
生物炭配施氮素对陆地棉盛花期根系形态与构型的影响
2019-04-01 12:53徐万里唐光木孙宁川蒲胜海耿增超
农业机械学报 2019年3期
冯 雷 徐万里 唐光木 孙宁川 蒲胜海 耿增超
(1.西北农林科技大学资源环境学院, 陕西杨凌 712100; 2.新疆农业科学院土壤肥料与农业节水研究所, 乌鲁木齐 830092)
0 引言
优化根系是提高作物产量的最可行方式之一[1],却较少被关注[2-3]。通过生物炭耦合作物根际生物地球化学过程[4],增强或减弱反应强度[5],从而减缓资源(水和养分)限制作用并降低机械阻力[6-7],进而影响植物生理生化反应[8],这一过程被认为可以较为全面解释生物炭-土壤-作物连续体之间相互作用机制[9],但关于核心过程中生物炭如何影响作物根系并改变根系不同水平特征的研究却仍显不足[10]。
生物炭对作物根系不同水平(形态、构型、信号传递)的影响与作物生产力、源库物质分配等密切相关。研究表明:4.5~9 t/hm2生物炭促进了主根生长和根系分枝密度增加,但抑制了二级根生长,小麦比根长增加33.57%[11];文献[12]也曾报道,添加4 t/hm2生物炭,配施氮156 kg/hm2、磷56 kg/hm2和钾28 kg/hm2,玉米根系生物量增加近两倍,根系夹角、比表面积、分枝和细根分别增加(14±11)%、(54±14)%和(70±56)%。SPOKAS等[13]报道生长素可能是影响生物炭修复土壤与增产的关键。另据报道,土壤添加生物炭,植物适应性会从根系形态以及生理等方面做出响应[14]。由此说明,不同物种植物、不同级别根系对生物炭响应规律具有一定差异,且存在多样化的适应资源变化机制。
花铃期中盛花期是棉花根系形态、构型及品质形成调控的关键时期,氮素供给是其主要影响因素之一。一般认为,氮素通过影响作物侧根伸长、侧根细胞分裂、L-谷氨酸信号传导,调控作物根系形态生长和发育,其中,低剂量氮素会刺激作物根系进一步生长,其吸收面积、比根长、根长比例等特征均呈显著增加趋势[15];常规施氮即最适施氮虽然可维持作物生长生活史,但吸收氮素根系比例却显著降低,一定程度上损失了氮资源,产生潜在水体污染[16]。对于棉花而言,最适氮素供给量在240~320 kg/hm2,其完成生活史后土壤中仍残存较多氮素[17],而应用生物炭可以较好解决上述问题。报道称,中低剂量生物炭复配氮素(1.5%~3.3%,质量比)普遍对植物根系生长产生了积极影响[18],并显著提高作物氮素利用效率[19]和增加玉米产量[20]、油麦菜产量[21],其中,根系体积、根长、活跃吸收面积等形态指标明显增加,但也有报道称生物炭降低了根系总量[22]。虽然,生物炭、氮素以及生物炭配施氮素对于作物根系的研究已分别报道,但关于生物炭配施氮素对作物根系构型及不同土层各根级根系响应[23]规律仍存在分歧。
灰漠土是新疆主要耕作土壤类型之一,2017年种植了全国棉花播种总面积60%以上的棉花,丰富的棉花秸秆资源为棉秆炭产学研提供了有利条件。目前,关于生物炭、氮素与棉花根系生长发育已分别有报道:马云艳[24]曾研究指出,棉花生长21周时,25、100 g/kg生物炭(质量比)处理分别使棉花根系鲜质量和须根数分别较对照增加40.6%和37.9%;于莎等[25]证明,根系氮素代谢能力下降的原因主要是谷氨酰胺合酶/谷氨酸合酶循环介导的氮素同化能力下降,其次为硝酸还原酶介导的氮素还原能力下降。然而,目前尚未见生物炭与氮素协同效应对棉花关键生育期根系形态及构型的研究。本文通过设置生物炭和氮素配施,采用广义加性模型(GMA)和冗余分析(RDA),探究二者复配对棉花关键生育期-盛花期根系构型特征的影响,以期阐明生物炭-土壤-棉花连续体相互作用机理,为新疆灰漠土棉花种植体系可持续发展提供科学依据。
1 材料与方法
1.1 试验区概况
试验区位于新疆维吾尔自治区乌鲁木齐市新市区安宁渠镇“中国国家灰漠土肥力与肥料效益野外科学观测台站”,北纬43°95′26″,东经87°46′45″,海拔680~920 m,年均气温5~7℃,年降水量180~250 mm,年蒸发量1 600~2 200 mm,属干旱半干旱荒漠气候。灰漠土土壤性质为pH值8.35、电导率0.22 mS/cm、有机碳质量比6.91 g/kg,全氮、全磷和全钾质量比分别为0.47、0.59、16.35 g/kg。
1.2 试验设计
棉花品种为新陆早49号(GossypiumhirsutumL.49)。播种时间2018年4月24日。试验采用完全随机设计,设计5种处理,3次重复,共计15个小区。小区面积33 m2(7.5 m×4.4 m)。棉花采用1.6 m宽膜滴灌栽培模式,一膜4行,膜内宽行距50 cm,窄行距20 cm,接行60 cm,株距10 cm(20+50+20+50+20+60)×10 cm。处理分别为N0(常规施氮,施氮量0.3 t/hm2),N0+Bc1%(常规施氮0.3 t/hm2+棉杆炭质量为耕层土质量1%),Nl+Bc1%(低氮量0.21 t/hm2+棉杆炭质量为耕层土质量1%),Nl+Bc2%(低氮量0.21 t/hm2+棉杆炭质量为耕层土质量2%),N0+Bc4%(常规施氮0.3 t/hm2+棉杆炭质量为耕层土质量4%)。常规施氮量0.3 t/hm2,低氮量0.21 t/hm2,氮肥施用方式均为基施40%,追施60%。60%追肥分6次随水追施,按棉花需肥规律施用比例分别为15%、10%、10%、10%、10%、5%。所有处理的磷钾肥作为基肥一次性施入,磷肥施入量为138 kg/hm2(P2O5),钾肥施入量为60 kg/hm2(K2O)。
播种后22 d,苗期滴灌1次,水总定额300 m3/hm2;蕾期滴灌2次,水总定额750 m3/hm2;花铃期7月12日—8月27日,每9天滴灌1次,总定额2 250 m3/hm2;成熟期滴灌1次,水总定额360 m3/hm2。
1.3 观测内容
于2018年7月14日棉花盛花期采集样品,掘根法采集0~10 cm和大于10 cm根系。
1.4 指标及测定方法
根系总长度(L,cm)和平均直径(D,mm):采用根系扫描和图像分析法测定。具体方法:收集洗净根系,将多级侧根及须根分开,尽量使其不与上一级根系交织。利用LA-S系列植物图像分析仪扫描获取根系扫描图像,后用Win RHIZO图像分析系统(Win RHIZO Pro2004b,version 5.0,加拿大)进行分析,获得根系总长度和平均根茎。
根系长度(Ln,cm):采用直尺法测定30~50根排放在玻璃板上的根系长度,求和。
根系干质量(M,g):60℃干燥24 h后,采用干燥法测定。
土壤体积(V,cm3):采用直尺法测定根系分布剖面体积。
1.5 指标计算
比根长、根生物量密度、根长密度、根系长度比例计算公式为
S=0.01L/M
G=M/(V×10-6)
R=L/V
B=Ln/L×100%
式中G——根生物量密度,g/m3
R——根长密度,cm/cm3
B——根系长度比例,%
其中,n∈(k,j,h),k表示极细根(直径0~0.1 mm),j表示细根(直径大于等于0.1 mm小于1 mm),h表示中等根(直径大于等于1 mm小于2 mm),共3种根系总根长[26]。
1.6 统计分析方法
采用GMA分析土壤深度、生物炭施用方式单独以及交互作用对根节间距影响。用协方差解释几者间相关性。进行GMA分析时,残差用正态性检验(Shapiro-Wilk’s test,p>0.05),若数据不符合正态分布,则进行对数或平方根转换。分析生物炭施用方式和根节间距因果关系,用Turkey HSD检验(p<0.05)。根系特征5次重复。
文献[1]表明非线性模拟更符合根系生长规律,能够相对准确反映统计指标的变化趋势,因此本文选择对数进行模拟分枝角与细根平均长度。首先,通过SPSS 19.0探索分析剔除数据中测定值与平均值偏差超过两倍标准差的测定值。然后,将所有样本的根系分枝角10°等分,每10°求取算术平均值,以降低数据本身对趋势拟合的干扰。最后,通过对数非线性模型模拟根系分枝角影响棉花细根生长趋势,其中分枝角和细根根长均以算术平均值(n=13)作为拟合指标。
运用Canoco 4.5软件对浅层根长密度和底层根长密度与环境因子进行RDA分析。利用软件OriginPro 8.5作图。
2 结果与分析
2.1 生物炭与氮素交互作用对棉花根系直径和长度的影响
图1a表明,根系平均直径存在一定差异,其中N0处理根系平均直径为5.57 μm,显著低于Nl+Bc1%和N0+Bc1%(p<0.05),相较二者分别减小1.48 μm和2.53 μm,与Nl+Bc2%和N0+Bc4%没有显著差异(p>0.05)。这与HONG等[27]研究结果不同,其认为氮素剂量差异显著影响平均根系直径,本次试验结果可能是Bc1%增强了低剂量氮素对根系生长的促进作用,而高剂量的生物炭可能抑制了根系中硝酸还原酶活性和根系细胞分裂。Nl+Bc1%处理根生物量密度是N0处理2.74倍,达到7 239.52 g/m3。Nl+Bc2%、N0+Bc4%和N0+Bc1%处理根生物量密度分别是N0处理的1.20、0.66、0.60倍。施用1%生物炭而施氮量不同,即施氮量0.21、0.3 t/hm2,前者相较后者棉花根生物量密度增加4.42倍,如图1b所示。这是因为低剂量生物炭包含的少量激素类物质促进了植物根系发育[28]。
图1d表明,生物炭也改变了细根的平均根长,N0+Bc1%处理细根平均根长与Nl+Bc2%处理没有显著差异(p>0.05),显著高于N0+Bc4%、Nl+Bc1%、N0(p<0.05)。其中,N0+Bc1%、Nl+Bc2%处理细根平均根长分别是N0处理的1.66倍和1.53倍,从4.77 cm增加到7.93 cm和7.28 cm。Nl+Bc1%细根平均根长减小1/10,降至4.33 cm。这与RAZAP等[29]报道一致,可能是4%生物炭增强异三聚体G蛋白[30]和谷氨酸转运蛋白[31]反馈调节,也可能影响了根系细胞壁结构[8],因而综合表现为4%生物炭与N配施在一定程度促进了根系伸长。
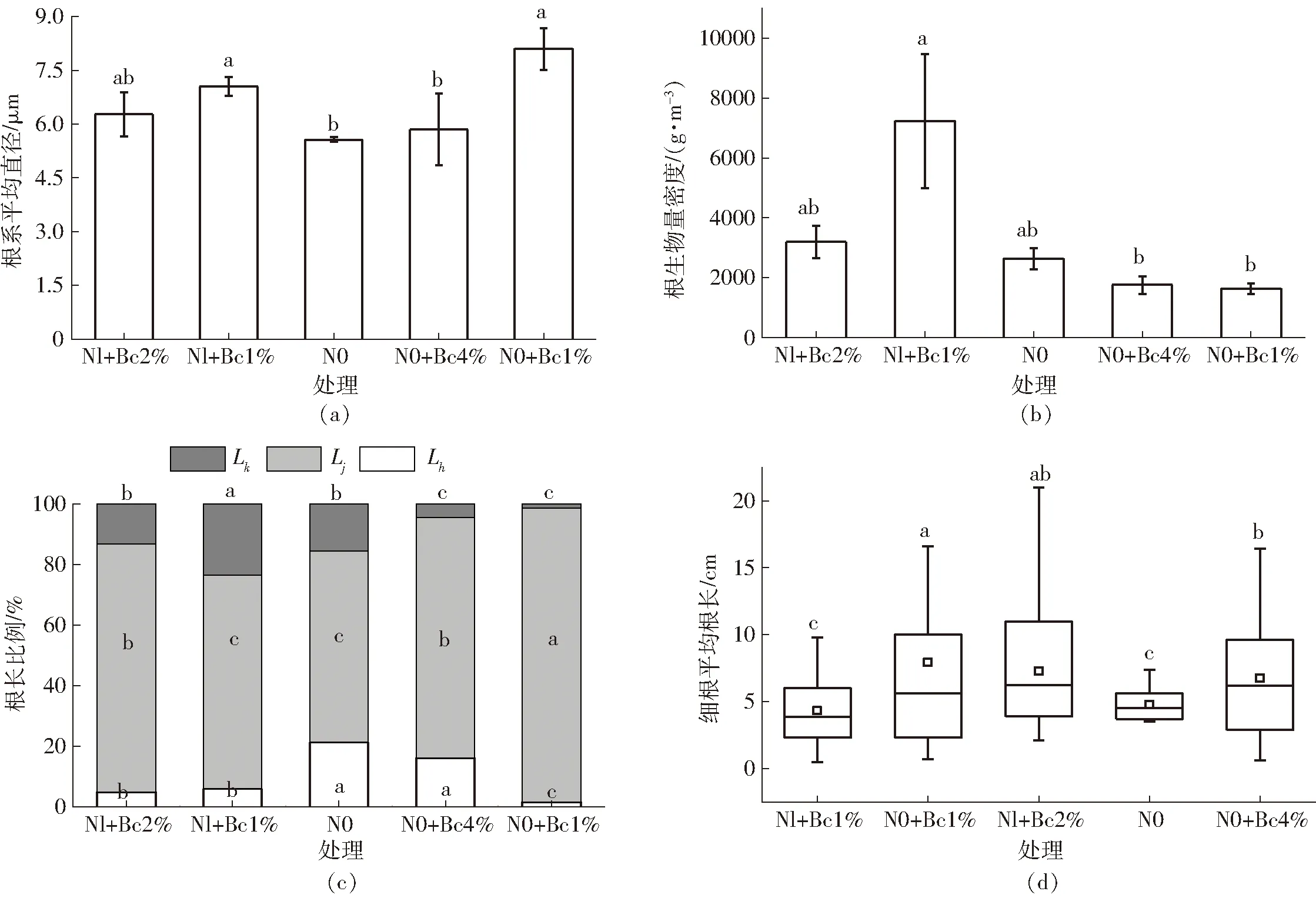
图1 生物炭与氮素交互作用对根系平均直径、根生物量密度、根长比例、细根平均根长的影响Fig.1 Effects of biochar interactionwith nitrogen on average root diameter, root biomass density, root length ratio and root length
2.2 生物炭与氮素交互作用对棉花比根长和根长密度的影响
棉花浅层(0~10 cm)和深层(大于10 cm)根系比根长呈相似变化规律。Nl+Bc2%、N0和N0+Bc1%处理比根长显著大于Nl+Bc1%和N0+Bc4%(p<0.05)。浅层根系比根长前三者间没有显著差异,后两者之间也未显示显著差异(p>0.05)。深层根系Nl+Bc2%比根长显著小于N0+Bc1%(p<0.05),N0与两者没有显著差异(p>0.05),如图2所示。这与李瑞霞等[11]研究结果类似,即中低剂量生物炭增加了作物比根长。一方面,中低剂量生物炭能够较大限度增加植物根系细胞膜载体活性[8],刺激生长素类物质合成与转运[13],而高剂量可能抑制激素类转运过程;另一方面,中低剂量生物炭也可能增强土壤介质氮素扩散系数[32]。

图2 生物炭与氮素交互作用下不同土层深度棉花比根长比较Fig.2 Comparison of specific root length of cotton in different soil layers under biochar interaction with nitrogen

图3 生物炭与氮素交互作用下棉花根长密度变化Fig.3 Change of root length density of cotton under biochar interaction with nitrogen
0~10 cm土层棉花根长密度显著高于大于10 cm土层。Nl+Bc2%的0~10 cm和大于10 cm根长密度具有极显著差异,且显著高于其他处理浅层根长密度(p<0.05),分别是Nl+Bc1%、N0、N0+Bc4%和N0+Bc1%的1.96、2.30、1.54、2.42倍。N0+Bc4%的大于10 cm根长密度显著高于其他处理,Nl+Bc1%、N0+Bc4%和N0+Bc1%之间没有显著差异(图3)。这是因为浅层土壤施入中等剂量生物炭可保存较多养分和水分[33],根长密度显著增加便可获取这部分资源,从而有利于植株进一步生长;深层土壤中,根系倾向分布于中等紧实度的土壤[34],而高剂量生物炭能够降低土壤阻力使根系获得优良生长环境,所以该层根长密度明显增大。
2.3 生物炭对棉花根分枝角和节间距的影响
生物炭未明显改变0~10 cm土层极细根分枝角而显著改变棉花细根分枝角。其中N0+Bc4%处理0~10 cm土层根系分枝角为88°41′,Nl+Bc2%、Nl+Bc1%、N0和N0+Bc1% 分枝角分别为80°21′、80°40′、83°34′和83°04′,平均为83°17′,这说明高剂量的生物炭抑制根系垂直生长,如图4所示。对于大于10 cm土层棉花细根而言,生物炭有利于细根水平延伸,其中Nl+Bc2%分枝角最大,为101°48′且更倾向于向浅层生长,这种表现可能是根系对资源异质性作出的响应。生物炭活化了土壤固定养分的有效性并且阻碍了氮素向深层淋失进而促进植物根系伸展[4],ROYCHOUDHRY等[34]进一步研究指出氮素亏缺引起的硝酸根不足能够增大根系向地性夹角。FORDE[31]也曾在植物氮素信号传递途径指出,根系构型主要受到主侧根之间氮的相互作用调控,侧根系氮素含量增加,主侧根分枝角则可能变大。拟南芥中AtDRO1过表达近来也被证实可以增加其向地性夹角[35]。
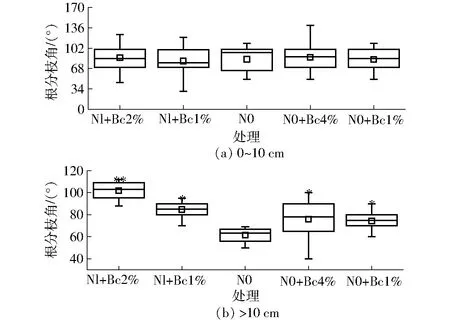
图4 生物炭与氮素交互作用对不同土层根系分枝角的影响Fig.4 Effect of biochar and nitrogen on branching angle of root systems in different soil layers
由图5(图中α、β、η、γ、θ分别表示不同处理根系分枝角,1、2、3、4表示与来水方向夹角(90°、180°、270°、360°))可知,α1、β1、η1、γ1、θ1根节长度相近,约为0.76 cm。不同处理根节长度的差异主要表现在α3、β3、η3、γ3、θ3,分别为1.24、0.99、1.13、4.00、1.60 cm。这种差异可能是由不同土层水分及养分扩散速率所决定。

图5 生物炭施用方式、根分枝角交互作用对根节长度的影响Fig.5 Effects of interaction between application methods of biochar and branching angle on root space
分枝角与细根平均根长呈极显著负相关(R2=0.82,p≤6.93×10-6),符合y=-52.24×ln(0.20lnx)(图6)。这与叶子奇等[36]研究结果一致,其研究认为垂直分枝角越小则土壤相对疏松,细根则可以累积相对较多物质支撑根系伸展。郑慧玲等[37]也曾报道根系分枝角与根长负相关,正是由于植物对资源配置的权衡所导致。

图6 着生于主根的细根平均根长与根分枝角非线性模拟Fig.6 Nonlinear simulation between fine root length and branch angle of fine root born on taproot
2.4 生物炭影响根系特征的广义加性模型和冗余分析

图7 生物炭施用方式与根系特征冗余分析Fig.7 Redundancy analysis of application mode of biochar and root characteristics
广义加性模型(GMA)表明,生物炭施用方式对棉花根系节间距有极显著影响(p<0.001),而根系垂直夹角对根系节间距没有产生显著影响(p>0.061)。二者交互作用显著影响棉花根系节间距(p<0.021)。土层深度(p<0.001)和生物炭交互作用(p<0.02)对棉花根长密度产生显著影响,并且土层深度影响效应高于生物炭。浅层根长密度和浅层细根分枝角呈极显著负相关。深层根长密度与浅层细根分枝角及浅层中等根分别呈极显著正相关和显著负相关。细根根长比例和极细根根长比例、中等根根长比例分别呈显著和极显著负相关,与根系平均直径呈显著正相关。浅层极细根分枝角与深层中等根分枝角呈极显著正相关(图7a)。这主要是由于生物炭导致不同土层深度各水平根系物质分配有所变化,另外,激素类物质的合成及运输也可能对上述结果产生一定程度影响。
冗余分析(RDA)表明,根系可塑性、形态特征、根长比例3类影响因素总叠加效应解释棉花浅层根长密度和深层根长密度(极细根根长比例、细根根长比例、中等根根长比例)、浅层(极细根、细根和中等根)分枝角、深层(极细根、细根和中等根)垂直分枝夹角、平均根系直径、单位体积根系质量变异的89.1%(图7b,图中Rs表示深层根长密度,Rt表示浅层根长密度,Vt表示浅层极细根分枝角,Ft表示细根分枝角,Mt表示中等根分枝角,Ms表示深层中等根分枝角),其中浅层根长密度和深层根长密度均受到浅层细根和中等根分枝角以及深层细根分枝角极显著影响(p<0.01)。上述规律可能是由于土壤资源有效性或者不同土层氮素配置诱导而形成。
3 讨论
生物炭可促进根系生长并优化作物地上地下资源分配[38-40]。本文研究表明,1%~2%生物炭与氮素复配处理下棉花根系平均直径和细根比例明显增加。这与RAZAP等[29]研究结果相似,即均促进细根生长。这是因为生物炭活化了干旱阶段土壤固定养分[12],减少养分淋失,从而可能定向诱导根系生长和发育。GRABER等[28]则认为生物炭包含的低剂量激素类物质促进根系生长,SPOKAS等[13]通过进一步研究指出生长素可能是影响其修复效果和作物增产的关键;而KOLTON等[40]则认为植物表现性能与生物炭影响的根际微生物多样性和代谢潜力强烈相关。本文试验表明,低氮0.21 t/hm2配施1%~2%生物炭处理根生物量密度显著高于常规施氮0.3 t/hm2配施相同生物炭或4%生物炭,可能是因为1%~2%生物炭弥补氮素不足并加快不同根级间氮素转运,而高剂量生物炭则可能增强了异三聚体G蛋白[30]和谷氨酸转运蛋白[31]反馈调节。由此表明,根系生长发育可能受到生物炭作用下多种因素耦合效应的共同影响。
根系构型及可塑性是植物适应环境变化、觅食的综合表征,与根系和资源的有效接触面积密切相关,其中比根长、根长密度、根系分枝角等特征决定养分摄取速率和转运效率[41]。比根长(单位质量的根长)和根长密度(单位体积根长)相比生物量更能反映植物根系投入产出比。研究表明,氮有效性较高土层,较多细根分布于该层并且根长密度较高,而氮有效性较低时则反之[41]。本文研究表明:0~10 cm土层比根长和根长密度显著高于大于10 cm土层,这与赵占军[42]在黄花苜蓿研究中的报道一致。与此同时,试验表明1%~2%生物炭显著增加大于10 cm土层根系比根长和根长密度,这是由于生物炭的活化以及吸附作用影响养分有效性,并且可能刺激主侧根氮素信号传递途径从而影响了根系构型中比根长和根长密度[31],其中将主根局部暴露于高浓度硝酸根会促进侧根发育和形成[43]。
根系分枝角也是根系构型的主要构成因子,垂直根系分枝角通常被称为向地性[34],能够反映植物根系生理变化,并能够对环境及资源变化作出反馈。垂直根系分枝角主要由激素梯度分布所决定,而硝酸根浓度会影响激素转运效率,低浓度硝酸根抑制激素转运及信号传递从而导致向地性显著减小[15,30]。近年来,模拟植物拟南芥AtDRO1过表达被证实增加垂直根系分枝角[35]。本文研究表明,0~10 cm土层根系垂直分枝角未发生明显变化,而大于10 cm土层根系分枝角发生显著变化,这与MITCHELL等[4]研究结果一致。另外,1%~2%生物炭可能减缓硝酸根淋失并抑制土壤中细菌反硝化作用,生物炭包含的少量激素或刺激植物根系激素合成及转运效率进而影响根系构型也可能是重要的影响因素。
4 结论
(1)生物炭与氮素协同作用影响棉花盛花期浅层(0~10 cm)和深层(大于10 cm)根系形态特征,具有显著差异,生物炭与氮素协同作用下0~10 cm土层根系特征未发生显著变化,而低剂量N和2%生物炭使大于10 cm土层的比根长、根长密度显著增加。1%~2%生物炭主要改变了深层(大于10 cm)棉花根系构型及形态,大于10 cm土层根系分枝角显著增大。氮素不足的条件下,2%生物炭增强根系资源获取能力。
(2)广义加性模型(GMA)显示,生物炭施用方式极显著影响根系节间距,分枝角未对根系节间距产生显著影响,与来水方向270°根系节间距变化最明显,γ3最大,为4 cm;生物炭和土层深度对棉花根长密度均有显著影响,交互作用小于土层深度单独对根长密度的影响。
(3)冗余分析(RDA)表明,根长比例、浅层和底层根系分枝角、平均根系直径以及根生物量密度可解释根长密度变异的89.1%。
猜你喜欢
果树实用技术与信息(2022年6期)2022-11-21
草业学报(2022年9期)2022-09-16
热带亚热带植物学报(2022年4期)2022-08-18
东北林业大学学报(2022年6期)2022-07-25
小小说月刊·下半月(2021年8期)2021-09-05
语文周报·教研版(2021年28期)2021-08-19
绿色科技(2021年5期)2021-04-09
农村科学实验(2021年2期)2021-04-03
陕西农业科学(2019年4期)2019-05-13
动漫界·幼教365(大班)(2018年3期)2018-05-14
