
外源NO对重楼种子活力及超弱发光的影响
2019-04-09 08:32李海明李梅香孟艳艳宋发军戴甲培李泽华
广东农业科学 2019年2期
李海明,李梅香,孟艳艳,宋发军,戴甲培,李泽华,耿 红
(1.中南民族大学生命科学学院/生物技术国家民委重点实验室/武陵山区特色资源植物种质保护与利用湖北省重点实验室,湖北 武汉 430074;2.中南民族大学武汉神经科学与神经工程研究所,湖北 武汉 430074)
【研究意义】重楼(Paris)为百合科重楼属(Trilliaceae)多年生草本植物,分布于云南、广西、江西、浙江、湖南等地[1],其正品药源包含七叶一枝花(Paris polyphyllavar.chinensis,PPC)和滇重楼(Paris polyphyllavar.yunnanensis,PPY)[2]。重楼的根状茎经济价值很高,是云南白药等药的重要组分之一,近年来受到了掠夺式采挖,导致重楼的野生资源锐减[3]。利用种子进行人工繁殖是缓解供需矛盾的有效途径,已得到人们的广泛关注。然而重楼种子的休眠期长且休眠程度高、萌发速度慢、萌发率较低。自然条件下,重楼种子一般需要两冬一夏才能开始萌发,15个月的出苗率仅为46.2%,漫长的休眠期和萌发周期会使大量种子的生活力下降甚至丧失生命力[4]。因此,随着市场对重楼种子需求的不断增加,重楼种子质量的高低、种子活力的大小和种子处理方式直接影响人工栽培重楼的产量和质量。目前,国内外对重楼种子休眠原因和解除休眠方法、种子繁育技术等方面展开了系列研究[5-8],而关于重楼种子质量检验技术和生活力检验技术等方面的研究较少,重楼种子超弱发光方面的研究更是未见报道。生物的超弱发光是生物体进行新陈代谢和体内生理生化反应过程中细胞自发辐射的非常微弱的光子流,无论是动植物还是单细胞生物,其体内都会有超弱发光的现象[9]。其中,种子的超弱发光通常由自由基和有丝分裂产生。自由基主要是由于种子代谢活动产生,而有丝分裂与细胞生长分裂、细胞内核酸的含量变化有关[10-12]。因此,种子的超弱发光不仅可以反映种子的生命活动状态、细胞内外的新陈代谢和信息交换的程度,还可以作为检测种子活力的一个重要指标。【前人研究进展】目前,国内对种子萌发过程中自发超弱发光的研究较少,尤其是对于包含重楼在内的具有深度休眠特性的药用植物种子而言,报道更是少见。现有研究表明,种子内部的生理生化反应、细胞活动等任何生成或利用ATP、NADPH、NADP的反应均与种子的自发超弱发光有着密切联系[13]。其原因可能是在这些代谢反应中,细胞内的部分能量以光子的形式释放出来,所以反应越剧烈,细胞活动越活跃,种子的自发超弱发光的强度就越高。种子的超弱发光可以借助不同的仪器设备测定。田茜等[14]研究了人工老化过程中超弱发光的变化及其与种子活力、ATP含量的关系。王春芳等[15]利用BPCL-2微弱发光测量仪完成了稻谷和小麦种子发芽率检测。赵燕燕等[16]利用自制的测量系统对高粱种子萌发过程中的超弱发光特性进行了研究。而利用新型的超弱发光图像检测装置可以达到更加直观、准确和实时检测种子的超弱发光变化情况的目的。【本研究切入点】选用本单位自行搭建的电子倍增式CCD(EMCCD)生物光子成像系统,对不同类型和不同处理的重楼种子活力进行检测。该检测系统具有超高灵敏性,在对极微弱光信号的实时快速探测方面,可真正做到单光子事件的检测[17],并且此装置可实时监测生物的不同部位及生物在不同代谢阶段或不同环境下的超弱发光及其变化情况。已有研究表明,NO作为一种活性分子,在适宜浓度下可以促进多种植物种子的萌发,并提高其萌发率[18-25]。我们前期研究也发现,适宜浓度的外源NO供体硝普钠(sodium nitroprusside,SNP)可以显著缩短种子的萌发周期,且提高重楼种子的萌发率[26]。【拟解决的关键问题】分别以七叶一枝花和滇重楼的种子为研究对象,通过检测SNP处理和对照的种子在萌发期间的超弱发光强度,建立重楼种子超弱发光值与其萌发率的关系,分析SNP对重楼种子超弱发光的影响,并建立通过超弱发光值预测种子生活力的模型,为重楼种子质量和发芽率的快速判定提供基础。
1 材料与方法
1.1 试验材料
供试材料为中南民族大学巴东实验基地采摘的3年生七叶一枝花与滇重楼的成熟蒴果,经万定荣教授鉴定。去颖壳和果皮,并以人工揉搓的方法去除红色浆果的外种皮和果肉,得到乳白色较坚硬的种子。经自来水充分洗涤后室温下晾干,存于4 ℃备用,具体方法参照文献[27]。试剂为购自TIANDZ的亚硝基铁氰化钠(SNP)。
主要仪器:探测仪EMCCD(日本Hamamatsu公司,ImagEMCCDC9100-13);专用暗箱(定做,上海欧海化工机械设备有限公司);变焦透镜(DO5095,美国Navitar公司,F stop值为1. 0)。
1.2 试验方法
1.2.1 浸种处理 挑选健康饱满的重楼种子,七叶一枝花与滇重楼各两份,每份120颗,分装于250 mL锥形瓶中,25 ℃条件下用蒸馏水浸种48 h,至种子发白为止。将分析纯级别的SNP配成100 μmol/L溶液,用该溶液浸种为48 h,以蒸馏水浸种为对照。
1.2.2 接种处理 浸种完成后,在超净工作台将所有种子用蒸馏水清洗5遍,用无菌水清洗3遍,并用75%乙醇清洗30 s并不停摇晃,再用无菌水清洗3遍。接着用0.1%升汞处理种子12 min,处理过程中不停摇晃锥形瓶。最后将升汞处理过的种子用无菌水清洗5遍。并转移至铺有2层无菌滤纸的培养皿中进行暗培养,每皿120粒,温度19(±2)℃。
1.2.3 观察记录 种子培养期间每隔7 d补1次水,补水量均为1 mL,保证培养皿内的种子湿润。培养过程中记录种子的形态变化。以胚根刚刚突破种皮为标准,统计种子培养0、10、20、30、40、50、60、70 d的发芽率(Gr),计算发芽指数(Gi):

式中,Gt培养t天的种子萌发数,Dt为萌发天数。
1.2.4 种子超弱发光值测定 重楼种子的自发超弱发光强度测定参照宋述燕[17]方法进行。不同处理重楼种子分别放置在专用暗箱中,种子超弱光子信号经连接在EMCCD的变焦透镜成像在EMCCD探测器上,然后经光电效应转换成电子图像,再用计算机对图像进行分析。培养后0、10、20、30、40、50、60、70 d,每份种子各取样9粒,每3粒1组,共测定3组重楼种子样品在萌发期间的自发超弱发光强度,3次重复,取平均值。光值采集参数设置均为:曝光时间60 ms,间隔0.3 s,增益1 200,持续采集90 min,试验过程中不存在外部漏光的可能。数据处理时删减前30 min的发光值,以除去自然延迟发光对实验结果的影响。
利用SPSS16.0和Graphpad prism 7进行数据分析和作图。
2 结果与分析
2.1 不同处理重楼种子萌发过程形态特征
由图1可知,滇重楼种子经100 μ mol/L SNP处理30 d时即有胚根突破种皮,而对照种子培养50 d后才有胚根突破种皮。SNP处理的种子萌发时间比CK提前20 d,萌发培养70 d时SNP处理种子的胚根明显长于CK。此外,SNP处理的七叶一枝花和滇重楼种子均比CK提前20 d萌发。

图1 不同处理重楼种子萌发过程中的形态变化Fig.1 Morphological changes in Paris seed germination process of different treatment groups
2.2 不同处理重楼种子发芽率和发芽指数
从表1可以看出,SNP处理的七叶一枝花和滇重楼种子均在培养30 d发生萌发现象,即胚根开始突破种皮,萌发时间分别比CK提前20 d。萌发培养60 d,SNP处理的两组种子萌发数均已超过50%,而对照种子的萌发率仅有25%左右。萌发培养70 d,两种重楼种子在SNP处理的萌发率都达到90%,分别比对照高36.41%、47.5%。此外,SNP处理的滇重楼和七叶一枝花种子发芽指数也明显高于CK。结合图1与表1发现,外源NO具有促进种子萌发、提高种子活力和改善种质的作用,且这种作用在七叶一枝花中表现更为明显。
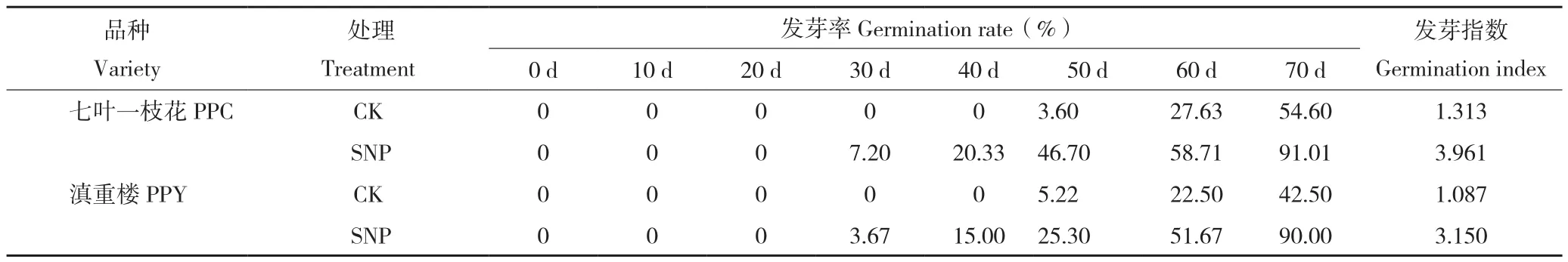
表1 不同处理重楼种子的发芽率Table 1 The seeds germination rate of Paris with different treatments
2.3 不同处理重楼种子超弱发光强度变化
重楼种子萌发过程中体内自发超弱发光强度变化如图2所示。由图2A可知,种子培养0~30 d为萌发前期(种子培养开始到胚根开始突破种皮),处理和对照七叶一枝花种子的发光强度无显著差异。在萌发中期(胚根突破种皮到胚根的长度达到1.5 cm,30~60 d),SNP处理中,七叶一枝花种子的自发超弱发光值迅速升高,萌发40~60 d,SNP处理种子的发光强度显著高于对照,在萌发后期(胚根长度>1.5 cm,60~70 d)又下降,种子发光强度最高峰值出现在萌发40 d。而对照种子在不同萌发时间节点上发光强度变化不大;在萌发中期,种子发光强度略有降低;萌发后期,又有小幅度抬升。滇重楼种子和七叶一枝花种子的发光强度变化趋势相似。其中SNP处理种子的发光值也在萌发40 d明显升高且显著高于对照,萌发50~60 d其发光强度也高于对照但差异不显著;其他时间点的发光强度对比中,两组差异均不显著(图2B)。未经SNP处理情况下,七叶一枝花和滇重楼的种子在不同时间段的发光强度无明显差异。与此同时,SNP处理后七叶一枝花种子和滇重楼的种子发光强度的变化趋势相同,但是在萌发50 d与萌发60 d时,七叶一枝花种子的发光强度均明显高于滇重楼的。

图2 不同处理重楼种子的自发超弱发光强度Fig. 2 The spontaneous ultra-weak luminescence intensity of Paris seeds with different treatments
2.4 不同处理重楼种子的发光强度与发芽率变化趋势
七叶一枝花对照在种子培养40 d后,发芽率为0,发光强度无明显变化。培养40~60 d种子开始萌发,此时发光强度有轻微下降。培养70 d,CK种子大量萌发,与此同时该阶段种子的发光强度有上升趋势(图3A)。SNP处理中,七叶一枝花种子在培养20 d时发芽率为0、发光强度变化波动较小,培养30~40 d种子开始萌发、发光强度明显上升,随着萌发种子数增加(50~60 d),发光强度保持较高水平,随后在培养70 d时下降(图3B)。滇重楼对照(图3C)与七叶一枝花的对照变化趋势相同,滇重楼的SNP处理(图3D)的变化趋势也与七叶一枝花SNP处理中种子的变化类似。
3 讨论
NO作为细胞中的一种重要信号分子,参与植物体内如防御机制和抗逆反应等多个生理过程[18-25]。此外,有研究发现硝普钠(SNP)对亚麻[28]、玉米[29]、九节木[30]和蒺藜苜蓿[31]等种子的萌发也有一定促进作用。我们前期研究也发现,适当浓度的外源NO可以大幅减短重楼种子的萌发周期,且提高种子的萌发率[26]。然而,外源NO对重楼种子活力及萌发期间种子超弱发光的影响还未见报道。因此我们从自然超弱发光的角度,探讨了自然条件下和SNP处理后重楼种子萌发过程中发光强度的变化。
结果表明,SNP处理种子的萌发形态变化和萌发效果均明显早于CK,表明SNP在一定程度上提升了种子活力,达到促萌作用。此外,不同试验的重楼种子,在萌发后0~30 d内种子内部自发超弱发光无显著差异,但在萌发后40~60 d SNP处理明显高于CK,随后差异不显著。其原因可能是由于在种子萌发前期,重楼种子正在适应萌发条件,克服种子休眠的生理现象。因此该时间段内,种子内部的生理生化反应与细胞活动均较少。SNP处理后,短时间内可能不会对种子发光现象产生影响,存在滞后期。种子萌发40 d开始,SNP处理中种子内部的生理生化反应与细胞活动加剧,种子萌发数迅速增加,该时期种子的自发超弱发光强度也最大。随着新增萌发种子数的减少,种子内部的有丝分裂等生理活动逐渐减弱,种子自发超弱发光强度也逐渐降低,与对照差别不大。对照重楼种子的自发超弱发光强度最大值的出现时间均与新增萌发种子数最高值的出现时间相吻合。

图3 不同处理重楼种子发光强度与发芽率的关系Fig. 3 The relation between luminescence intensity and germination rate of Paris seeds with different treatments
已有研究表明,小麦种子的发芽率与发光强度呈线性相关关系,而水稻种子的发芽率与发光强度符合非线性相关关系[15]。白菜与萝卜新陈种子的研究表明,新种子的发芽率与发光强度呈正相关,但陈种子的发芽率与发光强度呈负相关[32]。因此,了解种子发光强弱可为判定种子活力、预测种子发芽率提供理论依据。但从本试验发芽率与超弱发光强度的比较可以看出,无论是对照还是处理的种子萌发率与发光强度没有相同的变化趋势,均不呈明显的线性关系,这与小麦和萝卜等的研究结果不同。原因可能是上述4种类型的种子均为浅休眠型,而重楼种子在萌发过程中是深度休眠,且具有形态休眠和生理休眠的双重特性,从而使重楼种子发光强度的变化趋势呈现峰状变化。另外,不同试验处理方式不同,也会造成较大差异。本试验取样时间为种子萌发进程的前70 d,在此时段内SNP处理组种子的萌发程度已达90%,但对照只有50%种子萌发。在自然条件下,种子萌发进程第70天后的发芽率与发光强度是否有相关性还未知,有待深入研究。
4 结论
综上所述,本试验采用的EMCCD能有效检测种子萌发过程中的自发超弱发光强度,可准确得到待测种子二维光强分布的空间与时间信息,据此判断种子活性的强度,并建立有效的通过超弱发光值预测种子生活力的模型,为解决重楼种子质量和发芽率的快速判定问题提供基础。EMCCD对重楼种子二维光强进行检测,表明自发超弱发光能给种子的萌发进程提供重要相关信息,预示对光子的研究技术在农业科研上有广阔的利用前景。另外,我们从形态观察、发芽率、发芽指数和发光强度等方面综合得出,SNP对改善重楼种子萌发质量有明显作用,在种子萌发40~60 d时效果尤为明显。其改善机制是否因为提高了重楼种子体内的NO等信号分子的浓度,从而加快了种子内部的生理生化反应或促进了种子的萌发进程还有待进一步探索。
猜你喜欢
今日农业(2022年6期)2022-07-05
花卉(2021年12期)2021-12-07
安徽农业科学(2021年14期)2021-09-03
山西农业科学(2021年8期)2021-08-17
科学导报(2020年8期)2020-03-12
山西农业科学(2020年1期)2020-02-27
红楼梦学刊(2020年2期)2020-02-06
诗潮(2019年10期)2019-11-19
当代陕西(2019年6期)2019-04-17
现代园艺(2016年2期)2016-03-15
