
感染大肠杆菌F17湖羊羔羊脾脏中差异circRNA分析
2019-04-03 05:58:48邹双霞金澄艳鲍建军王悦陈炜昊吴天弋王利宏吕晓阳高雯王步忠朱国强戴国俊师东方孙伟
中国农业科学 2019年6期
邹双霞,金澄艳,鲍建军,王悦,陈炜昊,吴天弋,王利宏,吕晓阳,高雯,王步忠,朱国强,戴国俊,师东方,孙伟,6
(1扬州大学动物科学与技术学院,江苏扬州225009;2南京新九州农牧科技有限公司,南京210000;3江苏西来原生态农业有限公司,江苏泰州225300;4扬州大学兽医学院,江苏扬州225009;5东北农业大学动物医学院,哈尔滨150030;6扬州大学教育部农业与农产品安全国际合作联合实验室,江苏扬州225009)
0 引言
【研究意义】羊大肠杆菌病是规模化羊场最为常见高发的细菌性疾病之一,传统沿用的抗生素治疗方案存在诸多缺陷。利用RNA-seq筛选与不腹泻绵羊大肠杆菌病有关的circRNA是分析绵羊抗病分子机制的基础,从而发现与抗病性状相关的候选基因。【前人研究进展】环状RNA(circular RNA,circRNA)是一种特殊的内源性非编码 RNA(non-coding RNA,ncRNA),是继微小RNA(microRNA,miRNA)及长链非编码RNA(long noncoding RNA,lncRNA)后的 RNA 家族又一研究新热点[1]。SANGER 等[2]利用电镜首先在植物感染的类病毒中发现了这些以共价键形成的闭合环状单链 RNA 分子,它们具有高度的热稳定性。1990年,研究人员在酿酒酵母中发现 20S RNA 没有自由的5' 端和3' 端,通过电镜观察发现,这是呈环形的RNA分子[3-4]。随后人们陆续在丁型肝炎病毒中发现 circRNA[5],也发现在小鼠的睾丸内含有性别决定基因(sex-determining region Y,Sry)转录而来的circRNA[6],还证实了circRNA 也存在于人体细胞[7]。虽然很早就发现circRNA 广泛存在于各种不同类型的细胞中,但在过去的几十年里,circRNA的研究却始终进展缓慢,其生成机制和表达调节机制目前尚不完全清楚[8]。长久以来,circRNA 被认为是pre-mRNA 加工过程中选择性剪接产生的副产物。事实上仅很少的 circRNA 被发现是选择性剪接过程中产生的外显子[9]。最近的研究[10]发现,circRNA 不是mRNA 成熟过程中的副产物,而与mRNA一样,是pre-mRNA 加工后的一个重要产物,并且circRNA 的加工机制能与 mRNA 形成竞争。同时,经典的剪切信号和剪切机制对于反向剪切来说也是必需的[11]。【本研究切入点】目前,关于绵羊抗病方面的研究主要集中在疾病防治方面[12-13],而重要的抗病分子机制研究却鲜有报道。【拟解决的关键问题】本研究利用RNA-seq,首次筛选出对大肠杆菌F17菌株不腹泻与腹泻型个体中差异表达的circRNA,利用Miranda分析circRNA- miRNA-mRNA相互作用,寻找miRNA的靶基因,在此基础上利用q-PCR进行验证。本研究从circRNA层面上,加深了对绵羊拮抗大肠杆菌F17菌株的认识,同时有望确定绵羊拮抗大肠杆菌F17菌株的功能基因,解决中国地方羊品种对大肠杆菌病的抗病育种关键问题,为今后制订抗大肠杆菌病遗传选育策略奠定基础和提供理论依据。
1 材料与方法
1.1 试验设计和样本采集
试验用羊于2016年12月购自江苏西来原生态农业有限公司。随机选择包括性别、生长发育良好、日龄体重相近的18只3日龄羔羊,并将羔羊全部隔离饲养。先用10%羔羊奶粉(不含有任何抗生素和微生态制剂)饲喂,以确保在试验前适应饮食需要。5日龄时开始饲喂 12.5%羔羊奶粉和大肠杆菌 F17菌液(4.6×108CFU·mL-1),同时保证自由饮水。每天记录羔羊粪便形态[14],在部分羔羊持续腹泻 2d后,将羔羊分为不腹泻组和腹泻组,并对羊进行安乐死。用RNAlater收集每只羔羊的肝脏、脾脏、十二指肠、空肠和回肠,并立即在液氮中冷冻带回实验室以进行RNA提取。因为主要研究羔羊的免疫状态,而脾脏是动物体最大的免疫器官,所以选用脾脏这个器官作为试验对象。
1.2 文库建设和测序
分别从不腹泻组和腹泻组中选择了3个典型样本并从绵羊脾脏提取总RNA,使用NanoDrop 2000超微量分光光度计和Agilent 2100生物分析仪进行质控。用 Ribo-Zero TM试剂盒(Epicenter, Madison, WI,USA)除去核糖体RNA。将RNA片段化(平均长度约为 200bp),然后通过逆转录合成和纯化 cDNA。使用Qubit®dsDNA HS测定试剂盒进行PCR扩增和纯化后,选择使用NEBNext®Ultra™RNA文库制备试剂盒进行文库构建。在上海欧易生物医学科技有限公司使用Illumina HiSeq 2500平台,对文库进行末端配对测序(测序读长为150 bp)。
1.3 鉴定circRNA
circBase[15]数据库中只收录了人、小鼠、线虫、矛尾鱼和腔棘鱼5个物种的circRNA序列。由于绵羊不属于上述物种之一,使用 CIRI[16]软件从头预测circRNA。根据 circRNA在基因组上的位置,可以将circRNA分为以下 5类:exonic circRNA、intronic circRNA、antisense circRNA、sense overlapping circRNA、intergenic circRNA。
1.4 差异表达分析
差异表达分析旨在找出不同样本间存在差异表达的circRNA,得到差异表达circRNA之后,对其来源基因做Gene Ontology(GO)和KEGG Pathway显著性分析。DESeq[17]适用于有生物学重复的实验,可以进行样品组间的差异表达分析,获得两个生物学条件之间的差异表达circRNA列表;对于没有生物学重复的实验,则使用 edgeR[18]进行差异表达分析,获得两个样品之间的差异表达circRNA列表。
1.5 GO和KEGG通路分析
对差异表达转录本进行GO富集分析,对其功能进行描述(结合GO注释结果)。统计每个GO条目中所包括的差异转录本个数,并用 Fisher's exact test计算每个 GO条目中差异转录本富集的显著性。KEGG[19]是有关 Pathway的主要公共数据库,利用KEGG数据库对差异转录本进行Pathway分析(结合KEGG注释结果),并用Fisher's exact test计算每个Pathway条目中差异转录本富集的显著性。
1.6 circRNA-miRNA-mRNA互作研究
circRNA可以作为 miRNA的靶标分子,受到miRNA的调节。由于circRNA包含多个miRNA结合位点,通过circRNA-miRNA相互作用分析,可以帮助解析作为海绵发挥作用的 circRNA的功能和作用机制。对于动物,使用Miranda[20-21]软件来预测与miRNA结合的circRNA以及miRNA的靶基因,根据miRNA靶基因的功能注释来阐明此部分circRNA的功能。
1.7 验证差异转录本的表达水平
为验证筛选的DE circRNA在拮抗过程中发挥的作用,笔者用q-PCR检测了不腹泻和腹泻羔羊脾脏组织中DE circRNA和miRNA靶基因的表达水平,使用2-ΔΔCt方法将每个RNA的相对定量归一化为GAPDH,circRNA的引物见表1。
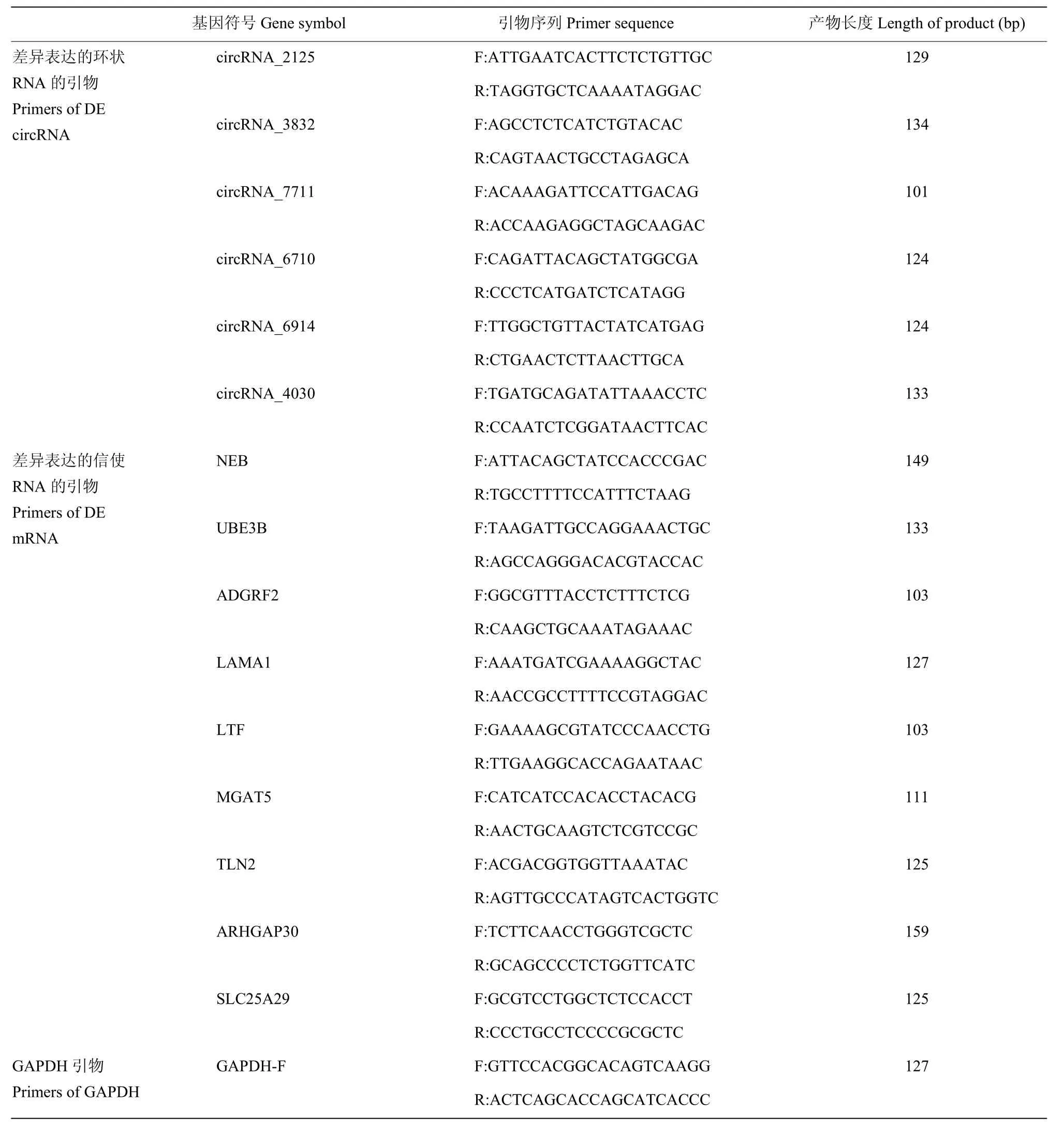
表1 GAPDH,DE circRNA和mRNA的引物Table 1 The primer of GAPDH, DE circRNAs and mRNAs
1.8 统计分析
使用SPSS软件(版本20.0)分析所有数据,使用单因素方差分析(ANOVA)分析差异转录本的相对表达量,并使用Tukey检验进行多重比较。P<0.05被认为具有统计学意义。每组包含3个样品,每个试验重复3次。
2 结果
2.1 绵羊脾脏中转录本的鉴定
在绘制参考序列后,鉴定出已知的 7 730个circRNA。由图1可以看出,circRNA长度主要分布于200—900 bp,平均长度为1 943bp,GC含量大约在43.5%,统计circRNA的外显子数目主要为2—4个。统计 circRNA序列中反向剪切位点的可变剪切信号(GT-AG)情况,并绘制图形,结果见下图 2。统计circRNA类型见下图3,其中sense overlapping circRNA占92.11%,exonic circRNA占3.27%,intergenic circRNA占3.18%,intronic circRNA占0.88%,antisense circRNA占0.56%。将circRNA与基因元件进行比较,从而探索circRNA在基因组上的分布,统计各条染色体上或scaffold上预测得到的circRNA数目情况,并绘制图形,结果见下图4,其中主要分布在3条染色体上,分别是NC_019458.2(853)、NC_019459.2(772)、NC_019460.2(787)。
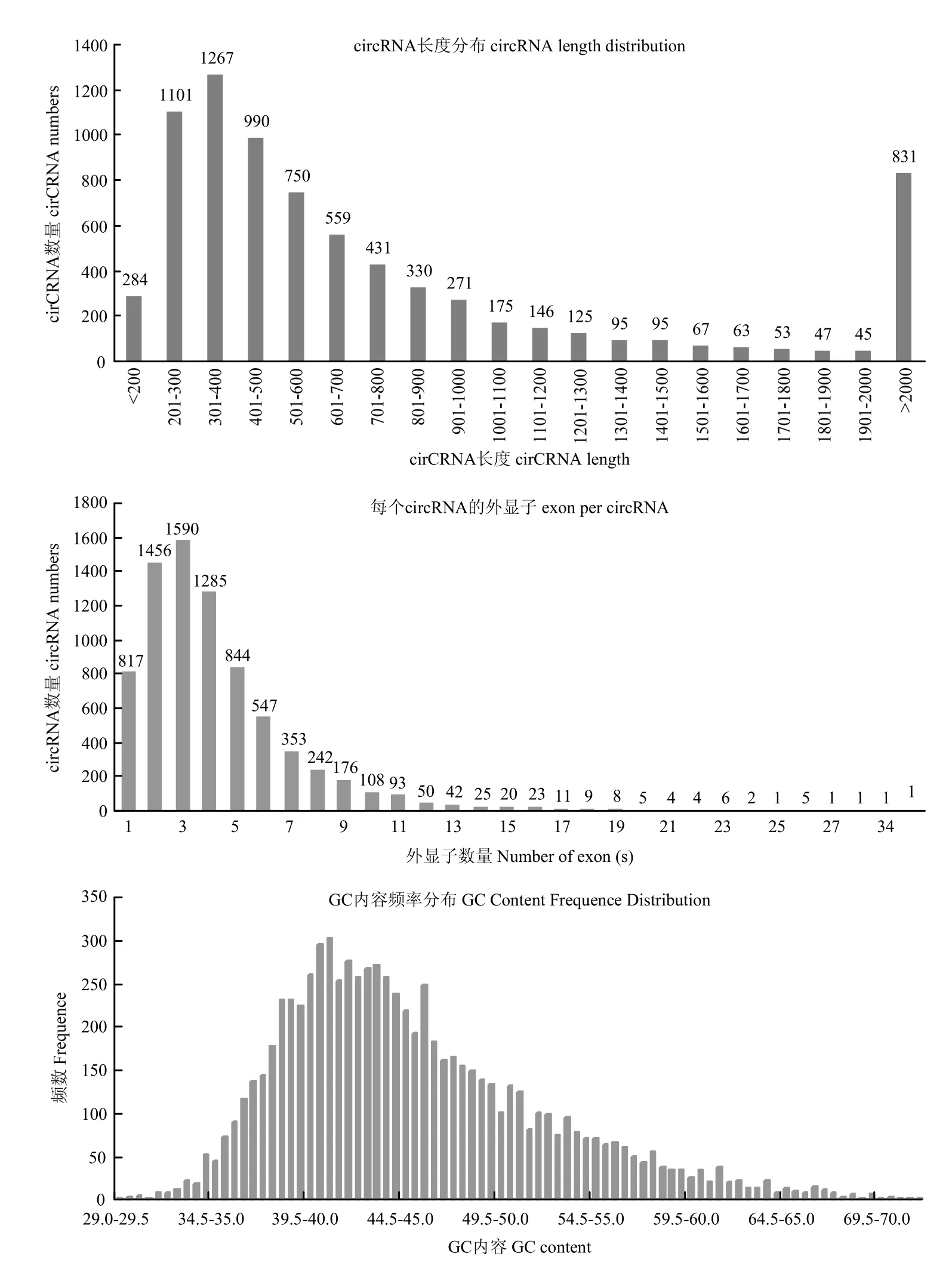
图1 circRNA长度、GC含量、预测circRNA的外显子数目统计Fig. 1 circRNA length, GC content, predicting the number of exons in circRNA

图2 circRNA剪切信号统计Fig. 2 circRNA shear signal statistics

图3 circRNA基因结构分布Fig. 3 circRNA gene structure map
2.2 分析和验证差异转录本
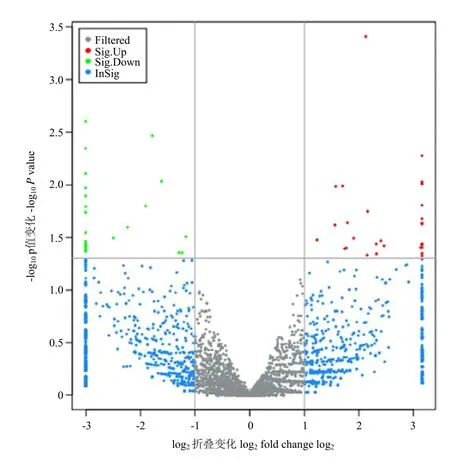
利用FPKM值估计circRNA转录物的表达水平,可以看出circRNA转录本表达水平相对很低(图5)。筛选出31个上调和29个下调的DE circRNA(图6)为了进一步验证 RNA-seq的可靠性,随机选择 6个DE circRNA,用q-PCR验证它们在不腹泻组和腹泻组羔羊体内的相对表达水平,发现与RNA-seq结果一致(图7),表明RNA-seq数据是可靠的。 这些分析还表明,高通量测序具有检测低表达水平(0 图4 circRNA在各条染色体上或scaffold上数目分布Fig. 4 Distribution of circRNA on various chromosomes or on scaffold DE circRNA与GO 数据库进行比对的结果表明,一共有60条circRNA被注释和分类到297个功能亚类中。图8显示了3种分类中DE circRNA数目大于2的GO条目。结果显示,绵羊oxidation-reduction process(GO:0055114)、transport(GO:0006810)、extracellular region(GO:0005576)、focal adhesion(GO:0005925)、extracellular exosome(GO:0070062)、extracellular space(GO:0005615)、zinc ion binding(GO:0008270)等7个功能亚类的circRNA较多,而其余的功能亚类的circRNA分布较少。DE circRNA与KEGG PATHWAY数据库进行比对的结果表明,一共有 60条 circRNA被注释和归类到73个KEGG PATHWAY 中。图8显示了KEGG通路中DE circRNA数目大于2的KEGG条目。结果显示,绵羊Estrogen signaling pathway(path:ko04915)、Protein processing in endoplasmic reticulum(path:ko04141)、Regulation of actin cytoskeleton(path:ko04810)等3个KEGG Pathway的circRNA较多,而其余的KEGG Pathway的circRNA分布较少。 图5 circRNA和mRNA转录本表达水平箱线Fig. 5 circRNA and mRNA transcript expression level box plot 图6 不腹泻和腹泻型羔羊之间差异表达的circRNAsFig. 6 circRNAs differentially expressed between lambs without diarrhea and diarrhea 图7 DE circRNA在不腹泻和腹泻羔羊体内的相对表达水平Fig. 7 Relative expression levels of DE circRNA in lambs without diarrhea and diarrhea 由于circRNA包含多个miRNA结合位点,因此可以使用miRNA靶基因预测的方法鉴定与miRNA结合的 circRNA,笔者使用 Miranda软件来预测与miRNA结合的circRNA以及miRNA的靶基因,根据miRNA靶基因的功能注释来阐明此部分 circRNA的功能,预测详细结果见表2。 由于传统的分子生物学方法对 circRNA数量和丰度的检测能效都非常有限,因此一直以来circRNA被认为是 RNA异常剪接的产物[7]。近年来随着生物信息学的快速发展和高通量测序技术的不断革新,目前通过高通量测序和生物信息学分析已经在真核生物中鉴定了大量的 circRNA[6,22-23],并发现circRNA可能在调控基因表达过程中发挥着重要作用[9,24-25]。研究发现大多数 circRNA包含有 miRNA结合位点,可以作为高效的竞争性内源RNA,有效吸附miRNA从而调控miRNA的靶基因[26-27],其具有以下4种生物学功能:miRNA海绵作用[25-26],作为蛋白质翻译模板[28-29],调控基因转录[30-31]以及通过竞争调控线性RNA的生成[32-33]。然而,迄今为止,关于羔羊腹泻的circRNA,特别是绵羊的研究报道很少。湖羊是一种具有高繁殖力和对湿热气候适应性强的中国特有品种,可以全年在室内饲养。在这项研究中,笔者不仅提供了绵羊腹泻过程中 circRNA的第一个概况,还研究了 circRNA在抗病过程中的可能作用。 表2 circRNA-miRNA-mRNA靶标关系预测Table 2 circRNA-miRNA-mRNA target relationship prediction 图8 circRNA的GO功能注释(上)KEGG PATHWAY富集分析(下)Fig. 8 GORNA PATHWAY enrichment analysis (down) of GO function annotation (up) of circRNA 长期以来,羔羊腹泻对牧场造成了严重的经济损失。在研究中,发现circRNA的表达水平很低。由于circRNA包含多个 miRNA结合位点,因此可以通过Miranda 软件使用miRNA靶基因预测的方法鉴定与6个circRNA结合的5个miRNA,并鉴定出其中4个circRNA的母基因,分别为Btnl 1、GSTM1、NRAMP2、B2M。 Btnl 1是T细胞活化和免疫疾病的关键抑制因子[34]。Btnl 1的作用机制不同于Btnl 2,BtnlA1,它是通过反受体接合直接抑制T细胞活化[34-37]。研究发现Btnl基因可能是肠道炎症的新型重要局部调节因子[38]。GSTM1编码谷胱甘肽-S-转移酶(glutathione-S-transferase,GST)M1酶,参与肺癌各种致癌物质的排毒[39],并且在保护细胞免受氧化应激方面发挥关键作用[40]。NRAMP2是一种金属转运蛋白,在锰缺乏的条件下,NRAMP2参与了高尔基体中Mn的再生,促进植物根系生长[41]。B2M编码主要组织相容性复合体(major histocompatibility complex, MHC)I类分子的β链,并在炎症和肿瘤细胞中上调[42]。 笔者也使用Miranda 软件预测了3个miRNA的靶基因,并且在不腹泻和腹泻组之间显著差异表达,分别为 NEB、UBE3B、ADGRF2、LAMA1、LTF、MGAT5、TLN2、SLC25A29。 NEB基因编码伴肌动蛋白,是细胞骨架基质的巨大蛋白组分,与骨骼肌肌小节中的粗细肌丝共存,NEB基因的突变是杆状体肌病最常见的原因,约占50%[43]。泛素化需要3种酶的连续作用:活化酶(E1),结合酶(E2)和连接酶(E3)。UBE3B是泛素化连接酶(UBE3)成员之一,它和 E2结合酶的独特组合提供了靶向特定蛋白质降解所需的高度底物特异性[44]。ADGRF2是粘附性G蛋白偶联受体家族成员之一,在细胞间与细胞-基质的粘附过程中扮演重要角色[45]。LAMA1突变可能与Poretti-Boltshauser综合征有关,有研究证明 LAMA1缺陷可导致细胞骨架形态的改变[46]。LTF是转铁蛋白家族基因的成员,其蛋白质产物能够启动宿主防御,抵抗广泛的微生物感染,抗原活性[47]。MGAT5编码的蛋白质属于糖基转移酶家族,它是参与调控糖蛋白寡糖生物合成的最重要的酶之一,细胞表面糖蛋白上寡糖的改变引起细胞粘着或迁移行为的显著变化,这种酶的活性增加与侵袭性恶性肿瘤的发展有关[48]。Talins是将粘附分子的整合素家族连接到 F-肌动蛋白的大的接头蛋白,Talin 1对于整联蛋白介导的细胞粘附是必需的,TLN2与Talin 1一样,被认为与独特的跨膜受体结合,在细胞外基质和肌动蛋白细胞骨架之间形成新的连接[49]。SLC25A29编码核编码的线粒体蛋白质,其是溶质运载体家族 25(SLC25)线粒体运载体大家族的成员,SLC25A29的主要生理作用是将碱性氨基酸导入线粒体,进行线粒体蛋白质合成和氨基酸降解[50]。 GO是一种广泛用于研究基因功能关系的生物信息学工具。对60个DE circRNA进行GO分析,结果表明,绵羊 oxidation-reduction process(GO:0055114)、transport(GO:0006810)、extracellular region(GO:0005576)、focal adhesion(GO:0005925)、extracellular exosome(GO: 0070062)、extracellular space(GO:0005615)、zinc ion binding(GO:0008270)等7个功能亚类富集到的circRNA较多。KEGG Pathway分析表明,信号通路如Estrogen signaling pathway(path:ko04915)、Protein processing in endoplasmic reticulum(path:ko04141)、Regulation of actin cytoskeleton(path:ko04810)可能是DE circRNA参与调控的重要途径,相关的 circRNA可能潜在地参与菌毛黏附肠道黏膜的过程。然而,这些通路在抗病过程中的作用很大程度上仍然是未知的。 笔者发现总共60个已知的circRNA在不腹泻组和腹泻组之间显著差异表达,其中31个上调和29个下调。另外,我们确定了两组中共有 1 942个新的circRNA。为了进一步验证RNA-Seq结果,采用q-PCR验证6个已知circRNA的表达水平,结果一致。 通过对不腹泻组和腹泻组羔羊脾脏中circRNA表达谱的研究,了解其在绵羊抗病发生过程中具有调控作用。进一步发现 circRNA的差异表达,预测了与circRNA竞争性结合的miRNA及其靶基因,初步构建circRNA-miRNA-mRNA网络互作模型。 [1] 邓齐文, 许晔琼, 王书奎. 长链非编码RNA 多态性与肿瘤相关性的研究. 医学研究生学报, 2014, 27(3): 303-306.DENG Q W, XU Y Q, WANG S K. Long-chain non-coding RNA polymorphism and tumor correlation study.Journal of Medical Postgraduate, 2014, 27(3): 303-306. (in Chinese) [2] SANGER H L, KLOTZ G, RIESNER D, GROSS H J,KLEISCHMIDT A K. VIROID are single-stranded covalently closed circular RNA molecules existing as highly base-paired rod-like structures.Proceedings of the National Academy of Sciences of the United States of America,1976, 11: 3852-3856. [3] MATSUMOTO Y, FISHEL R, WICKNER R. Circular single-stranded RNA replicon in Saccharomyces cerevisiae.Proceedings of the National Academy of Sciences of the United States of America,1990,87(19): 7628-7632. [4] ARNBERG A C, VAN OMMEN G J B, GRIVELL L A, BRUGGEN E F J V, BORST P. Some yeast mitochondrial RNAs are circular.Cell,1980, 19(2): 313-319. [5] KOS A, DIJKEMA R, AMBERG A C, VAN P H, SCHELLEKENS H.The hepatitis delta (delta) virus possesses a circular RNA.Nature,1986, 323(6088): 558-560. [6] CAPEL B, SWAIN A, NICOLIS S, HACKER A, WALTER M,KOOPMAN P, GOODFELLOW P, LOVELL-badqeBADQE R.Circular transcripts of the testis-determining gene Sry in adult mouse testis.Cell, 1993, 7(5): 1019-1030. [7] COCQUERELLE C, MASCREZ B, HETUIN D, BAILLEUL B.Mis-splicing yields circular RNA molecule.FASEB Journal, 1993,7(1): 155-160. [8] LASDA E, PARKER R. Circular RNAs: diversity of form and function.RNA, 2014, 20(12): 1829-1842. [9] SALZMAN J, GAWAD C, WANG P L, LACAYO N, BRWON P O.Circular RNAs are the predominant transcript isoform from hundreds of human genes in diverse cell types.PLoS One, 2012, 7(2): e30733. [10] ASHWAL-FLUSS R, MEYER M, PAMUDURITI N R, LAVNOV A,BARTOK O. HANMAN M, EVANTAL N, MEMCZAK S, RAJEWSKY N, KADENER S. circRNA biogenesis competes with pre-mRNA splicing.Molecular Cell, 2014, 56(1): 55-66. [11] STARKE S, JOST I, ROSSBACH O, SCHNEIDER T, SCHREINER S. HUNQ L H, BINDIREIF A. Exon circularization requires canonical splice signals.Cell Report, 2015, 10(1): 103-111. [12] 徐兴文. 羊大肠杆菌病防治. 中国畜禽种业, 2017, 4: 129-130.XU X W. Prevention and treatment ofEscherichia Coliin sheep.China Animal and Poultry Industry, 2017, 4: 129-130. (in Chinese) [13] 张文静. 羊大肠杆菌病的防控措施. 畜牧兽医科技信息, 2017, 6:76.ZHANG W J. Prevention and control measures for sheep colibacillosis.Herbivore, 2017, 6: 76. (in Chinese) [14] LEWIS S J, HEATON K W. Stool form as a useful guide to intestinal transit time.Scand J Gastroenterol. 1997, 32: 920-924. [15] P, G, P. P, and R. N, circBase: a database for circular RNAs.Rna-a Publication of the Rna Society, 2014. 20(11): 1666-1670. [16] GAO Y, WANG J, ZHAO F, CIRI: an efficient and unbiased algorithm for de novo circular RNA identification.Genome Biology, 2015. 16: 4. [17] ANDERS S. HUBER W. . Differential expression of RNA-Seq data at the gene level – the DESeq package.European Molecular Biology Laboratory, 2013. [18] ROBINSON, M D, MCCARTHY D J, SMYTH G K. edgeR: a Bioconductor package for differential expression analysis of digital gene expression data.Bioinformatics, 2010, 26(1): 139-140. [19] KANEHISA M, ARAKI M, GOTO S, HATTORI M, HIRAKAWA M,ITOH M, KATAYAMA T, KAWASHIMA S, OKUDA S, TUKIMATSU T,YAMANISHI Y. KEGG for linking genomes to life and the environment.Nucleic Acids Research, 2008, 36: 480-484. [20] JOHN B, ENRIGHT A J, ALEXIE A, THOMAS T, CHRIS S. Human MicroRNA targets.PLoS Biology, 2004. 2(11): e363. [21] ENRIGH A, JOHN B, GAUL U, TUSCHL T, SANDER C, MARKS S.MicroRNA targets in Drosophila.Genome Biology, 2004, 5(1): R1. [22] ABDELMOHSEN K, PANDA A C, DES, GRAMMITIKAKIS I, KIM J, DING J, NOH J H, KIM K M, MATTISON J A, DECABO R,COROSEP M . Circular RNAs in monkey muscle: age-dependent changes.Aging (Albany NY), 2015, 7 (11): 903-910. [23] BURD C E, JECK W R, LIU Y, SANOFF H K, WANG Z.SHARPLESS N E. Expression of linear and novel circular forms of an INK4/ARF-associated Non-coding RNA correlates with atherosclerosis Risk.PLoS Geneicst, 2010, 6 (12): e1001233. [24] SALZMAN J, CHEN R E, OLESN M N, WANG P L, BROWN P O.Cell-type specific features of circular RNA expression.PLoS Genetics,2013, 9 (9): e1003777. [25] MEMCZAK S, JENS M, ELEFSINIOTI A, TORTI F, KRUEGER J,RYBAK A, MAIER L, MACKOWIAK S D, GREGEISEN L H,MUNSCHAUER M, LOERWER A, ZIEBOLD U, LANDTHALER M, KOCKS C, NOBEL F, RAJEWSKY N. Circular RNAs are a large class of animal RNAs with regulatory potency.Nature, 2013, 495(7441): 333-338. [26] HANSEN T B, JENSEN T I, CLAUSEN B H, BRAMSEN J B,FINSEN B, DAMGAARD C K, KJEMS J. Natural RNA circles function as efficient microRNA sponges.Nature, 2013, 495 (7441):384-388. [27] ZHENG Q, BAO C, GUO W, LI S, CHEN J, CHEN B, LUO Y, LYU D, LI Y, SHI G, LIANG L, GU J, HE X, HUANG S. Circular RNA profiling reveals an abundant circHIPK3that regulates cell growth by sponging multiple miRNAs.Nature Communication, 2016, 7: 11215. [28] CHEN C Y, SARMOW P. Initiation of protein synthesis by the eukaryotic translational apparatus on circular RNAs.Science, 1995,268 (5209): 415-417. [29] WANG Y, WANG Z. Efficient backsplicing produces translatable circular mRNAs.RNA, 2015, 21 (2): 172-179. [30] ZHANG Y, ZHANG X O, CHEN T, XIANG J F, XING Y H , YIN Q,ZHU S, YANG S, CHEN L L. Circular intronic long noncoding RNAs.Molecular Cell, 2013, 51 (6): 792-806. [31] LI Z, HUANG C, BAO C, CHEN L, LIN M, WANG X, ZHONG G,YU B, HU W, DAI L, ZHU P, CHANG Z, WU Q, ZHAO Y, JIA Y,XU P, LIU P, SHAN G. Exon-intron circular RNAs regulate transcription in the nucleus.Nature Structural Molecular Biology, 2015, 22 (3):256-264. [32] LASDA E P, ARKER R. Circular RNAs: diversity of form and function.RNA, 2014, 20 (12): 1829-1842. [33] ASHWAL-FLUSS R, MEYER M, PAMUDURTI NR, LVANOV A,BARTOK O, HANAN M, EVANTAL N, MEMCZAK S,RAJEWSKY N, KADENER S. circRNA biogenesis competes with pre-mRNA splicing.Molecular Cell, 2014, 56 (1): 55-66. [34] YAMAZAKI T, GOYA I, GRAF D, CRAIG S, MARTIN-OROZCO N,DONG C. A Butyrophilin Family Member Critically Inhibits T Cell Activation.TheJournal of Immunology, 2010, 185 (10): 5907-5914. [35] NGUYEN T, LIU XK, ZHANG Y, DONG C. BTNL2, a butyrophilin-like molecule that functions to inhibit T cell activation.TheJournal of Immunology, 2006, 176: 7354-7360. [36] ARNETT H A, ESCOBAR S S, GONZALEZSUAREZ E,BUDELSKY A L, STEFFEN L A, BOIABI N, ZHANG M, SIU G,BREWER A W, VINEY J L. BTNL2, a butyrophilin/B7-like molecule,is a negative costimulatory molecule modulated in intestinal inflammation.TheJournal of Immunology, 2007, 178: 1523-1533. [37] SMITH I A, KNEZEVIC B R, AMMANN J U, RHODES D A, AW D,PALMER D B, MATHER I H, TROWSDALE J. BTN1A1, the mammary gland butyrophilin, and BTN2A2 are both inhibitors of T cell activation.TheJournal of Immunology, 2010, 184: 3514-3525. [38] BAS A, SWAMY M, ABELER-DORNER L, WILLIAMS G, PANG D J, BARBEE S D, HAYDAY A C , HAYDAY A C. Butyrophilin-like 1 encodes an enterocyte protein that selectively regulates functional interactions with T lymphocytes.Proceedings of the National Academy of Sciences of the United States of America,2011, 108(11): 4376-4381. [39] SHEPPERD J A, LIPKUS I M, SANDERSON S C, MCBRIDE C M,O’NEILL S C, DOCHERTY S. Testing different communication formats on responses to imagined risk of having versus missing the GSTM1 gene.Journal of Health Communication,2013, 18: 124-137. [40] SINGH R, MANCHANDA P K, KESARWANI P, SRIVASTAVA A,MITTAL R D. Influence of genetic polymorphisms in GSTM1,GSTM3, GSTT1 and GSTP1 on allograft outcome in renal transplant recipients.Clinical Transplantation, 2009, 23: 490-498. [41] SOCHA A L, GUERINOT M L. Mn-euvering manganese: the role of transporter gene family members in manganese uptake and mobilization in plants.Frontiers in Plant Science, 2014, 5: 106. [42] MIYATA T, JADOUL M, KUROKAWA K, YDSC C, VAN . beta-2 microglobulin in renal disease.Journal of the American Society of Nephrology, 1998 (9): 1723-1735. [43] WALLGREN-PETTERSSON C, SSEWRY C A, NONWAK K J,LAING N G, Nemaline myopathies. Semin.Pediatric Neurology,2011, 18 (4): 230-238. [44] KUMAR S, KAO W H. , HOWLEY P M. Physical interaction between specific E2 and Hect E3 enzymes determines functional cooperativity.Journal of Biology. Chemistry, 1997, 272: 13548-13554. [45] HAMANN J, AUGST G, ARAC D, ENGLE F B, FORMSTON C.International union of basic and clinical pharmacology. XCIV.Adhesion G protein-coupled receptors.Pharmacol Reviews, 2015;67(2): 338-367. [46] VILBOUX T, MALICDAN M C, CHANG Y M, GUO J, ZERFAS P M, STEPHEN J, CULLINANE A R, BYANT J, FISHER R, BROOKS B P, ZEIN W M, WIGGS E A, ZELEWSKI C K, PORETTI A,BRYAN M M, VEMULAPALLI M, MULLIKIN J C, KIRBY M,ANDERSON S M, HUIZING M, TORO C, GAHL W A, GUNAYAYQUN M. Cystic cerebellar dysplasia and biallelic LAMA1 mutations: a lamininopathy associated with tics, obsessive compulsive traits and myopia due to cell adhesion and migration defects.Cognitive and Behavioural Genetics, 2016, 53(5): 318-329. [47] FELIPE L O, JUNIOR W F D S, ARAUJO K C, FABRINO D L.Lactoferrin, chitosan and Melaleuca alternifolia-natural products that show promise in candidiasis treatment.Brazilian Journal Microbiology,2017, 49(2): 212-219. [48] HASSANI Z, SALEH A, TURPAULT S, KHIATI S, MORELLE W,VIQNON J, HUQNOT J P, URO-COSTE E, LEQRAND P,DELAFORGE M, LOISEAU S, CLARION L, LECOUVEY M,VOLLE J N, VIRIEUX D, IRAT J L, DUFFAU H, BAKALARA N.Phostine PST3. 1a Targets MGAT5 and inhibits glioblastomainitiating cell invasiveness and proliferation.Molecular Cancer Research, 2017, 15(10): 1376-1387. [49] DEBRAND E, ELJAI Y, SPENCE L, BATE N, PRAEKELT U,PRITCHARD C A, MONKLEY S J, CRITCHLEY D R. Talin 2 is a large and complex gene encoding multiple transcripts and protein isoforms.FEBS Journal, 2009, 276(6): 1610-1628. [50] PORCELLI V, FIERMONTE G, LONGO A, PALMIERI F. The human gene SLC25A29, of solute carrier family 25, encodes a mitochondrial transporter of basic amino acids.Journal of Bioogylogy Chemistry, 2014, 289(19): 13374-13384.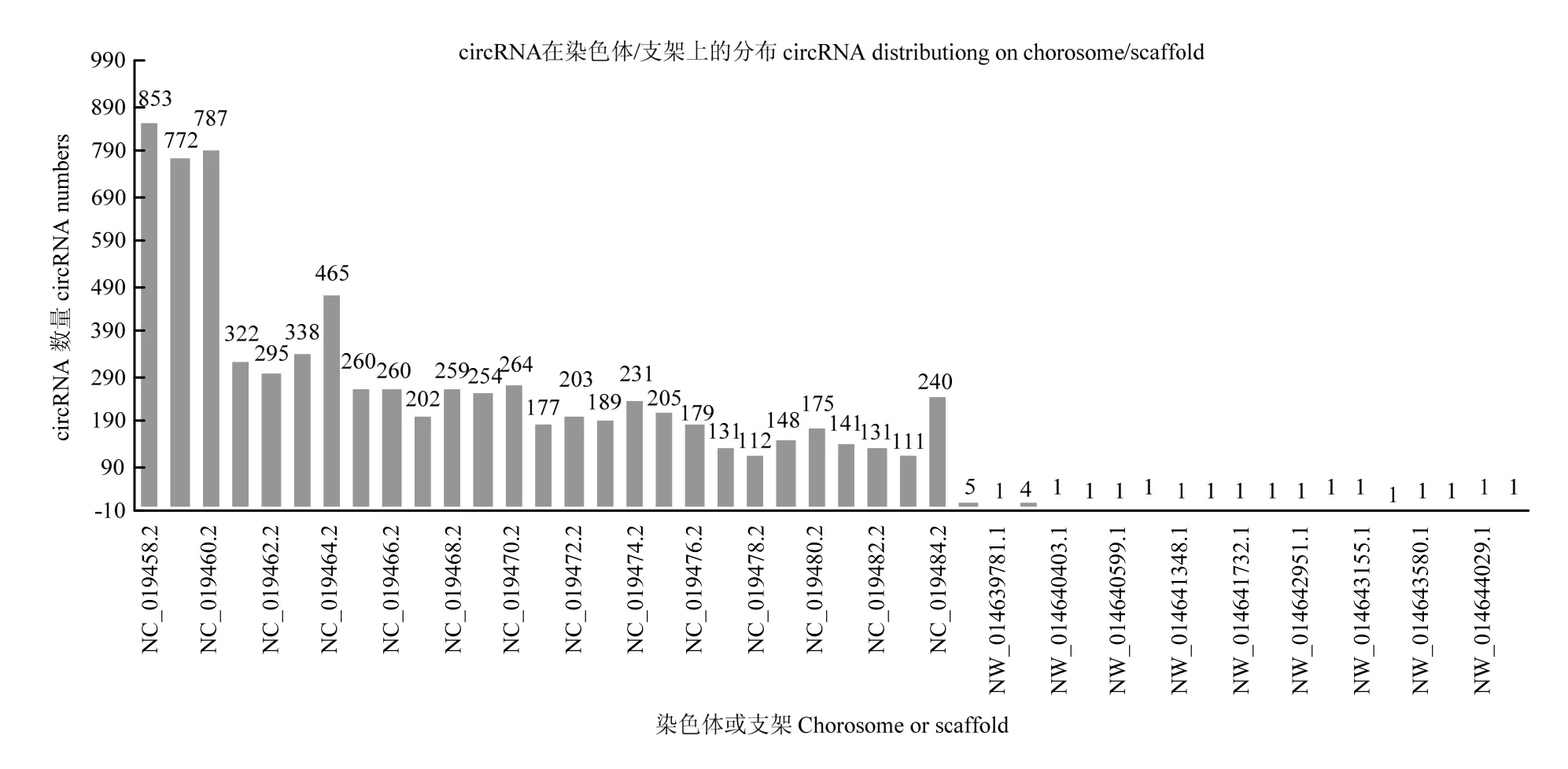
2.3 DE circRNA的GO功能注释和KEGG PATHWAY富集分析


2.4 circRNA-miRNA-mRNA靶标关系预测
3 讨论


4 结论
猜你喜欢
新民周刊(2022年27期)2022-08-01 07:04:49
传染病信息(2021年6期)2021-02-12 01:52:58
意林(儿童绘本)(2019年6期)2019-08-05 01:22:06
小学生必读(低年级版)(2019年12期)2019-04-20 08:39:08
幼儿画刊(2018年7期)2018-07-24 08:26:10
散文诗(2017年22期)2017-06-09 07:55:25
新疆农垦科技(2016年2期)2016-08-21 13:50:24
数学大王·低年级(2016年8期)2016-05-14 10:20:08
生物医学工程学进展(2015年1期)2015-02-28 14:53:42
化学工业与工程(2015年1期)2015-02-10 03:01:41
