和土壤样品采集
在研究区内选择立地条件相似的云杉(以 Pas表示)、红桦(Ba)、灰楸(Catalpa fargesii,Cf)、太白杨(Populus purdomii,Pp)、杜仲(Eucommia ulmoides,Eu)和秦岭槭(Acer tsinglingense,At)的成熟林分,于秋末在每一林分中随机选择 10株树木,采用震动树枝的方式震落凋落物,并用凋落物收集器收集新鲜凋落物。每种凋落物充分混匀后阴干至恒重。部分凋落物于 65 ℃下烘干,用于计算阴干与烘干质量的换算系数。
采集凋落物的同时,在林分周围选择无林荒草地,在其内设置样方,收集0-10 cm土层土壤,各样方土壤均匀混合后过5 mm筛,测定含水率及饱和持水量后适当风干,以便在后续试验中控水。以处理过的荒草地土壤作为分解介质,以避免林地土壤特殊性质导致的“主场效应”(立天宇等,2015)影响分解试验结果。土壤初始理化性质如表1所示。
1.3 凋落物分解试验
分别称取云杉和阔叶树种混合凋落物(两者均为6 g)或单种凋落物12 g,装入尼龙网分解袋中(规格为10 cm×20 cm、孔径为0.5 mm,略小于云杉针叶直径),每种凋落物准备15袋。将上述凋落物(每种 5袋)竖向间隔埋入规格为 30 cm×40 cm×20 cm、盛有4.0 kg土壤的不透水培养钵中,保证凋落物与土壤充分接触以加速分解。每种凋落物设置3钵处理,用以模拟独立的3次分解过程。埋设完毕后,使用喷雾器向钵内均匀加水,调节土壤含水量为前期测定的饱和持水量的50%(该湿度略高于采集土样的实测含水量以适当促进分解,缩减试验耗时),以带孔保鲜膜封口,防止过度蒸发并为微生物提供空气。在室温(20-25 ℃)、避光条件下进行180 d模拟分解培养试验。在此过程中,每隔1周称量培养钵质量,根据失重程度进行喷雾补水,以保持土壤湿度相对恒定。

表1 供试土壤基本性质Table 1 Basic properties of the tested soil
1.4 指标测定
为准确模拟分解过程,分解进行第30、60、90、120、180天时分5次回收凋落物分解残留物。每次回收时,在3个分解钵中各随机回收1袋(即每次测定均为 3个重复),将偶然漏出分解袋的云杉针叶回收,并人工分离混合分解残留物中的两种凋落物(本试验凋落物分解程度不高,可直接通过目视分离)。迅速漂洗除去其上附着的土壤和菌丝等杂物后,将残留物于65 ℃下烘干,准确测定其质量。回收残留凋落物时,分别在分解进行第30、90、180天采集凋落物分解袋周围的分解介质(土壤)样品,同样在每种处理的3个分解钵中各采集1份土样作为重复。回收的土样经风干、磨碎、过1 mm筛后,采用 3, 5-二硝基水杨酸比色法测定其蔗糖酶和羧甲基纤维素酶活性,采用邻苯三酚比色法测定其多酚氧化酶活性(关松荫,1986)。
分别采用重铬酸钾外加热法、浓硫酸-过氧化氢消煮-凯氏定氮法、钒钼黄比色法测定凋落物初始C、N、P含量,马弗炉灰化-HCl溶解-原子吸收光度法测定其初始Mn含量(鲍士旦,2000)。
1.5 数据处理
鉴于凋落物在不同基质质量指标上的高低顺序存在明显差异,为评价6种凋落物基质质量的总体优劣,采用SPSS软件中的综合主成分分析法对凋落物基质质量指标进行分析(其中C/N和C/P比值取倒数后纳入分析范围),计算综合主成分值,作为基质质量相对优劣的排序依据(仅依据数值大小排序,不考虑正负号)(张晓曦等,2013)。计算每种凋落物5次回收时的质量残留率,应用Olson指数衰减模型对分解过程进行拟合,据此计算每种凋落物的分解常数k。
对于混合凋落物及其分解介质而言,采用下式计算混合凋落物分解常数及土壤酶活性的理论预测值:

式中,Pkm和PEm分别为混合物分解常数和土壤酶活性的理论预测值;Okp和Okb以及OEm和OEb分别为云杉和阔叶凋落物单独分解时的分解常数和土壤酶活性的实测值;0.5为云杉或阔叶树种在混合凋落物中所占的质量比。
运用SPSS 21.0软件对混合物分解速率的实测值和预测值、特定凋落物单独及在混合物中分解时的分解速率进行t检验,并对混合分解中土壤介质酶活性的实测值、预测值以及响应2种凋落物单独分解时的土壤介质酶活性进行单因素方差分析,采用 LSD法进行多重比较,显著性检验水平均为0.05。绘图均在SigmaPlot 12.5软件中完成。
2 结果与分析
2.1 凋落物基质质量
一般认为,凋落物的易分解性与其C含量、C/N和C/P比呈负相关,而与其N、P和Mn含量呈正相关。除C含量外,供试凋落物的养分含量和化学计量比间存在显著差异(图1),依据综合主成分分析结果,6种凋落物中红桦凋落物的总体基质质量最佳,表现为N、P含量最高, Mn含量中等以及C/N和C/P比最低,其次依次为灰楸、太白杨、云杉和杜仲凋落物,秦岭槭凋落物基质质量总体最差,主要表现为N含量较低和C/N比最高,但其Mn含量显著高于其他凋落物。
2.2 分解速率
依据Olson模型计算的6种凋落叶单独和混合分解时的分解常数(表2)以及混合物分解常数的理论预测值和实测值计算结果(表 3),杉×桦混合将显著促进红桦分解,从而提高混合物整体的分解速率;杉×楸和杉×杜混合将显著抑制灰楸和杜仲分解,从而降低整体分解速率;杉×杨和杉×槭混合时针阔凋落物显著促进彼此分解。具体至前期分解过程(图2),杉×桦混合在分解第180天时显著促进彼此分解,呈正非加和效应;杉×楸混合在第 60、90、120天时显著抑制彼此分解,呈负非加和效应,而在第180天时云杉分解受到显著促进,灰楸分解则受到显著抑制,总体转呈加性效应;杉×杨混合在第120天时促进彼此分解,整体呈正非加和效应,第180天时仅太白杨分解受到显著促进,但整体呈加和效应;杉×杜混合在分解期间整体呈负非加和效应,其中第60天时抑制彼此分解,而在120-180 d时抑制杜仲凋落物分解;杉×槭混合在第120-180天时显著促进秦岭槭凋落物分解,整体呈正非加和效应。总体而言,混合分解过程中凋落物间的相互影响随分解时间延长而更加显著(图2)。

表2 Olson模型计算的分解常数(k)Table 2 Decomposition constant(k) calculated by the Olson model

图1 供试凋落物基质质量Fig. 1 Substrate quality of the tested litter

表3 混合凋落物分解常数k的实测值和预测值Table 3 The observed and predicted decomposition constant k of litter mixtures
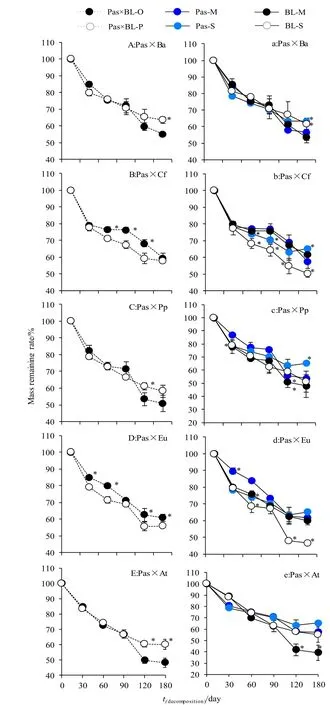
图2 凋落物分解过程Fig. 2 Decomposing processes of litter
2.3 酶活性动态
混合分解对土壤蔗糖酶活性影响普遍不显著(图3),仅杉×杨混合在分解第90天时对其产生显著促进效应,使蔗糖酶活性较预测值以及云杉、太白杨单独分解时显著提高,杉×槭混合分解使蔗糖酶活性较秦岭槭单独分解时显著提高。杉×桦、杉×杨和杉×槭混合分别在第120-180、30、180天以及第180天时产生促进效应,使羧甲基纤维素酶活性较预测值以及云杉、红桦、太白杨和秦岭槭单独分解时显著提高;杉×楸混合在第 180天时产生促进效应,使纤维素酶活性较预测值和云杉单独分解显著提高,但较灰楸单独分解显著降低;杉×杜混合在第30天时产生抑制效应,使纤维素酶活性较预测值以及云杉、杜仲单独分解时显著降低,在120-180 d则使其活性较云杉单独分解显著提高而较杜仲单独分解显著降低。杉×桦、杉×杨和杉×槭混合在第 30天时均对多酚氧化酶活性产生促进效应,使其活性较预测值以及云杉、红桦、太白杨和秦岭槭单独分解时显著提高,杉×楸和杉×杜混合则完全相反;5种混合在第90天时普遍未产生非加和效应,但多使多酚氧化酶活性较云杉单独分解显著降低;5种混合处理在第180天时普遍产生促进效应,使多酚氧化酶活性较预测值、云杉以及阔叶树种凋落物单独分解时显著提高。
3 讨论

图3 分解过程中酶活性动态Fig. 3 Dynamics of the enzymatic activities during decomposition
本研究结果表明,云杉与5种阔叶凋落物混合分解均产生显著非加和效应,这与 Mao et al.(2015)、Santonja et al.(2017)和李英花等(2017)的研究结果一致。现有多数研究表明,混合分解中基质质量较高的凋落物可促进基质质量更差的凋落物的分解,而其自身分解则受到一定限制(Chomel et al.,2015;郭鲲等,2015;Liu et al.,2016),本研究部分结果与之相符。例如,杉×槭混合时,秦岭槭凋落物分解在120 d和180 d时受到显著促进(图 2e)。尽管两者基质质量相似,但云杉凋落物C/N比更低,N素相对丰富,因此在混合分解中云杉凋落物中的N可能被动(通过淋溶)或主动(通过菌丝)地转移至秦岭槭凋落物,从而促进其分解(Zhang et al.,2014)。此外,混合分解可能通过改变凋落物的空间结构而为微生物提供更优的水分、空间、温度条件,从而加速其繁殖,并提高其活性,最终加速凋落物的分解(Gessner et al.,2010)。(前期研究表明蔗糖酶、羧甲基纤维素酶和多酚氧化酶分别主要在凋落物分解前期、分解中期以及分解前期和后期参与分解(张瑞清等,2008),本试验主要涉及前半程分解,因此与之对应的时间为分别为分解试验第 30天、分解试验第180天和第30天左右。本研究中,杉×槭混合时分解介质中蔗糖酶、羧甲基纤维素酶和多酚氧化酶在其参与分解的主要时期较之秦岭槭单独分解时均有显著提高(图3),表明混合分解对分解介质酶活性具有显著非加和效应,与袁亚玲等(2018)的结论相似。鉴于微生物是土壤酶的主要来源之一(肖文娅等,2018),上述事实也证实了混合分解提高了分解介质中微生物的活性。另外,有研究表明,混合物中基质质量较高的凋落物可吸引大量分解者定殖,由于空间(表面积)限制,过剩的微生物可能定殖在与其毗邻的凋落物上,从而加速其分解(郭鲲等,2015)。另外,多数研究认为,针阔混合分解将显著促进针叶凋落物的分解及其养分释放(袁亚玲等,2018;覃宇等,2018),相应地,阔叶凋落物分解则受到一定抑制,本研究结果与之部分结果相符,例如杉×楸混合在分解第180天时灰楸凋落物分解受到显著抑制,而云杉凋落物分解受到显著促进,其主要原因可能是分解过程中出现养分(主要是N和P)和次生代谢物的转移(Zhang et al.,2014),导致前者养分损失,分解抑制物含量增加,不利于微生物分解,而后者则相反。例如凋落物浸水后将释放一定的N、P养分(如 NH4+、NO3-和 PO43-,Del Giudice et al.,2017),而笔者的前期研究表明针叶凋落物浸提液中含有大量的酚、萜类衍生物,其中云杉凋落物浸提液中检出α-蒎烯、β-蒎烯、樟脑、异龙脑、氧化芳樟醇、苯甲酸和邻苯二甲酸二丁酯等(Zhang et al.,2018)。因此,混合分解中灰楸凋落物可能通过淋溶损失大量养分促进云杉分解而减缓自身分解,而云杉则因释放次生代谢物抑制分解者活性从而妨碍灰楸分解,同时在一定程度上提高自身基质对分解者的可利用程度。本研究中,云杉凋落物的加入使分解介质中纤维素酶活性(第180天)和多酚氧化酶活性(第30天)较灰楸单独分解时显著降低,其可能原因为云杉释放的上述酚、萜类衍生物抑制芽孢杆菌(Cho et al.,2015)、白腐菌、褐腐菌、丝状真菌(Roy et al.,2006)、青霉、毛霉(Hammer et al.,2003;Shi et al.,2015)和黑曲霉(Shi et al.,2015)等分解者的生长,从而抑制酶分泌,并通过与蛋白质结合改变酶构象或通过酸化土壤(Guan et al.,2014)降低酶活性(Adamczyk et al.,2015;Adamczyk et al.,2013;Triebwasser et al.,2012),从而抑制灰楸凋落物的分解。此外,本研究中云杉凋落物的Mn含量最低,其与灰楸凋落物混合后对Mn的供应产生限制,不利于多酚氧化酶的合成,这也可能是造成混合分解介质中多酚氧化酶活性低于灰楸分解介质中的,抑制灰楸分解的原因之一(Liu et al.,2016)。
然而,与前人研究相反,本研究表明凋落物基质质量差异较大的情况下,基质质量较差的凋落物亦可促进基质质量较高的凋落物的分解,如云杉显著促进了混合物中红桦和太白杨凋落物的分解。其原因可能是混合分解提高了凋落物基质质量的异质性(Chomel et al.,2015),一定程度上为微生物提供了更为全面的养分(Guénon et al.,2017),从而促进其生长并提高其活性,上述推测也得到了土壤酶活性检测结果的支持:杉×桦和杉×杨组合均使分解介质中蔗糖酶、纤维素酶和多酚氧化酶活性较对照以及上述树种凋落物单独分解时显著提高(图3)。另一方面,有研究表明,混合分解中养分的被动转移并非绝对依从养分浓度梯度,而是可从低基质质量凋落物向高基质质量凋落物逆向转移(Leppert et al.,2017)。本文推测,同时定殖于针阔凋落物的真菌将获取的养分优先供给阔叶一侧的生长和有机物降解活动,因此可能主动从针叶凋落物处获取养分转移给阔叶凋落物,从而加速其分解。除此之外,有研究表明,较之单种凋落物,混合凋落物中的土壤动物(如螨类和其他小型节肢动物)数量显著提高,从而促进混合凋落物的分解进程(李宜浓等,2016)。本研究中,混合分解可为微生物提供多样的碳源和养分(不同凋落叶间基质质量存在显著差异),使其避免了因资源供应条件大幅变动而导致种群数量剧烈波动,从而保证了土壤动物稳定的食物来源;且微生境的改变为土壤动物提供了更有利的繁殖和发育的环境,从而促进土壤动物种群数量增加(李宜浓等,2016)。当然,由于针阔叶凋落物的适口性差异,云杉凋落物质地坚硬,且含有较多酚类和萜类次生代谢物,适口性较差(Zhang et al.,2018),混合分解对土壤动物的促进作用最终更倾向于促进阔叶凋落物的分解。在后续研究中,可通过采用不同孔径的分解袋限制不同体型的土壤动物与凋落物的接触,配合镜检分类,进一步研究混合分解对不同分解者的具体影响(张瑞清等,2008)。
需要说明的是,(1)凋落物分解是一个复杂过程,凋落物混合对微生境、分解者和酶所产生的正向或负向影响以及凋落物间的物质(养分、次生代谢物)转移往往同时作用于混合分解过程(李宜浓等,2016),因此非加和效应以及凋落物间彼此影响的具体结果应当视为上述多种机制的相互抵消后的最终结果,而非某种机制单独作用。当然,本文为简化描述,在观察结果为抑制效应时着重分析产生负向影响的因素,反之亦然。(2)混合分解效应与针阔叶分解的相互影响均随时间增强,至试验末期才达到显著水平。上述结论与 Zhang et al.(2014)的研究结果相似,其原因可能是凋落物分解前期以淋溶形式为主,因此由凋落物基质质量差异和分解者化学和物理环境改变导致非加和效应难以体现;同时,由于分解抑制物的降解,凋落物间的养分转移产生的非加和效应在分解后期更明显(Liu et al.,2016);此外,随分解进行,微气候和凋落物的化学组成可导致分解者的种群演替,后期分解者趋向于以大型食腐生物为主,而该类生物可产生较微生物而言更为明显的非加和效应(Liu et al.,2016)。(3)本研究结果表明,凋落物分解过程中,尽管非加和效应以及凋落物间相互影响的多次实测结果与Olson模型的预测结果在总体趋势上基本一致,然而对凋落物混合分解效应以及凋落物间相互影响的判断中,两种方法仍然存在一定差异,上述差异出现的具体原因尚有待进一步研究分析,但在应用模型预测凋落物分解过程时尚需慎重考虑。(4)纯林混交改造涉及树种对光照、水分和空间的竞争,彼此间的化感影响以及混交对养分循环影响等多个方面的问题,本文仅从混交应不妨碍林地凋落物分解(即不限制林地养分循环)这一原则考虑,以针阔叶凋落物对彼此分解影响的实测情况为依据,认为除杜仲外其他4种阔叶树种均适宜于与云杉进行混交。当然,后续工作中尚需全面考虑各树种凋落物的分解特征(养分释放和后续的腐殖质形成等),佐证和丰富本研究的结论。
4 结论
混合分解非加性效应及凋落物间相互影响均随时间延长趋于明显。至分解试验结束时,杉×桦混合促进红桦分解;杉×楸混合中云杉分解受到显著促进,灰楸分解受到显著抑制,整体呈加性效应;杉×杨和杉×槭混合促进太白杨和秦岭槭分解;杉×杜混合抑制杜仲分解,上述趋势与Olson模型预测有所差异。混合分解对土壤蔗糖酶、羧甲基纤维素酶和多酚氧化酶的非加和影响与其对凋落物分解的影响一致。总体而言,某些情况下混合分解的总体非加和效应因凋落物间的相互作用而无法被检测,但鉴于不同凋落物分解特征的差异,混合后其对彼此分解的影响仍可能干扰林地物质循环。仅从引入阔叶树种后应不妨碍林地凋落物分解的角度考虑,除杜仲外,其他4种阔叶树种均适宜于云杉纯林改造。
猜你喜欢
落叶果树(2021年6期)2021-02-12
昆明医科大学学报(2020年12期)2021-01-26
现代农村科技(2020年7期)2020-12-19
文苑(2020年12期)2020-04-13
花卉(2020年6期)2020-01-07
海峡姐妹(2019年1期)2019-03-23
中成药(2018年10期)2018-10-26
启迪与智慧·教育版(2018年12期)2018-02-26
现代园艺(2017年22期)2018-01-19
上海农业学报(2017年4期)2017-04-10
