
鹅伴性羽色性状的遗传分析
2019-03-29 02:17于金成李喆于宁刘况赵辉
中国农业科学 2019年5期
于金成,李喆,于宁,刘况,赵辉
鹅伴性羽色性状的遗传分析
于金成1,李喆1,于宁1,刘况2,赵辉1
(1辽宁省农业科学院,沈阳 110161;2海城正丰牧业有限公司,辽宁海城 114222)
【目的】豁眼鹅是我国著名的白鹅品种,其1日龄雏鹅往往呈现黄色或淡黄色。然而,笔者在进行豁眼鹅纯系继代选育过程中,发现自由交配群体的后代中有20%雏鹅个体绒羽呈现浅褐色,且公母比例差别明显(公母比例接近1﹕3),推测豁眼鹅群体白羽性状存在伴性遗传可能。以雏鹅期浅褐色公鹅和黄色母鹅为亲本,利用杂交试验,检验雏鹅绒羽表型的情况是否符合孟德尔遗传规律,从而揭示鹅羽色性状的遗传方式。为此开展鹅的羽色性状遗传规律及机制研究,以期对鹅新品种或品系的培育以及鹅羽色自别雌雄配套系生产提供指导。【方法】选用40只豁眼鹅公鹅和200只豁眼鹅母鹅为亲本,组建随机交配群产生豁眼鹅纯系G1代,观察G1代中1日龄雏鹅中黄色绒羽和浅褐色绒羽表型的表现和分离比例;4只浅褐色豁眼鹅公鹅和20只黄色豁眼鹅母鹅为亲本,杂交产生F1代,观察其1日龄雏鹅中黄色绒羽和浅褐色绒羽表型的表现和分离比例。【结果】①在豁眼鹅随机交配G1代中,浅褐色绒羽表型个体的比例为19.5%。其中,公母雏之间浅褐色个体占比差别比较大,公雏中浅褐色的比例在10%左右,而母雏有30%左右的个体呈现浅褐色。根据Hardy-Weinberg定律,绒羽颜色性状等位基因频率计算如下,母雏中浅褐色个体的比例就是豁眼鹅群体中绒羽浅褐色等位基因的频率,本试验中29.8%的母雏个体是呈现浅褐色,所以该等位基因的频率约为0.3。同时,公雏中浅褐色个体比例的开方也能计算出豁眼鹅群体中浅褐色等位基因的频率,本例中,10%的公雏是浅褐色,所以该等位基因的频率约为0.3左右,同母雏计算所得基因频率比较接近。相应的,绒羽黄色等位基因频率则为0.7左右。②杂交F1中,黄羽81只,占50%;浅褐色79只,占50%。且黄羽全为公雏,浅褐色羽全是母雏。结果表明,浅褐色性状呈伴性遗传;已研究证实,银色羽(S*S)和不完全白化(S*AL)是位于Z染色体上的一对等位基因控制,不完全白化基因AL对银色羽基因S为隐性。因此,根据杂交试验结果,可以初步判断浅褐色和黄色是由一个基因座的复等位基因所决定的结果,且浅褐色对于黄色性状呈隐性遗传。【结论】本试验通过羽色性状选配杂交和随机交配试验,分析了豁眼鹅1日龄雏鹅绒羽颜色表型的分离情况,结果表明:(1)浅褐色绒羽性状相对黄色绒羽性状为隐性遗传;(2)豁眼鹅雏鹅绒羽浅褐色/黄色性状呈伴性遗传;(3)豁眼鹅雏鹅绒羽颜色性状主要为Z染色体银色羽基因座控制,控制该基因座的等位基因存在变异位点与相应表型关联。
羽色性状,隐性遗传,伴性遗传,豁眼鹅
0 引言
【研究意义】羽色是家禽一种重要的遗传标记,利用伴性羽色基因进行雏禽的雌雄鉴别,在现代家禽生产中发挥了重要的作用,产生了巨大的经济效益。豁眼鹅是我国著名的白羽鹅品种,其1日龄雏鹅往往呈现黄色或淡黄色。然而,笔者在进行豁眼鹅纯系继代选育过程中,发现自由交配群体的后代中有20%雏鹅绒羽呈现浅褐色,且公母比例差别明显(公母比例接近1﹕3),推测豁眼鹅群体白羽性状存在伴性遗传可能。研究鹅的羽色性状遗传规律及机制对鹅新品种或品系的培育以及鹅羽色自别雌雄配套系生产具有重要意义。【前人研究进展】禽类羽色性状的研究已有大量成果,主要集中在鸡[1-4]、鸭[5-9]和鹌鹑[10-12]等家禽当中,其中呈伴性遗传方式的羽色性状主要有横斑[1]和银色羽[13]。鹅的羽色研究较少,主要集中在由灰雁驯化而来的欧美鹅品种上,包括斑点、羽色稀释和浅黄色等羽色特征[14],而关于中国鹅羽色研究还未有报道。【本研究切入点】在豁眼鹅群体中发现的浅褐色雏鹅个体,随着日龄的增加,片羽的生发,其个体和黄色雏鹅一样,最终白羽覆身。这种现象,类似于鸡的银色羽(Silver, S)基因座突变形成的连锁不完全白化现象[15],即S座位只是抑制机体的红色色素[13],而浅褐色可能是该座位发生突变,使得鹅在绒羽期不能实现完全白化。早在1963年,Cole等[16]通过试验将银色羽基因座不同等位基因之间的显隐性关系确定为银羽(S*S)>金羽(S*N)>性连锁不完全白化(S*AL)。据此,我们假设决定鹅浅褐色绒羽性状的基因(S基因座)为性连锁隐性遗传,根据孟德尔遗传定律,基因型为Z Z与基因型为ZW交配产生的F1代中,母鹅为浅褐色绒羽,公鹅为黄色绒羽。以雏鹅期浅褐色公鹅和黄色母鹅为亲本,采用杂交试验,检验雏鹅绒羽颜色的情况是否符合孟德尔遗传规律,从而揭示鹅羽色性状的遗传方式。【拟解决的关键问题】通过对豁眼鹅群体中不同羽色表型的观测,找出其变化规律,阐明该性状的遗传机制,以期为鹅伴性羽色利用提供理论支撑。
1 材料与方法
1.1 试验鹅群的建立
本试验选用4只浅褐色豁眼鹅公鹅和20只黄色豁眼鹅母鹅为亲本,杂交产生F1代,观察1日龄雏鹅中黄色绒羽和浅褐色绒羽表型的表现和分离比例。此外,选用40只豁眼鹅♂和200只豁眼鹅♀为亲本,组建随机交配群,观察绒羽颜色性状在自由交配群中的传递情况。杂交群和随机交配群的公母比为1﹕5,每组5只母鹅单栏饲养。
1.2 杂交试验
选择9月龄的同日龄白羽豁眼鹅公鹅(雏期为浅褐色绒毛)4只,白羽豁眼鹅母鹅(雏期为黄色绒毛)20只,于个体栏内公母1﹕5混群饲养交配。1周后,开始收集种蛋,每10d为1个孵化批次,连续孵化3批。此外,选择10月龄白羽豁眼公鹅40只,白羽豁眼母鹅200只,于个体栏内,公母1﹕5混群饲养交配,孵化1个批次。种蛋采用电孵化器孵化,固定孵化程序,9 d左右进行一次照蛋,剔除无精蛋和死精蛋。通过观察雏鹅的羽色及性别分离情况,以推断亲本及其后代的基因型。
1.3 性状测定方法
鹅在同一栋舍内分栏饲养,自由采食、饮水。按常规方法进行饲养管理及免疫。出雏时即测定试验鹅的羽色性状情况。
记录F1代群体中不同性状个体数量,Excel软件进行统计分析。性状的分析采用卡方适合性检验,用来检验性状的观察值次数与该性状的理论比率是否相符合。
当df=1,采用英国叶斯(F. Yates)提出的矫正公式。矫正后的χ2公式如下:
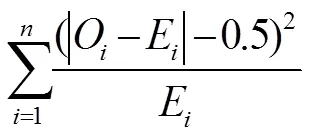
其中,i:实际观察值;i:理论值;:表型数。
卡方适合性检验的无效假设:H0:实际次数和理 论次数的偏差等于零,H1:实际次数和理论次数的偏差不等于零。如果>0.05,差异不显著,即理论值与实际值相符合;若0.01<<0.05,差异显著,<0.01差异极显著,说明理论值与实际值不符合。
2 结果
2.1 豁眼鹅随机交配群中羽色性状的表现
表1显示了纯种豁眼鹅随机交配下一代群体的羽色表型比例情况。在豁眼鹅随机交配G1代中,绒羽浅褐色个体的比例为19.5%,基本占全群的五分之一。其中,公雏中浅褐色的比例在10%左右,而母雏有30%左右的个体呈现浅褐色。
在随机交配的群体中,绒羽浅褐色鹅比例雌雄之间存在显著差异,从而推测绒羽浅褐色可能是伴性遗传。根据Hardy-Weinberg定律,绒羽浅褐色/黄色伴性性状等位基因(S基因座)频率计算如下,母雏中浅褐色个体的比例就是豁眼鹅群体中S座位AL等位基因的频率,本试验中29.8%的母雏个体是呈现浅褐色,所以AL等位基因的频率约为0.3。同时,公雏中浅褐色个体比例的开方也能计算出豁眼鹅群体中S座位AL等位基因的频率,本例中,10%的公雏是浅褐色,所以AL等位基因的频率约为0.3左右,同母雏计算所得基因频率比较接近。与AL等位基因相对应的S基因频率则为0.7左右。
2.2 浅褐色豁眼鹅公鹅与黄色豁眼鹅母鹅杂交后代中羽色性状的表现
表2显示了杂交F1群体中的羽色表型比例情况。3批总计,黄羽81只,占50%;浅褐色79只,占50%。且黄羽全为公雏,浅褐色羽全是母雏。结果表明,浅褐色性状呈伴性遗传。
由表2看出,F1代公雏中浅褐色消失了。已研究证实,银色羽(S×S)和不完全白化(S×AL)是位于Z染色体上的一对等位基因控制,不完全白化基因AL对银色羽基因S为隐性[14]。因此,根据杂交试验结果,可以初步判断浅褐色和黄色是由一个基因座的复等位基因所决定的结果,且浅褐色对于黄色性状呈隐性遗传。

表1 豁眼鹅本品种内随机交配G1代群体中羽色性状情况

表2 杂交选配F1群体中的羽色表型比例比较
综上,杂交试验中实际公母雏中羽色的比例与预测的相一致,证实了我们的决定浅褐色性状的基因为性连锁隐性遗传的假设。
3 讨论
3.1 鹅浅褐色羽色性状呈伴性隐性遗传
首次发现中国白鹅羽色伴性遗传现象。通过不同羽色个体的杂交选配,对其表型分离情况进行了详细的统计分析,结合豁眼鹅本品种内随机交配群体中羽色表型数据,证实了笔者关于决定浅褐色性状基因为伴性隐性遗传的假设。
横斑芦花性状可能是第一个被发现和研究的家禽伴性遗传现象[1,17],之后在家鸡中先后发现了金银羽[13]、羽速[18]、胫色[19]、矮小型[20]等伴性性状,在鹌鹑中发现羽色的伴性遗传[21],如今,这些性状已广泛应用于现代蛋鸡、肉鸡和鹌鹑生产中,产生了巨大的经济效益。目前,有关鹅伴性遗传性状主要有豁眼[22]和羽色,其中,鹅的羽色性状中,由灰雁驯化而来的一些欧洲鹅的羽色和斑纹性状呈伴性遗传[23-24],但尚未在由鸿雁驯化而来的中国地方鹅中发现该现象。我国早已是世界第一鹅业生产大国,肉鹅出栏量约为5亿只,约占世界出栏量的90%,鹅肉产量约120万吨,产值约300 亿元[25],但在规模化和产业化的关键技术环节,我们还缺乏相配套的研究。在我国白鹅品种发现羽色的伴性遗传现象,在鹅的杂交利用中可用于1日龄雏鹅自别雌雄,其应用前景非常广阔。
3.2 鹅羽色性状遗传规律分析
豁眼鹅雏鹅绒毛颜色有浅褐色和黄色两种。借鉴鸡和鹌鹑[15-16]银色羽基因座研究成果,笔者做了假设,即黄色由显性基因S控制,浅褐色由隐性基因AL所致,S/AL基因是性染色体上一对等位基因,其行为具有伴性遗传的特点,即ZALZAL、ZALW为浅褐色绒毛,ZSZS、ZSW为黄色绒毛。根据孟德尔遗传定律,雏期呈现浅褐色公鹅与黄色母鹅杂交下一代,母鹅为浅褐色,公鹅为黄色。从实际的试验鹅的羽色表现,正好符合孟德尔遗传规律。
禽类的羽色种类很多,但主要分为白色和有色两种。随着分子生物学的不断发展,人们逐渐揭开了羽色性状的遗传基础,各种羽色均为一对或几对基因控制,主要涉及3个基因座[26],而与白羽有关的基因座主要有2个,一个是常染色体基因座[27-29](显性白C与隐性白c),另一个是性染色体基因座[15](银色羽S)。本研究中的试验鹅均为白羽,有文献称中国白鹅的羽色基因被常染色体隐性基因控制(基因型为c/c)[14],从本试验结果上看,此结论有待商榷,至少豁眼鹅的白羽是由于性连锁显性银色羽基因座导致的。这种有争议的结果,有待于建立不同品种鹅羽色资源群,通过分析羽色表型的分离情况,来进一步阐释鹅羽色性状的特点。
3.3 鹅羽色性状分子机制的分析
随着分子生物学的不断发展,人们逐渐揭开了禽类羽色性状的分子遗传基础,各种羽色涉及众多基因,但与银色羽基因座相关的主效基因是水溶载体45家族第2成员(Solute Carrier Family 45 Member 2, SLC45A2)。Gunnarsson等[30]鉴定到与银色羽等位基因S和性连锁不完全白化等位基因AL 所对应的变异。其中,突变Tyr277Cys和Leu347Met与S对应,这两个突变抑制嗜黑素的表达,Leu347Met 突变位于基因跨膜域的高度保守区,该突变和很多银色羽品种完全连锁,如洛岛白、Fayoumi鸡和白金凤花鸡等。这些结果为研究中国白鹅羽色性状遗传机制,实现分子层面快速验证与检测提供了理论依据。
研究者[30]还将SLC45A2基因外显子1上的移码突变-106delT对应了鸡AL等位基因,该移码突变导致基因翻译的提前终止,而造成转录本无义介导的mRNA降解;将SLC45A2基因一个剪接位点突变对应鹌鹑AL等位基因,该突变导致外显子4框内跳(In-frame skipping of exon 4),造成成熟蛋白中缺失47个氨基酸。最终导致携带AL等位基因个体的真黑素和嗜黑素的表达几乎完全被抑制。该结果为我们克隆鹅SLC45A2基因、筛查变异位点、建立浅褐色性状分子快速检测方法打下了坚实的基础。
4 结论
本试验通过羽色性状选配杂交和随机交配试验,分析了豁眼鹅雏鹅羽色表型的分离情况,结果表明:(1)浅褐色绒羽性状相对黄色绒羽性状为隐性遗传;(2)豁眼鹅雏鹅绒羽颜色性状呈伴性遗传;(3)豁眼鹅绒羽颜色性状可能主要为Z染色体银色羽基因座控制,控制该基因座的等位基因存在变异位点与相应表型关联。
[1] PEARL R, SURFACE F M. On the inheritance of the barred color pattern in poultry.1910, 30(1): 45-61.
[2] KNOX C W. The genetics of plumage colour in poultry. Iowa Agricultral Experiment Station, 1926, Bulletin, No.105.
[3] NORDSKOG A W. Inheritance of shank and plumage color in the Fayoumi breed.1970, 49(6): 1735-1737.
[4] 王存芳, 李宁, 吴常信. 基于F2群体的藏鸡羽色、胫色性状的遗传分析. 遗传, 2006, 28(7): 810-814.
WANG C F, LI N, WU C X. Genetic analysis of feather color and shank color trait based on F2resource population in Tibetan chicken., 2006, 28(7): 810-814. (in Chinese)
[5] JAAP R G. Alleles of the mallard plumage pattern in ducks.1934, 19(4): 310-322.
[6] GONG Y, YANG Q, LI S, FENG Y, GAO C, TU G, PENG X. Grey plumage coloration in the duck is genetically determined by the alleles on two different, interacting loci., 2010, 41: 105-108.
[7] LI S J, WANG C, YU W H, ZHAO S H, GONG Y Z. Identification of genes related to white and black plumage formation by RNA-Seq from white and black feather bulbs in ducks., 2012, 7(5): e36592.
[8] LIN R L, CHEN H P, ROUVIER R, POIVEY J P. Selection and crossbreeding in relation to plumage color inheritance in three chinese egg type duck breeds (Anas Platyrhynchos)., 2014, 27(8): 1069-1074.
[9] ZHOU Z K, LI M, CHENG H, FAN W L, YUAN Z R, GAO Q, XU Y X, GUO Z B, ZHANG Y S, HU J, LIU H H, LIU D P, CHEN W H, ZHENG Z Q, JIANG Y, WEN Z G, LIU Y M, CHEN H, XIE M, ZHANG Q, HUANG W, WANG W, HOU S S, JIANG Y. An intercross population study reveals genes associated with body size and plumage color in ducks.2018, 9(1): 2648.
[10] LAUBER J K. Sex-linked albinism in the Japanese quail., 1964, 146(3646): 948-950.
[11] TRUAX R E, JOHNSON W A. Genetics of plumage color mutants in Japanese quail.1979, 58(1): 1-9.
[12] MINVIELLE F, ITO S, INOUE-MURAYAMA M, MIZUTANI M, WAKASUGI N. Brief communication. Genetic analyses of plumage color mutations on the Z chromosome of Japanese quail.2000, 91(6): 499-501.
[13] STURTEVANT A H. An experiment dealing with sex-linkage in fowls.1912, 12: 499-518.
[14] 陈国宏, 王继文, 何大乾, 王志跃, 王宝维. 中国养鹅学. 北京: 中国农业出版社, 2013: 43-44.
CHEN G H, WANG J W, HE D Q, WANG Z Y, WANG B W.Beijing: China Agricultural Press, 2013: 43-44. (in Chinese)
[15] WERRET W F, CANDY A J, KING J O L, SHEPPARD P M. Semi-albino: a third sex-linked allelomorph of silver and gold in the fowl., 1959, 184: 480.
[16] COLE R K, JEFFERS T K. Allelism of silver, gold, and imperfect albinism in the fowl., 1963, 200: 1238-1239.
[17] MORGAN T H, GOODALE H D. Sex-linked inheritance in poultry., 1912, 22(1): 113-133.
[18] SERBROVSKY A S. Crossing-over involving three sex-linked genes in Chickens., 1922, 56: 571-572.
[19] WARREN D C. Sex-linked characters of poultry.1928, 13(5): 421-433.
[20] MAW A J G. The inheritance of skeletal dimensions in the domestic fowl. Madison: University of Wisconsin, 1935.
[21] 庞有志, 赵淑娟. 鹌鹑羽色遗传的研究及应用. 遗传, 2003, 25(4): 450-454.
PANG Y Z, ZHAO S J. The study and use on genetics of plumage color in quail.2003, 25(4): 450-454. (in Chinese)
[22] 于金成, 李喆, 于宁, 赵辉. 基于F2群体的豁眼鹅豁眼性状遗传分析. 中国农业科学, 2016, 49(19): 3845-3851.
YU J C, LI Z, YU N, ZHAO H. Genetic analysis of hoyan trait based on F2 resource population in Huoyan Goose.2016, 49(19): 3845-3851. (in Chinese)
[23] JEROME F N. Color inheritance in geese and its application to goose breeding., 1953, 32(1): 159-165.
[24] JEROME F N. Color inheritance of geese: Study 2., 1954, 33(3): 525-528.
[25] 侯水生. 我国肉鹅产业发展应重视的问题与建议. 水禽世界, 2014(03): 6-9.
HOU S S. Problems and suggestions for the development of meat goose industry in China.2014(03): 6-9. (in Chinese)
[26] 何艺平, 许继国, 谢袖娟, 詹惠娜. 鸡羽色性状基因座研究进展. 广东农业科学, 2014, 41(9): 129-134+237.
HE Y P, XU J G, XIE X J, ZHAN H N. Research progress of feather color loci in Gallus gallus domestica., 2014, 41(9): 129-134+237. (in Chinese)
[27] Brumbaugh J A, Bargar T W, William O. A “new” allele at the C pigment locus in the fowl.1983, 74(5): 331-336.
[28] KERJE S, SHARMA P, GUNNARSSON U, KIM H , BAGCHI S, FEDRIKSSON R, SCHU K, JENSEN P, VON HEIJNE G, OKIMOTO R, ANDERSSON L. The dominant white, dun and smoky color variants in chicken are associated with insertion/ deletion polymorphisms in the PMEL17 gene.2004, 168(3): 1507-1518.
[29] CHANG C M, COVILLE J L, COQERELLE G, GOURICHON D, OULUMOUDEN A, TIXIER-BOICHARD M. Complete association between a retroviral insertion in the tyrosinase gene and the recessive white mutation in chickens., 2006, 7: 19.
[30] GUNNARSSON U, HELLSTRON A R, TIXIER-BOICHARD M, MINVIELLE F, BED'HOM B, ITO S, JENSE P, RATTINK A, VEREIJKEN A, ANDERSSON L. Mutations in SLC45A2 cause plumage color variation in chicken and Japanese quail., 2007, 175(2): 867-877.
Genetic Analysis of Sex-linked Plumage Color Traits of Goose
YU JinCheng1, LI Zhe1, YU Ning1, LIU Kuang2, ZHAO Hui1
(1Academy of Agricultural Sciences, Shenyang 110161;2Haicheng Zhengfeng Animal Husbandry Company, Haicheng 114222, Liaoning)
【Objective】The Huoyan breed of geese is a famous white goose breed in China, 1-day-old gosling of the Huoyan goose are yellow or light yellow. In the offspring of the free mating group, 20% of 1-day-old goslings were found to have a light brown variation during Huoyan goose breeding process, and the ratio of male to female is obviously different (the ratio of male to female is close to 1﹕3), it is speculated that there is a sex-linked inheritance tendency in goose plumage color trait. Using the light brown male geese and yellow female geese as the parents, the hybrid experiment was used to test whether the phenotype of the gosling downy feathers conformed to the Mendelian inheritance law, so as to understand the genetic mechanism of goose plumage color trait. The study on the inheritance and mechanism of feather color traits of goose is of great significance for the breeding of new breeds or strains of goose and the goose specialized line cultivation of auto-sexing by feather color. 【Method】40 males and 200 females of Huoyan geese were chosen to generate the G1generation of Huoyan goose pure line by random mating, and plumage color appearance and segregation ratio in G1were observed. 4 light brown males and 20 white feather color females of Huoyan geese were chosen to generate the F1generation, and plumage color appearance and segregation ratio in F1were observed. 【Result】① In the random mating G1generation of the Huoyan goose, the proportion of light brown downy feather trait individuals was 19.5%. Among them, the ratio of light brown individuals between male and female chicks is relatively large, the proportion of light brown in male chicks is about 10%, and 30% of females are light brown. According to Hardy-Weinberg’s law, the allele frequency of the color traits of the downy feather is calculated as follows: the ratio of light brown individuals in the female goslings is the frequency of the light brown trait alleles in the Huoyan goose population, in this experiment, 29.8% of the female are light brown, so the frequency of the light brown trait allele is about 0.3. At the same time, the ratio of the light brown individual in the male goslings can also calculate the frequency of the light brown allele, in this case, 10% of the male are light brown, so the frequency of the light brown trait allele is about 0.3. Correspondingly, the frequency of the yellow downy feather trait allele is about 0.7. ②Among the F1populations, 81 were yellow feathers, accounting for 50%; 79 were light brown, accounting for 50%. And the yellow feathers are all male, and the light brown feathers are all female. The results showed that the light brown traits were associated with sex-linked inheritance. It has been confirmed that silver feather (S*S) and imperfect albinism (S*AL) are controlled by a pair of alleles located on the Z chromosome, and the imperfect albinism gene AL is recessive to the silver feather gene S. Therefore, based on the results of the hybridization test, it can be concluded that the light brown and yellow are the results determined by the multiple allele of one locus, and the light brown color is recessive inheritance for the yellow trait.【Conclusion】In this experiment, the separation of 1-day-old goslings feather phenotypes was analyzed by downy feather color matching hybridization and random mating test. The results showed that the light brown downy feather color trait of Huoyan goose exhibits recessive heredity to the yellow, and demonstrates sex-linked inheritance. The downy feather color of the Huoyan goose is mainly controlled by the Z chromosome silver feather locus. The variation site of the allele controlling the locus is associated with the corresponding appearance.
plumage color trait; recessive inheritance; sex-linked inheritance; Huoyan goose
10.3864/j.issn.0578-1752.2019.05.016
2018-07-25;
2019-01-16
辽宁省科学事业公益研究基金(20170037)、沈阳市高层次人才创新计划项目(RC180047)、国家水禽产业技术体系专项资金(CARS-43-32)
于金成, Tel:024-31029891;13898156386;E-mail:yujincheng_pi@126.com。通信作者赵辉,Tel:024-31029891;E-mail:zhaohui_sy@126.com
(责任编辑 林鉴非)
猜你喜欢
儿童时代·幸福宝宝(2022年11期)2022-12-09
中国生育健康杂志(2022年4期)2022-07-13
川北医学院学报(2022年6期)2022-06-24
智慧健康(2021年17期)2021-07-30
检验医学(2021年2期)2021-03-09
魅力中国(2017年46期)2017-12-20
阅读与作文(小学高年级版)(2016年6期)2016-11-14
人生十六七(2015年5期)2015-02-28
青少年科技博览(中学版)(2015年10期)2015-01-11
