
Estimationof genetic parameters for juvenile growth performance traits in olive flounder(Paralichthys olivaceus)
2019-03-22 02:29YangzhenLiBoZhangYingmingYangSonglinChen
Aquaculture and Fisheries 2019年2期
Yangzhen Li,Bo Zhang,Yingming Yang,Songlin Chen,∗
aYellow Sea Fisheries Research Institute,Chinese Academy of Fishery Sciences(CAFS),Laboratory for Marine Fisheries Science and Food Production Processes,Qingdao National Laboratory for Marine Science and Technology,Qingdao,266071,China
bCollege of Fisheries and Life Science,Shanghai Ocean University,Shanghai,201306,China
cChina Key Laboratory for Sustainable Development of Marine Fisheries,Ministry of Agriculture,Qingdao,266071,China
dTianjin Bohai Sea Fisheries Research Institute,Bohai Sea Fisheries Research Center of Chinese Academy of Fishery Sciences,Tianjin,300457,China
Keywords:Genetic parameter Growth trait Heritability Genetic correlation Olive flounder Paralichthys olivaceus
ABSTRACT The aim of this study was to evaluate genetic parameters of growth traits for improving selective breeding in olive flounder(Paralichthys olivaceus).A total of 82 full-sib families from 2 year-classes with complete pedigree information were used to estimate heritabilities and genetic/phenotypic correlations for 4 growth performance traits(body weight BW,total length TL,body height BH and condition factor K)based on uni-trait and multi-trait linear mixed models,respectively.Data were analyzed by the restricted maximum likelihood(REML)method.The heritabilities of BW,TL and BH were 0.81,0.70 and 0.61,which are relatively high,however,heritability for K is low(0.11).For all growth traits,the common full-sib effects(c2)were at a low to moderate level(0.14-0.20);the maternal effects(m2)of the traits(except K)were also detected(0.15-0.19).The phenotypic and genetic correlations among BW,TL and BH were highly positive(0.70-0.91),however,the correlations between K and the other 3 traits was low or negligible(-0.22-0.26).According to our results,these traits(except K)could be selected and enhanced simultaneously.In addition,direct selection based on TL or BH,which is an easily measurable trait,will be more favorable than selection on BW.Moreover,our results indicated that growth parameters estimated in the juvenile stage could be used as a predictor of growth performance at later growth stages.
1.Introduction
Olive flounder(Paralichthys olivaceus)is an economically important marine aquaculture flat fish in Northeast Asia,such as China,Japan(referred to as the Japanese flounder)and Korea(Li et al.,2018).In the last decade,with the rapid development of breeding and rearing techniques in China,indoor intensive industrial aquaculture has emerged as a practice.It is precisely because of this that the market needs are satis fi ed and more importantly,this species is widely used in fisheries resource enhancement through release.For a higher yield and welfare of the animals,breeding strategies in flounder have been carefully designed.For example,growth and disease resistance performance have been investigated based on family selection(Chen et al.,2008;Tian,Xu,Liang,&Chen,2011;Zheng et al.,2016).Traditional breeding methods(e.g.crossbreeding or hybridization)and modern genetic technologies(e.g.gynogenesis,androgenesis and polyploid)(Hou et al.,2016;Wang et al.,2011,2012;Yi et al.,2012)have been practiced for genetic improvement in flounder.Recently,extensive genetic studies(e.g.genetic linkage map and QTL mapping)have been conducted on the olive flounder(Castaño-Sánchez et al.,2010;Cui,Wang,Qiu,Liu,&Yang,2015;Shao et al.,2015;Song et al.,2012;Wang et al.,2014,2016;Zhao et al.,2018).However,systematic selective breeding programs based on quantitative genetics evaluation for this species are still limited.
Growth is one of the most important traits in fish aquaculture(Gjedrem,2005),and the response to selection in growth performance traits will directly benefi t farmers.Genetic improvement programs for growth traits in aquaculture species have been successfully implemented with considerable genetic gains(Gjedrem,2012).Most estimates of genetic parameters of fish growth traits mainly focus on body weight(BW),total length(TL)and condition factor(K),such as Chinese tongue sole,Cynoglossus semilaevis(Liu,Li,Du,Shao,&Chen,2016),grass carp,Ctenopharyngodon idella(Fu,Shen,Xu,&Li,2016),Asian seabass,Lates calcarifer(Ye et al.,2017)and turbot,Scophthalmus maximus(Guan et al.,2016;Lyu,Wang,Luan,Hu,&Kong,2017).
Estimation of genetic parameters for early growth traits is of importance because selecting future breeding candidates or families at an early growth stage would decrease fish maintenance costs and accelerate breeding progress.Heritability and genetic correlation are two important parameters,which measure the relative magnitude of additive genetic variation for a trait and covariation between traits.Besides,for estimating genetic parameters,integrated pedigree information is essential in selective breeding of aquaculture species for genetic improvement and hatchery management(Gjedrem&Baranski,2009).In summary,a permanent genetic improvement can be obtained from selective breeding which is a key benefi t.In other words,the sum of the genetic improvement made in one generation will be transferred to the following generations.
Genetic parameters for growth traits at harvest(involving 186 families produced from 2012 to 2015)have been estimated for olive flounder in our previous studies(Li et al.,2018)and breeding efficiency of this species has been promoted.In China,however,to maximize benefi ts,most of the flounder famers or aquaculture industries usually eliminate 10%-20%of smaller individuals at the age of about 150 days,only 80%-90%of the larger individuals are maintained to the commercial size.Thus the production costs could be lowered significantly by reducing the duration of the rearing cycle.Since juvenile growth traits are of great interest to famers,there is an urgent need to estimate the genetic parameters of growth traits at the juvenile stage in olive flounder.Our current work advances the current state of the art in relation to the optimization of existing flounder selective breeding programs.
To our knowledge,little information about quantitative genetic studies was available in juvenile olive flounder(120-240 days).In our present study,total 82 full-sib families from 2 year-classes with complete pedigree information were used to estimate heritabilities and correlations for four growth performance traits(i.e.body weight BW,total length TL,body height BH and condition factor K)based on unitrait and multi-trait linear mixed models,respectively.Our research provides substantial support for determining future breeding strategy for olive flounder in juvenile stage.
2.Materials and methods
2.1.Brood stocks
All the olive flounder brood stocks were raised in our breeding station in Haiyang,China.The original breeding population was from three distinct geographic locations(G0),namely South Korea,Japan and Chinese wild-caught fish,which were collected in 2005(Chen et al.,2008).From 2007,these fish were used to production and selective breeding by mass spawning or artificial insemination.In the present study,63 sires and 67dams were used,which were from 6 yearclasses(Table 1)involving 3 generations(G2-G4).Note that each generation was formed in different year-classes.All the parental fish were individually tagged with Passive Integrated Transponders(PIT)(Qingdao Star fish Instruments Co.,Ltd,China)for identification during the trial.
2.2.Production of families and juvenile fish rearing
The experimental families were established and raised in ourbreeding station in Haiyang,China.The parental fish were spawned using artificial insemination procedures to create full-sib families in 2 year-classes(2016-2017).Due to a consecutive spawning type and high fecundity,some dams or sires were mated with 1-2dams or sires.Further,some sires and dams were reused across year-classes to create genetic links.Not all the matings were successful for poor egg quality.During late April and early May in 2016 and 2017,mature eggs and sperms were stripped from breeders for artificial insemination.In each year,all the families were produced in several batches(days),so the age of each family was not exactly the same(ranging from 131 to 147 days and 122-142 days for 2016 and 2017 respectively).The fertilized eggs were incubated in hatchery cages and about 6h before hatch they were transferred to 2.5m3plastic tanks for hatching and larvae rearing.
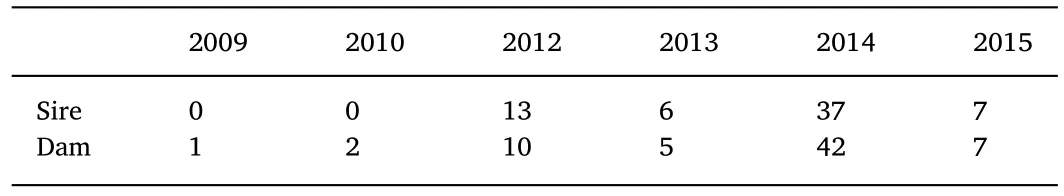
Table 1 Number of brood stocks from each year-class used in this study.
Finally,each family was cultured in a separate tank,with a similar rearing environment.Because of space limitations in the broodstock facility it was not possible to maintain duplicate or triplicate tanks of all families.For flounder larvae rearing and maintaining,the management regime was strictly maintained in different year-classes.So we assumed that all the fish were under a common environment.The tanks were supplied with filtered and sterilized sea water through a flow-through system(200%water exchange per day),with continuous aeration.From 3 days post-hatching(dph)to 18 dph,the larvae were fed with enriched rotifers,Brachionus plicatilis,and from 13 to 30 dph,enriched Artemia,Artemia salina,nauplii was supplemented.Fish were weaned onto commercial pellet diets from 25 to 30 dph and then fed with commercial feed(Santong Chubu Feed(Shandong)Co.,Ltd,China)three times daily.Before 30 dph,the water temperature was controlled at 16-20 °C,thereafter,the temperature was maintained below 25 °C.Salinity was 28-32,and dissolved oxygen was 6-8mg/L.The environmental rearing conditions of every year-class were highly consistent.At approximately 90-120 dph,800 fry of each family remained in the tanks.More details can be found in Table 2.In total,4051 fish from 82 full-sib families were used in this research.
2.3.Data collection and traits
For evaluation of juvenile growth performance,in each year-class of September or October,about 50 individuals were randomly selected from each family for measurements(BW,TL and BH)(Table 2).At the same time,the age of each family was record.Before measuring, fish were anaesthetized with MS-222(3-aminobenzoic acid ethylester methanesulfonate)to avoid handling stress.Fulton's coefficient of condition factor(K)based on BW and TL was calculated according to the following formula(Nash et al.,2006):
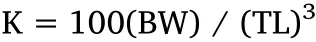
Where BW is in grams(g),and TL is in centimeters(cm).
2.4.Statistical model
The heritabilities for 4 growth performance traits were estimated using univariate models,and phenotypic and genetic correlations between different traits were estimated using a multivariate model,which were obtained based on linear mixed models in the ASReml-R 3.0 software package(Butler,Cullis,Gilmour,&Gogel,2009).For each trait,an animal model(Model 1)using the restricted maximum likelihood(REML)algorithm was implemented for the genetic analysis asfollow:
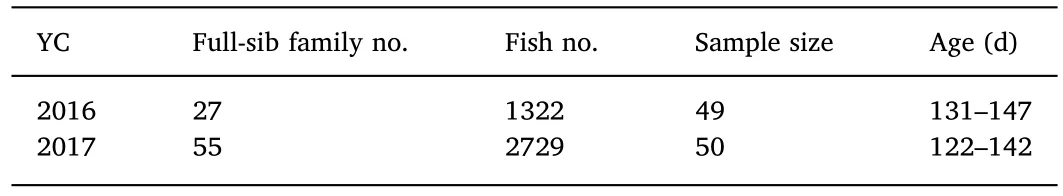
Table 2 Number of full-sib families and total fish,sample size per family and age in days post-hatching.
Model 1:y=μ+f+a+c+e
Where y is the phenotypic observation for the traits;μ is the overall mean;f is the fi xed effects,including year-class and age;a is the random additive effect for individuals;c is the effect of common full-sib families(i.e.environmental effect caused by the separate rearing of each full-sib family(tank effect)and maternal effects);e showed the residual effects.The significance of fi xed effects was estimated using Wald F statistics.
In order to detect the maternal effect,another model was used as follow:
Model 2:y=μ+f+a+m+e
Where m is the maternal effect,the other terms are the same as indicated above.In this model,the tank effect which was included in the additive effect cannot be separated.So the estimation of heritability was biased,and this model was used only for calculating the maternal effect.To verify that the Model 1 was the more appropriate model,Akaike's Information Criterion(AIC)was applied according to the ASReml-R reference manual(Butler et al.,2009).Model 1 had the smallest AIC value so it was considered to be the most suitable model(data not shown).For all analyses,the additive genetic relationship matrix included in the model was constructed based on a complete pedigree tracing back to the G0generation.The significance of the fi xed effects of year and age was detected for all traits in the current study.The fi xed effect of year was highly significant(P < 0.001)for all traits.The fi xed effect of age was significant(P < 0.05)for BW,however,for the other traits it was not in significance so it was excluded from the model.
2.5.Genetic analysis
Estimated heritability(h2),common full-sib effect(c2)and maternal effect(m2)were calculated as follows:
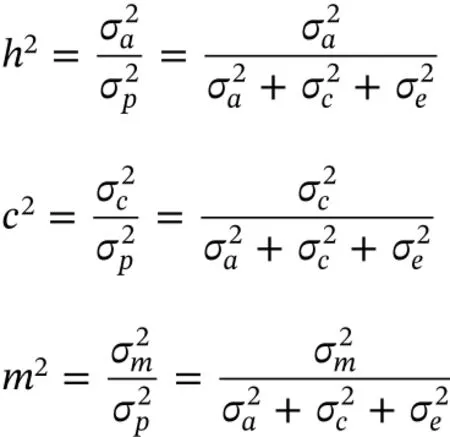
For estimating the phenotypic(rp)and genetic(rg)correlations for growth traits,Model 1 was used for multivariate analyses.The correlations were calculated as follow(Falconer&Mackay,1996):
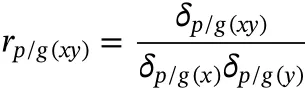
3.Results
3.1.Descriptive statistics
Overall means,standard deviations(SD)and coefficients of variation(CV)for the studied traits in each year-class are given in Table 3.The mean value of BW,TL,and BH differed between year-classes and should be attributed to the age at measurement.However,the mean value of K was at a similar level among all the different year-classes.The CVs of BW in all the year-classes were larger than the other traits.
3.2.Heritability
Estimated variance components,heritabilities,common full-sib effects and maternal effects for growth traits are shown in Table 4.Because Model 1 was the most appropriate model,the accurate heritabilities of BW,TL and BH were 0.81,0.70 and 0.61,respectively,which showed a high to very high level based on the following classifications:low(0.05-0.15),medium(0.20-0.40),high(0.45-0.60),and very high(>0.65)(Navarro et al.,2009).However,heritability for K was low(0.11),with a larger proportion of residual variance(63.6%).The heritabilities estimated based on Model 2 were all biased.Moreover,the common full-sib effects(c2)for all traits were at a low to moderate level(0.14-0.20).The maternal effects(m2)of the traits(except for K)were also detected(0.15-0.19).
3.3.Genetic correlations
The phenotypic and genetic correlations among growth traits are presented in Table 5.At both phenotypic and genetic levels,all the estimates among BW,TL and BH were highly positive(0.70-0.91),and significantly different from zero(P < 0.001).However,the phenotypic and genetic correlations between K and the other three traits were at a low positive or negative level(-0.22-0.26),and they were significantly different from zero.The significance level of the phenotypic and genetic correlations for growth traits varied depending on the traits(Table 5).
4.Discussion
4.1.Heritability
For genetic selective breeding,heritability is an important parameter for predicting how a trait will respond to selection.Evidence showed that heritabilities of growth performance traits usually range from 0 to 0.83 in aquaculture species(Gjedrem,1983;Gjedrem&Baranski,2009).A higher heritability of the desirable trait means a higher selective potential and a higher expected genetic gain.In present study,the heritabilities of BW,TL and BH at juvenile stage were at a high or very high level(0.81,0.70 and 0.61,respectively),suggesting a good selective breeding potential for these traits.Our results showed a high reliability for the estimation of genetic parameters for growth traits in juvenile olive flounder.Shikano(2007)examined heritability for body length at 210 dph was 0.81 in this species,which was highly consistent with the present study.Liu et al.(2011)found that the heritability of BW at 180 and 240 dph was 0.72 and 0.76 respectively,which was very similar to our results,but at 360 dph there was a significant reduction which was 0.47.Evidence shows that at harvest stage the heritabilities of growth traits in olive flounder were much lower.For example,at the age of 450 dph,the heritability of BW was 0.37,at the age of 720 and 830 dph it was reduced to 0.12 and 0.22 respectively(Liu et al.,2016a;2016b).Similar results can also be found in the literature about the heritability of growth traits at harvest and they ranged from 0.12 to 0.39(Li et al.,2018;Liu,Yang,Liu,&Sun,2017).From our results and previous studies in olive flounder,it was clearly evident that the heritabilities of important growth traits at the juvenile stage were much higher than at later growth stages,which could be attributed to a shorter growth time and thus fish were less influenced by environmental factors.In addition,heritability can also be affected by other factors such as the population(strain,gender,location,etc.)in which it was measured(measurement error)(Wessel,2013).
Another trait K was also investigated in this study,however,the heritability(0.11)was much lower than all the other growth related traits.This observation is consistent with our previous study thatshowed that the heritability of K was 0.17 at harvest stage of olive flounder(Li et al.,2018).Similar conclusions have been reported for other species(Domingos,Mulder,van Arendonk&Komen,2013;Dupont-Nivet et al.,2008;Fu,Shen,Xu,Liu,&Li,2015;Navarro et al.,2009;Trong,Mulder,van Arendonk,&Komen,2013).However,in another flat fish,the turbot Scophthalmus maximus(Guan et al.,2016),the heritability of K was very low(0.04),which can be explained by the morphology of the turbot,which has an oval body shape and a short length(Li et al.,2018).Even so,more research is needed to elucidate whether the existing parameters used to measure K are suitable for assessment of flat fish.If not,a modified version for the formula should be developed.
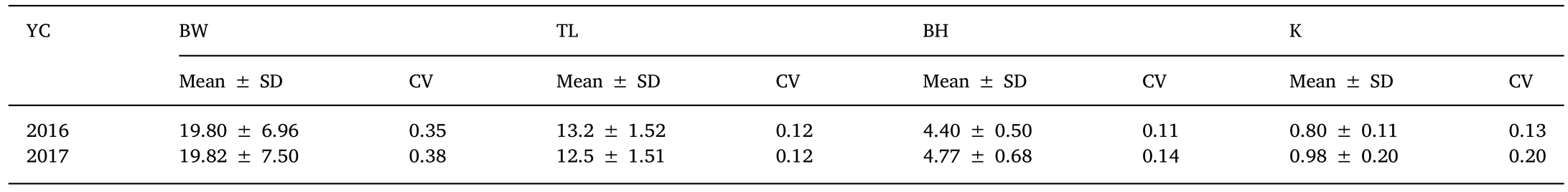
Table 3 Mean± standard deviation(SD)and coefficient of variation(CV)of BW(g),TL(cm),BH(cm),and K(g/cm3)in juvenile olive flounder in each year-class(YC).
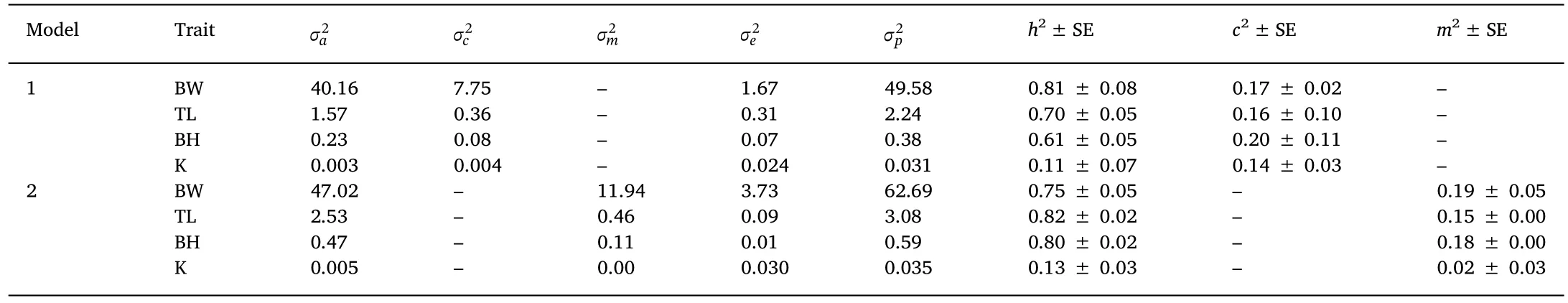
Table 4 Variance components,heritabilities,common full-sib family effects and maternal effects of growth traits of juvenile olive flounder.
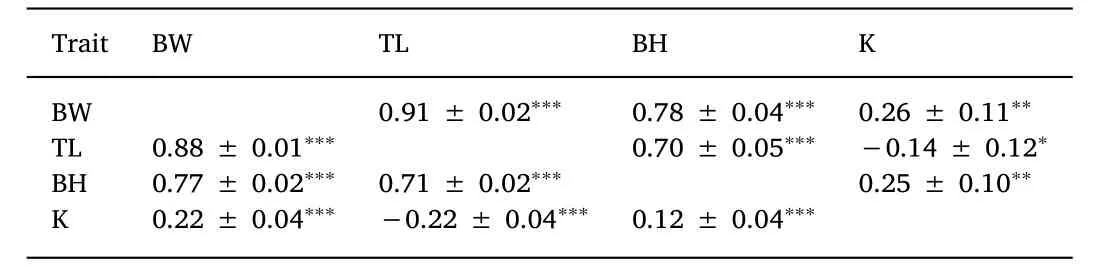
Table 5 Genetic correlations(above diagonal)and phenotypic correlations(below diagonal)among juvenile flounder growth traits.
4.2.Common full-sib family effect and maternal effect
The full-sib effect(c2)and maternal effect(m2)are important sources of genetic variation in aquaculture fish species.In the present study,the c2included the tank effect and maternal effect.However,the tank effect could not be separated(in Model 1)because each family was reared independently.The maternal effect is an important environmental and genetic source of phenotypic variation(Rasanen&Kruuk,2007);it is determined not only by environment but also by the environment and genotype of the mother.Failure to account for this significant effect may bias the estimation of genetic parameters(Fu et al.,2015).Previous studies found that the maternal effect could be detected in olive flounder,especially in the early growth stage,however,at the harvest age,it was negligible(Liu et al.,2011;Liu et al.,2016a;2016b;Shimada,Shikano,Murakami,Tsuzaki,&Seikai,2007).In our study,the m2of the growth traits(except K)was successfully separated from other effects(by Model 2),and provided evidence for the existence of maternal effect in the juvenile stage of olive flounder.However,at harvest size the effect was negligible(Li et al.,2018).
4.3.Correlations
Our research revealed high positive genetic and phenotypic correlations(>0.70)among BW,TL and BH of juvenile olive flounder,indicating that these traits share an identical genetic basis.These results were in high conformity with the results of previous studies in other growth stages of the olive flounder(Liu et al.,2011;Shikano,2008).The genetic and phenotypic correlations between growth performance traits at a given age were close to unity in olive flounder,as well as in other aquaculture species(Fu et al.,2016;Liu et al.,2016a;2016b;Lyu et al.,2017).Therefore,the selection for one trait will result in another positively correlated response.We suggest that selection for growth traits could be selected indirectly through the selection by TL or BH.Particularly since these two traits can be more easily and accurately measured with a smaller coefficient of variation and less influence from the volume of gonads,especially during the spawning season(Fu et al.,2016).The magnitude of genetic and phenotypic correlations between K and other growth traits was low(positive or negative,ranging from-0.22 to 0.26),which was close to the results previously reported in turbot(Guan et al.,2016).Further investigation is needed to determine whether K can be used to improve the efficiency of breeding programs in olive flounder,although the unique phenotype of the flat fish should be taken into account in the method of determination.
5.Conclusion
Our results confirmed that it is reliable to estimate genetic parameters for growth traits in juvenile stages of olive flounder.High heritability estimates and large magnitude of correlations between growth traits suggest that simultaneous improvement of growth traits at juvenile stage by selective breeding is feasible,eg.an increase in BW would be associated with an increase in TL or BH,and vice versa.Furthermore,our results highlight that growth parameters estimated in juvenile stages could be used as a predictor of growth performance at later growth stages.
Acknowledgements
This work was supported by Central Public-interest Scienti fi c Institution Basal Research Fund,CAFS,China(NO.2016HY-ZD0201)and Taishan Scholar Project Fund of Shandong of China.
 Aquaculture and Fisheries2019年2期
Aquaculture and Fisheries2019年2期
- Aquaculture and Fisheries的其它文章
- Full-length mRNA sequencing in Saccharina japonica and identification of carbonic anhydrase genes
- Effects of dietary lysolecithin(LPC)on growth,apparent digestibility of nutrient and lipid metabolism in juvenile turbot Scophthalmus maximus L.
- In vivo effects of neomycin sulfate on non-specific immunity,oxidative damage and replication of cyprinid herpesvirus 2 in cruciancarp(Carassius auratus gibelio)
- Effectiveness of stocking Sparus macrocephalus fry in situ in the East China Sea
- Current status of research on aquaculture genetics and genomicsinformation from ISGA 2018