
硫代腺苷甲硫氨酸促进番茄百菌清降解的生理机制
2019-04-03 05:58顾超珩闫燕燕魏夕雅史庆华巩彪
中国农业科学 2019年6期
顾超珩,闫燕燕,魏夕雅,史庆华,巩彪
(山东农业大学园艺科学与工程学院/作物生物学国家重点实验室/农业部黄淮海地区园艺作物生物学与种质创新重点实验室,山东泰安 271018)
0 引言
【研究意义】樱桃番茄(Lycopersicon esculentumvar.cerasiforme)是一类具有较高营养价值和独特风味的鲜食番茄。其产业经济效益高,栽培面积大,在我国发展迅速,甚至已经成为一些地区的农业支撑产业。但设施栽培条件下,樱桃番茄易受各种病原菌侵染,引起叶霉病、灰霉病、早疫病、晚疫病和炭疽病等病害。作为生产中广泛使用的广谱性杀菌剂,百菌清(2,4,5,6-tetrachloroisophthalonitrile,chlorothalonil,CHT)常被应用于上述病害的化学防控,其用药安全期为 25 d[1]。但樱桃番茄生产多连续采摘,一旦用药,将会导致后续产品农残超标,影响经济效益和人类健康。因此,研究番茄快速代谢农药的技术及机制,对鲜食番茄安全生产具有重要意义。【前人研究进展】对于植物而言,农药是一类异源有毒物质,施用过多会对植物造成毒害,引起生理、生化代谢异常,导致植株生长、发育受阻,产量、品质下降[2]。而植物在长期进化中形成了一套独特的解毒系统,能够转化、分解对其生长和发育不利的异源化学物质;其解毒过程分为4个阶段,也称为4项代谢[3]:包括(I)细胞色素P450s和过氧化物酶体的氧化作用;(II)谷胱甘肽 S-转移酶(GST)催化的谷胱甘肽共价结合反应;(III)ABC转运蛋白介导的有毒物质区域化隔离;(IV)水解酶参与的降解反应。4项代谢中,前两项代谢负责将有毒的大分子集团破坏,形成毒性低、移动性差的水溶性代谢产物;后两项代谢负责将低毒小分子物质区域化并进一步水解[4]。由此可见,催化氧化反应和谷胱甘肽螯合反应是植物进行农药代谢的关键步骤。前人研究表明,外源添加油菜素内酯[2]、一氧化氮[5]、胡敏酸[6]等均可促进植物农药代谢。笔者研究发现,硫代腺苷甲硫氨酸合成酶(S-adenosylmethionine synthetase,SAMS)是控制硫代腺苷甲硫氨酸(S-adenosylmethionine,SAM)合成的关键酶,可对多种非生物胁迫产生响应[7];且对盐碱胁迫下番茄的活性氧(ROS)和谷胱甘肽代谢具有显著的调控作用[8-9]。此外,SAM自1975年开始在欧洲作为抗忧郁和关节炎的处方药投入使用;1999年,SAM首度以膳食补充剂的身份在美国推广和发售[10]。【本研究切入点】推测 SAM可能参与调控包含农药在内的异源有毒物质降解,但至今该研究领域尚属空白。【拟解决的关键问题】拟通过研究外源 SAM对植物谷胱甘肽解毒途径的影响,阐明SAM对番茄CHT代谢的作用效果及机理,探讨一种可用于鲜食樱桃番茄农药降解的简便方法,为蔬菜安全、绿色生产提供理论依据和技术支撑。
1 材料与方法
试验于2018年3—10月在山东农业大学作物生物学国家重点实验室进行。
1.1 试验设计
供试番茄品种为樱桃番茄‘千禧’。采用常规育苗和栽培技术管理,待果实绿熟期时向果实表面按推荐剂量喷施CHT(75%可湿性粉剂,先正达生物科技有限公司)。2 d后,采用小型喷雾器将清水(Control)、0.1、0.5、1 和 2 µmol·L-1的 SAM 溶液均匀喷施于用药番茄的表面,以刚能完全打湿果实表面作为处理标准,于农药处理后2、5、10和15 d取番茄果实,测定CHT含量,以确定SAM的合理用量。ROS与谷胱甘肽代谢分析采用上述试验材料与培养方法,设置对照(Control)、单独施用CHT处理(CHT)和施用 CHT 并喷施 0.5 µmol·L-1的 SAM 处理(CHT+SAM)共计 3个处理,于农药处理后 2、5、10和15 d后取番茄果实,对ROS及谷胱甘肽代谢相关指标进行测定。
1.2 测定方法
1.2.1 百菌清含量 取番茄果实样品10 g,利用高速分散器磨碎,以80 mL 80℃的石油醚与40 g无水硫酸钠浸提过夜,滤液经真空抽滤后,采用旋转蒸发仪蒸发,以正己烷溶解,定容至5 mL,提取液经固相萃取柱净化吸附,收集洗脱液用于色谱分析。气相色谱检测条件为:GC-ECD(Agilent Santa Clara, CA, USA);色谱柱:30 m×0.32 mm,0.25 µm;载气:氮气;流速:3.3 mL·min-1;进样口温度:250℃;检测器温度:300℃;柱温:以25℃·min-1的速度由80℃升至260℃,最终保持3.8 min[5]。
1.2.2 ROS和丙二醛(MDA)含量 采用磺胺比色法测定超氧阴离子()含量[11];四氯化钛还原法测定过氧化氢(H2O2)含量[12];硫代巴比妥酸法测定MDA含量[13]。
1.2.3 谷胱甘肽代谢酶活性 谷胱甘肽 S-转移酶(GST)活性的测定参照李顺欣等[14]的方法;谷胱甘肽还原酶(GR)活性的测定参照 FOYER和HALLIWELL[15]的方法;脱氢抗坏血酸还原酶(DHAR)活性的测定参照 NAKANO和 ASADA[16]的方法。
1.2.4 谷胱甘肽含量 采用GONG等[17]的方法测定还原型谷胱甘肽(GSH)含量和总谷胱甘肽(GSH+GSSG)含量,计算氧化型谷胱甘肽(GSSG)含量和谷胱甘肽还原氧化比(GSH/GSSG)。
1.3 数据处理
试验处理及指标测定均执行3次生物学重复,数据采用Microsoft Excel 2007和SAS8.0统计软件进行分析,采用最小差异差数法(LSD)进行差异显著性分析(P<0.05)。

图1 不同浓度SAM对番茄果实CHT含量的影响Fig. 1 Effects of different concentrations of SAM on CHT contents of tomato fruits
2 结果
2.1 SAM对CHT的降解效果
图1表明,果实表面喷施CHT后会产生CHT积累,且随着时间的延长,CHT可被番茄果实自行降解,处理后15 d下降至喷药2 d时的30.71%(P<0.05)。与对照相比,外源喷施 0.1 µmol·L-1的 SAM 在 2—10 d内虽能降低番茄果实CHT含量,但差异不显著(P<0.05);至 15 d 时,0.1 µmol·L-1SAM 处理组的 CHT含量才显著低于对照(P<0.05)。与对照相比,外源喷施0.5、1和 2 µmol·L-1的SAM 均可在5 d时显著降低番茄果实CHT含量,其降低效果持续至15 d;但0.5、1和2 µmol·L-1SAM处理组之间的CHT含量差异不显著(P<0.05)。因此,0.5 µmol·L-1的 SAM 可作为番茄 CHT降解的有效处理组用于后续试验,其CHT残留量在处理后5、10和15 d较同时期对照组分别下降了33.72%、52.87 %和63.75 %。
2.2 SAM对ROS代谢的影响
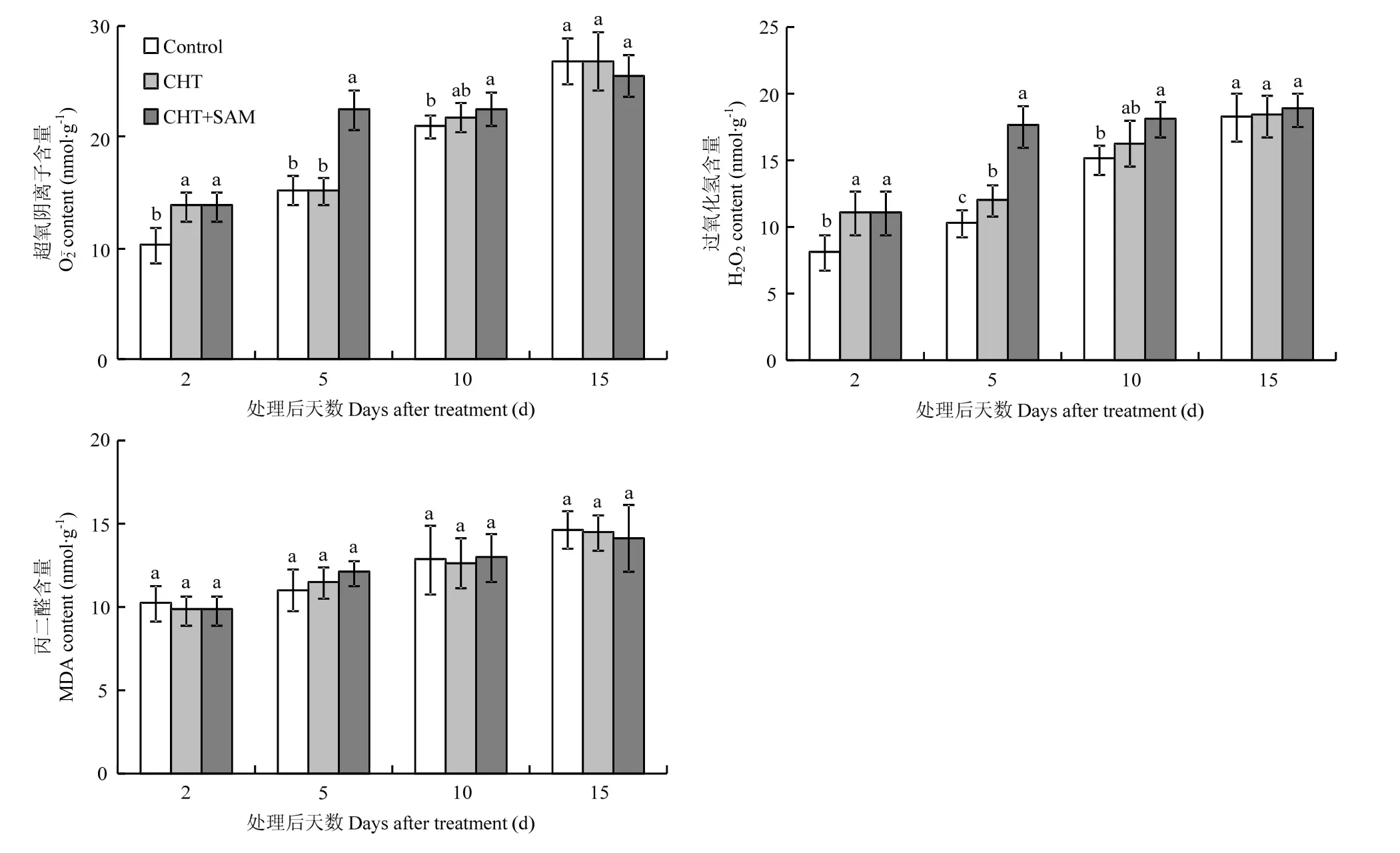
图 2 CHT 和 CHT+SAM 对番茄果实 O2•-、H2O2和 MDA 含量的影响Fig. 2 Effects of CHT and CHT+SAM on contents of, H2O2 and MDA of tomato fruits
鉴于 ROS在参与植物降解农药的氧化过程中发挥重要作用,本研究先就SAM和CHT对ROS含量的影响进行分析(图2)。结果表明,番茄果实的含量于喷施CHT 2 d时显著高于对照,其余处理时段与对照差异不显著(P<0.05);H2O2含量于喷施CHT 2 d和5 d时显著高于对照,其余处理时段与对照差异不显著(P<0.05);而MDA含量在喷施CHT的各处理时段下均与对照差异不显著(P<0.05)。说明喷施CHT可在短期内引起番茄的氧化应激反应,但这种氧化应激并不足以造成细胞损伤。而 CHT+SAM处理下第5天的和 H2O2含量显著高于 CHT处理;CHT+SAM处理下第10天的和 H2O2含量略高于CHT处理,但显著高于对照(P<0.05)。不同时期各处理组间的 MDA含量差异不显著(P<0.05)。说明CHT+SAM处理在没有造成氧化伤害的情况下,较 CHT处理更强烈且更持久的引起细胞内氧化应激反应。
2.3 SAM对谷胱甘肽代谢的影响
谷胱甘肽循环是植物应对胁迫和解毒的关键代谢途径,主要涉及GSH和GSSG之间的转换与利用,而GR和DHAR则是控制GSSG再生成GSH的关键酶。与对照相比,CHT处理可在各时期内增强GR与DHAR的活性;CHT处理下的GR和DHAR活性与对照间的差距均呈现先增后减的趋势,于处理后 5 d达到最大(图3)。说明CHT处理可激活GR和DHAR,提高谷胱甘肽循环效率。CHT+SAM处理下第5天和第10天的GR和DHAR活性显著高于CHT处理,而第15天的GR和DHAR活性显著低于CHT处理(P<0.05);但各时期内CHT+SAM处理下的GR和DHAR活性均显著高于对照(P<0.05)(图3)。说明CHT+SAM处理较CHT处理能更有效的在农药降解早期激活GR和DHAR活性。
图4显示,GSH含量在CHT处理2—10 d时显著高于对照,至15 d时与对照差异不显著(P<0.05);而CHT+SAM处理5—15 d时的GSH含量显著高于对照及 CHT处理(P<0.05)。GSSG含量在 CHT处理2—10 d时显著高于对照,至15 d时与对照差异不显著(P<0.05);但 CHT+SAM 处理 2—5 d时的 GSSG含量与 CHT处理差异不显著,至处理10—15 d时显著低于 CHT处理(P<0.05)。GSH+GSSG在CHT处理2—10 d时显著高于对照,至 15 d时与对照差异不显著(P<0.05);而CHT+SAM处理5 d时的GSH+GSSG显著高于CHT处理,其余处理时段与 CHT处理差异不显著(P<0.05)。GSH/GSSG在CHT处理2—5 d时显著高于对照,至10—15 d时显著低于对照(P<0.05);但CHT+SAM处理5—15 d时的GSH/GSSG均显著高于对照及CHT处理(P<0.05)。说明CHT处理能促进谷胱甘肽的从头合成,提高GSH的还原效率,而SAM主要从提高GSH还原效率层面调控谷胱甘肽代谢以适应番茄的农药降解需求。
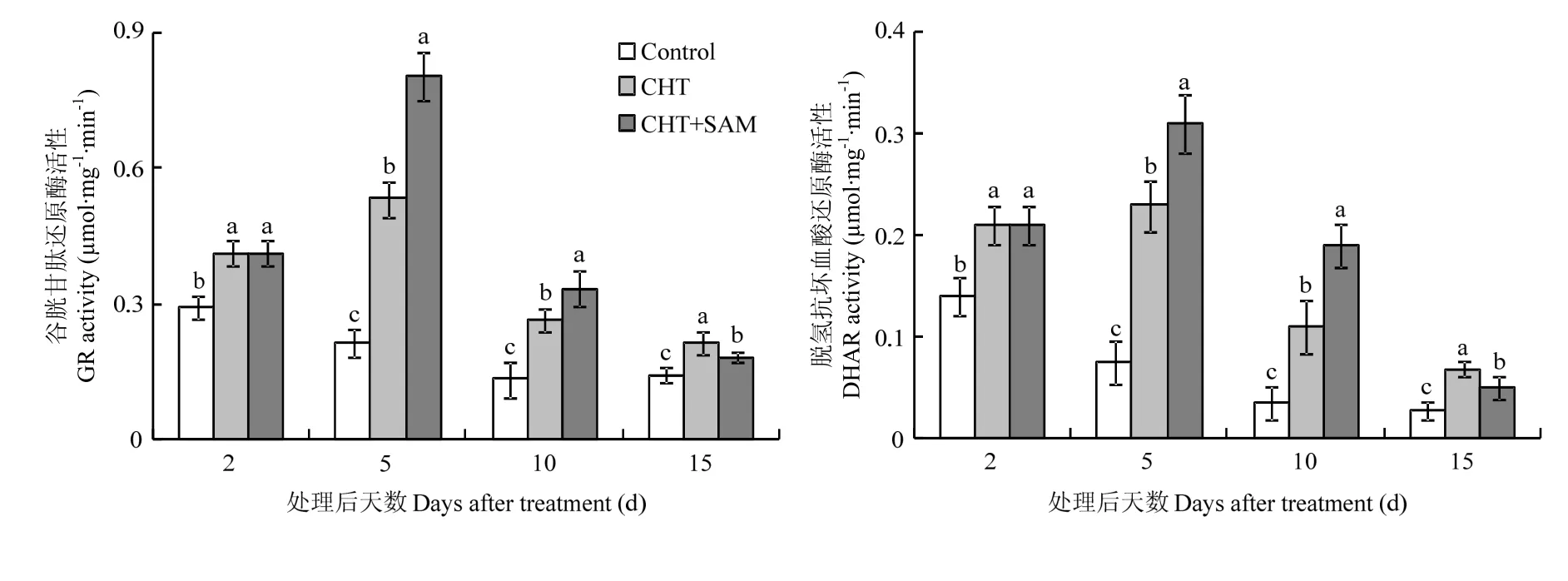
图3 CHT和CHT+SAM对番茄果实GR和DHAR活性的影响Fig. 3 Effects of CHT and CHT+SAM on activities of GR and DHAR of tomato fruits

图4 CHT和CHT+SAM对番茄果实GSH、GSSG、GSH+GSSG和GSH/GSSG的影响Fig. 4 Effects of CHT and CHT+SAM on contents of GSH, GSSG, GSH+GSSG and GSH/GSSG of tomato fruits
2.4 SAM对农药降解关键酶GST活性的影响
GST是生物体针对各类有毒物质进行解毒的关键酶。图5显示,GST活性在CHT处理2—10 d时显著高于对照(P<0.05),至15 d时略高于对照,其在处理后第5天达到峰值。说明GST是CHT代谢响应的关键酶之一。而CHT+SAM处理5—10 d时的GST活性显著高于CHT处理(P<0.05),但CHT+SAM处理 15 d时的 GST活性显著低于 CHT处理(P<0.05),略低于对照。说明CHT+SAM处理较CHT处理能更有效的在农药降解早期激活GST活性。
3 讨论
SAM是蛋氨酸循环的中间产物,其在动物、植物和微生物中的代谢通路相对保守。除了参与蛋氨酸循环外,SAM还参与了转甲基、转氨丙基、转硫等多种生理作用,从而影响谷胱甘肽、烟酰胺、多胺和乙烯等多类信号物质的代谢反应[18]。笔者前期研究发现,SAMS1可受多种逆境诱导,促进SAM合成,以帮助植物应对受胁迫造成的伤害[7-8,19]。本研究表明,外源添加0.5 µmol·L-1的SAM即可显著激活番茄的谷胱甘肽解毒系统,促进农药降解。说明 SAM 并非作为能量代谢物质,而是通过担当信号分子调控番茄的农药解毒过程。

图5 CHT和CHT+SAM对番茄果实GST活性的影响Fig. 5 Effects of CHT and CHT+SAM on GST activity of tomato fruits
植物进行农药代谢的第一步反应是借助细胞内的氧化力对农药分子中不稳定的化学键进行氧化修饰,其主要涉及细胞色素 P450s。作为植物最大的基因家族,目前已有5 100多个细胞色素P450s基因被注释[20]。ZHOU等[2]借助转录组测序技术在番茄中鉴定到2个受CHT诱导的细胞色素基因P450(基因ID:AI776109和 BF112381),但更为深入的遗传验证工作尚未开展。由于细胞色素P450s基因家族过于庞大,科学家至今未能确定植物农药代谢过程中的关键细胞色素基因P450s。但细胞色素P450介导的氧化修饰过程需要细胞内有足够的氧化性物质来提供氧化力,故本研究分析了CHT与CHT+SAM对和H2O2的含量的影响。发现外源添加0.5 µmol·L-1的SAM可在没有造成细胞氧化伤害的情况下引起细胞内氧化应激反应(图2)。SAM是生物体内多胺和乙烯的共同合成前体[18]。笔者前期研究发现,超表达SAMS1[8]或外源添加SAM[9]均可诱导番茄内源多胺和乙烯含量的增加。多胺的积累会激活多胺氧化酶活性,促进多胺降解,释放出反应副产物—H2O2[9];另外,乙烯信号代谢通路往往也涉及 ROS爆发[21],这可能是本研究中外源SAM能进一步诱导ROS的主要原因。综上所述,在对照处理下,番茄细胞的 ROS产生和消除处于平衡态;CHT造成的轻度胁迫可引起ROS产量增加,激活植物的系统性防御反应[22],协同细胞色素 P450促进CHT的氧化降解;外源添加SAM可能从多胺和乙烯代谢通路上协同增强ROS信号介导的防御反应,进一步促进CHT的氧化降解。但无论是CHT处理还是CHT+SAM处理,其细胞内的ROS积累均未超越抗氧化系统的清除能力,故本研究中MDA含量在不同处理间并未产生显著差异。但随着处理时间的延长,、H2O2和MDA含量均呈现上升趋势,这可能是由于番茄在成熟过程中呼吸速率增加,造成的ROS积累和细胞膜系统降解所致。
SAM经过转甲基、水解和转硫作用生成半胱氨酸(Cys),Cys在γ-谷氨酰半胱氨酸合成酶的催化下经γ-谷氨酰半胱氨酸生成 GSH[18]。这可能是本研究中CHT+SAM 处理的 GSH含量和谷胱甘肽总量均高于CHT处理的原因之一。另外,在番茄[8-9]和苜蓿[23]中超表达SAMS1均能通过激活GR和DHAR活性,促进GSSG向GSH转换,增加谷胱甘肽还原氧化比。这与本研究中外源 SAM 对谷胱甘肽代谢的影响相吻合。在植物中,GSH一方面能够减少由于环境和代谢产物胁迫而引发的ROS积累,消除ROS对植物机体的伤害作用;另一方面,GSH又能够在GST的催化作用下与异源有毒物质结合,促进有害物质的区域化降解[24-25]。本研究中GSH含量与和H2O2含量的变化趋势相一致,说明SAM通过诱导ROS积累,激活了GSH的从头合成和再生,增加了谷胱甘肽还原氧化比,使得更多的GSH可以用于GST介导的农药代谢过程。此外,大量研究表明,多胺[26-28]和乙烯[29-30]信号对谷胱甘肽代谢也存在显著的激活作用,这可能是外源SAM促进谷胱甘肽代谢的间接调控途径。
4 结论
硫代腺苷甲硫氨酸作为一种对人类和环境无害的天然生物代谢物质,可通过激活活性氧和谷胱甘肽代谢途径,促进番茄对百菌清的降解。基于ROS和谷胱甘肽在植物解毒途径中功能的保守性,推测硫代腺苷甲硫氨酸可延伸应用于其他作物中的农药代谢降解,但其相关研究仍需后续试验验证。研究结果为硫代腺苷甲硫氨酸在蔬菜绿色生产和农药化控降解技术应用的研究领域奠定了一定的理论基础。
[1] 于安芬, 车宗贤, 王东晖, 李瑞琴, 白滨, 徐瑞, 宋政平. 百菌清在番茄上的自然消解动态及健康风险研究. 西北农业学报, 2016,25(10): 1508-1514.YU A F, CHE Z X, WANG D H, LI R Q, BAI B, XU R, SONG Z P.Study on natural dynamic degradation of chlorothalonil in tomato and its health risks.Acta Agriculturae Boreali-occidentalis Sinica, 2016,25(10): 1508-1514. (in Chinese)
[2] ZHOU Y H, XIA X J, YU G B, WANG J T, WU J X, WANG M M,YANG Y X, SHI K, YU Y L, CHEN Z X, GAN J, YU J Q.Brassinosteroids play a critical role in the regulation of pesticide metabolism in crop plants.Scientific Reports, 2015, 5: 9018.
[3] MUNTANE J. Regulation of drug metabolism and transporters.Current Drug Metabolism, 2009, 10(8): 932-945.
[4] SIMINSZKY B. Plant cytochrome P450-mediated herbicide metabolism.Phytochemistry Reviews, 2006, 5: 445-458.
[5] YIN Y L, ZHOU Y, ZHOU Y H, SHI K, ZHOU J, YU Y L, YU J Q,XIA X J. Interplay between mitogen-activated protein kinase and nitric oxide in brassinosteroid-induced pesticide metabolism inSolanum lycopersicum.Journal of Hazardous Materials, 2016, 316:221-231.
[6] 杜丽亚, 章钢娅, 靳伟. 土壤含水量和胡敏酸对有机氯农药降解的影响. 土壤学报, 2006, 43(2): 158-162.DU Y L, ZHANG G Y, JIN W. Effects of soil water content and humic acid on degradation of organochlorine pesticides.Acta Pedologica Sinica, 2006, 43(2): 158-162. (in Chinese)
[7] 刘鑫, 李晓彤, 荆鑫, 王硕硕, 巩彪, 魏珉, 史庆华. S-腺苷甲硫氨酸对黄瓜断根扦插苗生长及生理代谢的影响. 园艺学报, 2018,45(8): 1513-1522.LIU X, LI X T, JING X, WANG S S, GONG B, WEI M, SHI Q H.Effect of S-adenosylmethionine on growth and physiological metabolism of cucumber cutting seedlings.Acta Horticulturae Sinica,2018, 45(8): 1513-1522. (in Chinese)
[8] GONG B, LI X, VANDENLANGENBERG K M, WEN D, SUN S S,WEI M, LI Y, YANG F J, SHI Q H, WANG X F. Overexpression ofS-adenosyl-L-methionine synthetase increased tomato tolerance to alkali stress through polyamine metabolism.Plant Biotechnology Journal, 2014, 12(6): 694-708.
[9] GONG B, WANG X F, WEI M, YANG F J, LI Y, SHI Q H.Overexpression ofS-adenosylmethionine synthetase 1enhances tomato callus tolerance to alkali stress through polyamine and hydrogen peroxide cross-linked networks.Plant Cell, Tissus and Organ Culture,2016, 124: 377-391.
[10] CHEN Y W, TAN T W. EnhancedS-adenosylmethionine production by increasing ATP levels in baker's yeast (Saccharomyces cerevisiae).Journal of Agricultural and Food Chemistry, 2018, 66(20):5200-5209.
[11] 何建社, 张利, 刘千里, 朱欣伟, 刘兴良, 冯秋红, 罗达, 史作民.岷江干旱河谷区典型灌木对干旱胁迫的生理生化响应. 生态学报,2018, 38(7): 2362-2371.HE J S, ZHANG L, LIU Q L, ZHU X W, LIU X L, FENG Q H, LUO D, SHI Z M. Physiological and biochemical response of typical shrubs to drought stress in the Minjiang River dry valley.Acta Ecologica Sinica, 2018, 38(7): 2362-2371. (in Chinese)
[12] 孙莎莎, 巩彪, 温丹, 王秀峰, 魏珉, 杨凤娟, 李岩, 史庆华. 对羟基苯甲酸胁迫下褪黑素对黄瓜胚根生理生化特性的影响. 应用生态学报, 2016, 27(3): 897-903.SUN S S, GONG B, WEN D, WANG X F, WEI M, YANG F J, LI Y,SHI Q H. Effect of exogenous melatonin on physiological and biochemical characteristics of cucumber radicles under p-hydroxybenzoic acid.Chinese Journal of Applied Ecology, 2016, 27(3): 897-903. (in Chinese)
[13] GONG B, LI X, BLOSZIES S, WEN D, SUN S S, WEI M, LI Y,YANG F J, SHI Q H, WANG X F. Sodic alkaline stress mitigation by interaction of nitric oxide and polyamines involves antioxidants and physiological strategies inSolanum lycopersicum.Free Radical Biology and Medicine, 2014, 71: 36-48.
[14] 李顺欣, 郅军锐, 杨广明, 岳文波, 叶茂. 外源茉莉酸诱导的菜豆叶片生化抗性及其对西花蓟马体内保护酶和解毒酶活性的影响.应用生态学报, 2017, 28(9): 2975-2983.LI S X, ZHI J R, YANG G M, YUE W B, YE M. Resistance of bean leaves induced by exogenous jasmonic acid and its effects on activities of protective and detoxification enzymes inFrankliniella occidentalis.Chinese Journal of Applied Ecology, 2017, 28(9):2975-2983. (in Chinese)
[15] FOYER C H, HALLIWELL B. The presence of glutathione and glutathione reductase in chloroplasts: A proposed role in ascorbic acid metabolism.Planta, 1976, 133: 21-25.
[16] NAKANO Y, ASADA K. Hydrogen peroxide scanvenged by ascorbate-specific peroxidase in spinach chloroplast.Plant and Cell Physiology, 1981, 22: 867-880.
[17] GONG B, WEN D, BLOSZIES S, LI X, WEI M, YANG F J, SHI Q H,WANG X F. Comparative effects of NaCl and NaHCO3stresses on respiratory metabolism, antioxidant system, nutritional status, and organic acid metabolism in tomato roots.Acta Physiologiae Plantarum, 2014, 36: 2167-2181.
[18] 柴兴苹, 张玉秀, 谭金娟, 冯珊珊, 柴团耀. Zn胁迫下小麦S-腺苷甲硫氨酸代谢途径关键基因表达模式分析. 植物生理学报, 2013,49(4): 375-384.CHAI X P, ZHANG Y X, TAN J J, FENG S S, CHAI T Y. Analysis of expression patterns of genes participated inS-adenosylmethionine(SAM) metabolic pathway in wheat under Zn stress.Plant PhysiologyJournal, 2013, 49(4): 375-384. (in Chinese)
[19] EZAKI B, HIGASHI A, NANBA N, NISHIUCHI T. AnS-adenosyl methionine synthetase(SAMS) gene fromAndropogon virginicusL.confers aluminum stress tolerance and facilitates epigenetic gene regulation inArabidopsis thaliana.Frontiers in Plant Science, 2016, 7:1627.
[20] NELSON D, WERCK-REICHHART D A. P450-centric view of plant evolution.Plant Journal, 2011, 66(1): 194-211.
[21] LV B S, TIAN H Y, ZHANG F, LIU J J, LU S C, BAI M Y, LI C Y,DING Z J. Brassinosteroids regulate root growth by controlling reactive oxygen species homeostasis and dual effect on ethylene synthesis inArabidopsis.PLoS Genetics, 2018, 14(1): e1007144.
[22] NOCTOR G, FOYER C H. Intracellular redox compartmentation and ROS-related communication in regulation and signaling.Plant Physiology, 2016, 171(3): 1581-1592.
[23] GUO Z F, TAN J L, ZHUO C L, WANG C Y, XIANG B, WANG Z Y.Abscisic acid, H2O2and nitric oxide interactions mediated coldinducedS-adenosylmethionine synthetase inMedicago sativasubsp.falcatathat confers cold tolerance through up-regulating polyamine oxidation.Plant Biotechnology Journal, 2014, 12: 601-612.
[24] 丁顺华, 陈珊, 卢从明. 植物叶绿体谷胱甘肽还原酶的功能研究进展. 植物生理学报, 2016, 52(11): 1703-1709.DING S H, CHEN S, LU C M. Research progress on functions of glutathione reductase in chloroplasts of plants.Plant Physiology Journal, 2016, 52(11): 1703-1709. (in Chinese)
[25] 谭维嘉, 梁革梅, 郭予元, 王武刚. B.t杀虫剂预处理缓解棉铃虫对化学农药抗性及机理的研究. 中国农业科学, 1997, 30(5): 13-19.TAN J W, LIANG G M, GUO Y Y, WANG W G. Study on the mechanism alleviation of cotton bollworm to insecticides by B.t treatment.Scientia Agricultura Sinica, 1997, 30(5): 13-19. (in Chinese)
[26] KWAK M K, LEE M H, PARK S J, SHIN S M, LIU R, KANG S O.Polyamines regulate cell growth and cellular methylglyoxal in high-glucose medium independently of intracellular glutathione.Febs Letters, 2016, 590(6): 739-749.
[27] DIAO Q N, SONG Y J, QI H Y. Exogenous spermidine enhances chilling tolerance of tomato (Solanum lycopersicumL.) seedlings via involvement in polyamines metabolism and physiological parameter levels.Acta Physiologiae Plantarum, 2015, 37(11): 230.
[28] AGRAWAL S B, AGRAWAL M, LEE E H, KRAMER G F, PILLAI P.Changes in polyamine and glutathione contents of a green alga,Chlorogonium elongatum(Dang) France exposed to mercury.Environmental & Experimental Botany, 1992, 32(2): 145-151.
[29] 化文平, 刘文超, 王喆之, 李翠芹. 干涉丹参SmORA1对植物抗病和丹参酮类次生代谢的影响. 中国农业科学, 2016, 49(3): 491-502.HUA W P, LIU W C, WANG Z Z, LI C Q. Effect of RNAi ofSmORA1on disease resistance and tanshinones secondary metabolism inSalvia miltiorrhiza.Scientia Agricultura Sinica, 2016, 49(3):491-502. (in Chinese)
[30] 周琦, 郑幸果, 何辉煌, 康杰, 肖海华. 植物 S-腺苷甲硫氨酸合成酶的新功能展望. 生命的化学, 2017(4): 57-63.ZHOU Q, ZHENG X G, HE H H, KANG J, XIAO H H. Progresses in new functions of plantS-adenosylmethionine synthetase.Chemistry of Life, 2017(4): 57-63. (in Chinese)
猜你喜欢
舰船科学技术(2022年11期)2022-07-15
今日农业(2021年21期)2022-01-12
中国土壤与肥料(2021年5期)2021-12-02
农业资源与环境学报(2021年5期)2021-10-06
现代养生·上半月(2021年10期)2021-09-24
今日农业(2020年23期)2020-12-15
中国药剂学杂志(网络版)(2019年4期)2019-07-17
水产科学(2019年3期)2019-05-30
分析化学(2018年4期)2018-11-02
作文大王·笑话大王(2018年12期)2018-03-23
