
昆虫分布南界的形成原因及影响因素
2019-03-16 07:44:36党英侨王小艺杨忠岐
生态学报 2019年24期
辛 蓓,党英侨,王小艺,杨忠岐
中国林业科学研究院森林生态环境与保护研究所,国家林业和草原局森林保护学重点实验室, 北京 100091
每种生物都有适宜生存的地理范围,超出适生区域生物往往无法建立种群或完成生活史。生物的地理分布不仅取决于其扩散能力,也会受到环境、气候和生物因素等外界条件的影响[1]。生物在北半球低纬度地区形成的分布界限,常称为其分布的南界(Southern limits)[2]。南半球生物分布的南北界,恰好与北半球的称谓相反。本文讨论的分布南界,指北半球生物的分布南界。目前,关于生物地理分布的北界研究较多,仅有少量关于生物分布南界的文献报道[2](表1)。研究发现藻类、植物、动物等均存在分布南界[3- 5]。生物分布南界的形成一方面受其自身扩散能力的限制,同时也受外界条件的影响,其中气候是主要影响因素[6, 7]。在北半球低纬度地区,年均温度和年降水量都高于高纬度地区,夏季易出现极端高温,而冬季平均温度较其适生区高且低温持续时间较短,使生物在低纬度地区形成分布上的极限[5]。植物和藻类的分布南界往往是由低纬度地区的高温环境引起的[3, 4],而动物的分布除受环境影响外,还受其食物资源分布的限制[5]。
昆虫作为动物中的主要类群,其分布受环境和寄主分布的影响较大[8]。随着气候变暖,大部分昆虫的分布范围将扩大[9- 11],而一些对生境有特殊需求的物种的分布范围将受到限制[12]。目前仅有关于马铃薯甲虫Leptinotarsadecemlineata[8]、美国白蛾Hyphantriacunea[13]、秋家蝇Muscaautumnalis[14]、地中海粉螟Ephestiakuehniella[15]等昆虫分布南界的报道,且它们形成分布南界的具体原因尚不清楚。明确昆虫分布南界的形成原因,有助于准确预测全球气候变暖趋势下昆虫分布的变化,为危险性有害生物的预防与治理提供应对策略,为利用天敌昆虫进行生物防治提供指导,同时也为一些检疫害虫在其入侵地的分布范围预测和防治提供理论基础。另外,在全球气候变暖的大背景下,生物在其分布南界面临高温和昼夜温差变小等挑战,不同物种对气候变暖的适应能力不同,明确气候变化条件下物种分布南界的变化趋势对保持物种多样性具有重要意义。本文对昆虫地理分布南界的形成原因进行总结,以期对今后昆虫分布南界的研究有所启示。
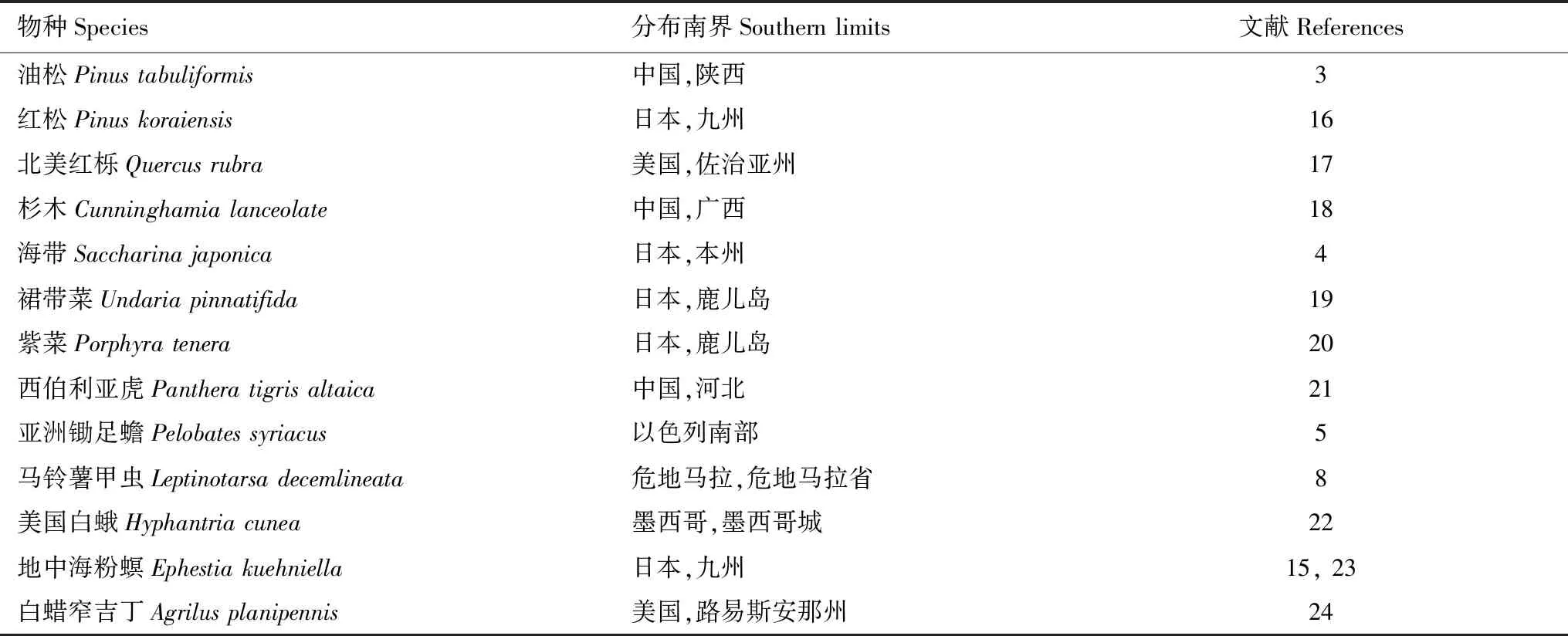
表1 部分生物的分布南界
1 温度对昆虫分布南界形成的影响
昆虫是变温动物,其保持和调节体温的能力较弱。昆虫分布南界的年平均温度较其适生区高,夏季易出现极端高温,而冬季平均温度较高且低温持续时间较短,这些气候因素均会影响昆虫的正常生长发育,最终影响其地理分布。如在适生区以南,冬尺蠖Operophterabrumata[25]、云杉芽卷蛾Choristoneurafumiferana[26]等多种森林昆虫的幼虫孵化期提前,而树木发芽的时间晚于昆虫的孵化期,导致昆虫在该区域无法建立种群[27]。玉米蚜Rhopalosiphummaidis在30℃以上不适于存活[28],麦无网长管蚜Metopolophiumdirhodum在27.5℃的恒温下未发育到成虫就全部死亡[29]。已有研究发现,极端气候条件较平均气候变化对昆虫分布产生的影响更大[30]。
1.1 高温对昆虫生理的影响
极端高温时昆虫虽然有躲避行为,但其生理结构仍会受到影响,主要包括高温致使虫体水盐平衡被打破、昆虫体壁被破坏、细胞结构发生变化等[31]。当外界环境温度升高至某一阈值时,昆虫体壁的蜡质层开始瓦解,油脂融化,导致昆虫体壁失水量加剧,昆虫水分的摄入和散失失衡,引起水分胁迫[32]和体内离子平衡被破坏[33-34]。热胁迫条件下,小金蝠蛾Thitarodesxiaojinensis因发生氧化损伤而不能存活[35]。高温导致昆虫细胞磷脂膜的流动性增强,造成细胞膜不完整和细胞骨架损坏,影响神经传导[36-37]。高温会对一些昆虫体内的重要酶类产生影响,如黑腹果蝇Drosophilamelanogaster在29℃以上饲养时,体内乙酰胆碱酯酶失去活性,不能进行神经传导,最终导致死亡[38]。
1.2 高温对昆虫生殖力的影响
在低纬度地区,夏季极端高温影响昆虫生殖器官的发育和生殖细胞的生长,最终影响其生殖效率。如29℃以上的高温使天蚕Antheraeayamamai5龄幼虫卵巢发育停止,导致其生殖力严重下降[39]。高温会影响雄虫精细胞囊、睾丸和附腺的发育[40]。棉铃虫在33℃高温条件下,精细胞囊会发生畸形[41]。遭遇高温时,黑腹果蝇的精母细胞减数分裂出现异常,雄虫失去生殖能力[42]。
通常天敌昆虫的抗逆能力较寄主害虫弱,大多数寄生蜂的最适温度范围为25—28℃[43-44],当温度超过28℃时,其雌雄性比发生变化,成虫寿命开始缩短[40]。如半闭弯尾姬蜂Diadegmasemiclausum[45]、玉米螟赤眼蜂Trichogrammaostriniae[46]的雌雄性比均会随温度的升高而逐渐降低。椰甲截脉姬小蜂Asecodeshispinarum在平均温度超过30℃时不能完成世代发育[44]。寄生蜂雌雄性比失衡,致使其交配和繁殖率严重下降,最终导致种群退化甚至消失[40]。
1.3 高温对昆虫体内共生菌的影响
很多昆虫体内存在的共生菌可以帮助其适应高温,当共生菌在极端高温下解体时,昆虫会出现死亡或生殖率下降的现象[47]。如昆虫立克次氏体Rickettsia的存在提高了B型烟粉虱Bemisiatabaci对高温的耐受力[48]。极端高温下,金毛双色弓背蚁Camponotuschromaiodes和黑色弓背蚁C.pennsylvanicus的内共生菌Blochmannia99%以上均会失去活性,导致其耐热能力显著降低[49]。当环境温度达到33℃时,家蚕追寄蝇Exoristasorbillans体内的共生菌沃尔巴克氏体Wolbachiasp.消失,沃尔巴克氏体解体后严重影响家蚕追寄蝇的生殖率[50]。Prado和Almeida[51]报道在30℃以上的温度下培养喜绿蝽Acrosternumhilare和卷心菜炑蝽Murgantiahistrionica,其体内的共生菌在两代后消失,最终导致它们无法繁殖。高温也可导致一些寄生性天敌体内的共生菌失活,降低寄生蜂的环境适应性和控害能力[40]。
1.4 低温对昆虫滞育的影响
昆虫的滞育可由光周期和温度诱导[52],低纬度地区秋季平均温度和冬季最低温度过高等影响昆虫滞育,导致其形成分布南界[53]。一般来说,昆虫的分布南界与其适生区存在明显的气候差异,冬季平均温度较其适生区高,光周期也相对较长,导致一些昆虫在分布南界不滞育或者滞育期缩短,滞育强度减弱[54]。如北美瓶草蚊Wyeornyiasmithii[55]、二斑叶螨Tetranychusurticae[56]等低纬度地区种群的滞育强度较高纬度地区弱。一部分稻绿蝽Nezaraviridula成虫在秋季平均气温偏高条件下不滞育,继续产卵,致使孵化后的子代无法度过冬季;而这部分成虫到9月中旬之后才进入滞育,导致物候失衡、繁殖困难,最终亲代和子代全部死亡[57]。
滞育的解除通常需要经历一定时间的低温,一些昆虫进入滞育后由于冬季低温持续时间过短而无法解除滞育。如棉铃虫Helicoverpaarmigera需要在3℃下保持15d才能解除滞育[58]。一般马铃薯甲虫需要在低于10℃的环境条件下持续5个月才能解除滞育、顺利完成生活史[8]。低温维持葡萄卷叶蛾Endopizaviteana蛹的滞育状态,当积温达到148日度时滞育才解除[59]。低温条件下长角直斑腿蝗Stenocatantopssplendens成虫更容易解除滞育[60]。
2 寄主对昆虫分布南界形成的影响
2.1 寄主植物对植食性昆虫分布南界的影响
植食性昆虫分布南界的形成除受温度等影响外,也受寄主植物的影响,寄主植物对植食性昆虫的生长发育和分布等起着直接的调控作用。寄主植物为植食性昆虫提供氨基酸、维生素、甾醇等营养物质以及必要的活动场所。不同地区的寄主植物种类存在差异,其体内营养成分的品质和含量也明显不同[61]。此外,为免受植食性昆虫危害,植物可产生多种不同的毒素、拒食剂等抗性物质来抑制昆虫的生长发育、免疫反应和基因表达[62]。
为完成正常的生长发育,植食性昆虫对寄主植物种类及利用部位进行选择,从而产生食性的分化[63],单食性和寡食性昆虫的分布受其寄主植物分布的影响更明显[63-64]。如寡食性昆虫白蜡窄吉丁Agrilusplanipennis只以木犀科Oleaceae白蜡属Fraxinus少数几种植物为食[65],其在我国的分布范围因受到南方抗性树种光蜡树F.griffithii、中国白蜡F.chinensis等的限制,目前仅在北方部分地区有发生(内部资料)。马铃薯Solanumtuberosum是马铃薯甲虫的最适寄主,在马铃薯甲虫分布南界形成中起着重要作用[8]。盾负泥虫Lemascutellaris只取食鸭跖草Commelinacommunis,其分布南界受寄主鸭跖草分布范围的影响较大[66]。
2.2 寄主对天敌昆虫分布南界的影响
天敌昆虫包括捕食性和寄生性两类[67],寄主昆虫主要针对天敌昆虫中的寄生性类群而言,寄生性昆虫特定阶段的正常生长发育和生活史的完成需要依靠寄主昆虫。在取食消耗寄主营养的过程中,寄生性昆虫会受到来自寄主昆虫的免疫系统等多方面的抵抗作用。为顺利完成发育和繁殖,寄生性昆虫发展出一系列针对寄主昆虫的调控机制。经过长期的相互作用,不同的寄生性昆虫的寄主专化性和寄主适应性差异更加明显,寄生性昆虫表现出更强的寄主偏好性[68],因此寄生性昆虫的种群动态和空间分布格局会受到寄主种类、大小、虫态、虫体生理状况、行为习性及适生环境等多种因素的影响[69]。如烟蚜茧蜂Aphidiusgifuensi专性寄生有翅桃蚜Myzuspersicae,后者的迁飞行为有利于该寄生蜂的传播扩散[70]。板栗潜叶蝇Camerariaohridella的分布、传播扩散方向及种群密度均对其小蜂类天敌存在影响,其中该寄主昆虫的种群密度对小蜂产生的影响最为显著[71]。恩蚜小蜂Encarsiaaleurochitonis、桨角蚜小蜂Eretmocerusrajasthanicus等主要寄生烟粉虱,其分布范围受寄主粉虱的种类和空间分布状况等因素的限制[72]。
一般来说,天敌昆虫直接或间接以植食性昆虫为食,其搜寻食料的行为较为复杂,包括远距离定向和近距离定位等,搜寻过程中除受自身的视觉、嗅觉等影响外,还会受到取食对象植食性昆虫及其寄主植物等多种因素的影响[73]。一般认为天敌昆虫对取食对象的定位主要依靠植物遭受害虫为害后所释放的挥发性物质[74]。如小黑瓢虫Delphastuscatalinae专性捕食烟粉虱,主要通过感受烟粉虱的寄主植物所释放的挥发性物质来搜寻猎物[75]。二化螟盘绒茧蜂Cotesiachilonis利用受到二化螟Chilosuppressalis危害的水稻所释放的化合物来进行定位[76]。水果受到桔小实蝇Bactroceradorsalis危害后产生的挥发物对阿里山潜蝇茧蜂Fopiusarisanu具有引诱效果[77]。因此,植食性昆虫寄主植物的种类、适生环境和健康状况等都会在一定程度上间接影响天敌昆虫的分布南界。需要注意的是,天敌昆虫包括专食性、广食性等类群,广食性天敌对取食对象的选择范围较大,受寄主或猎物的限制作用较小[78];而专食性昆虫因取食对象单一,因此受取食对象及其寄主植物的影响更大。
3 人类活动对明确昆虫分布南界的影响
昆虫本身的迁飞和扩散行为较为复杂[79],一些种类当前的自然分布或许远未到达南界,而人类活动在一定程度上可能会加速其扩散。在全球气候变暖和国际贸易快速发展的背景下,昆虫的传播扩散受人类活动的影响加剧。不同地区间货物流通加快,苗木材料和木制加工产品的运输往来逐渐增多,昆虫被携带和传播的机会相应增大。昆虫一旦扩散至条件适宜的环境中就极有可能成功定殖,从而扩大其分布范围。
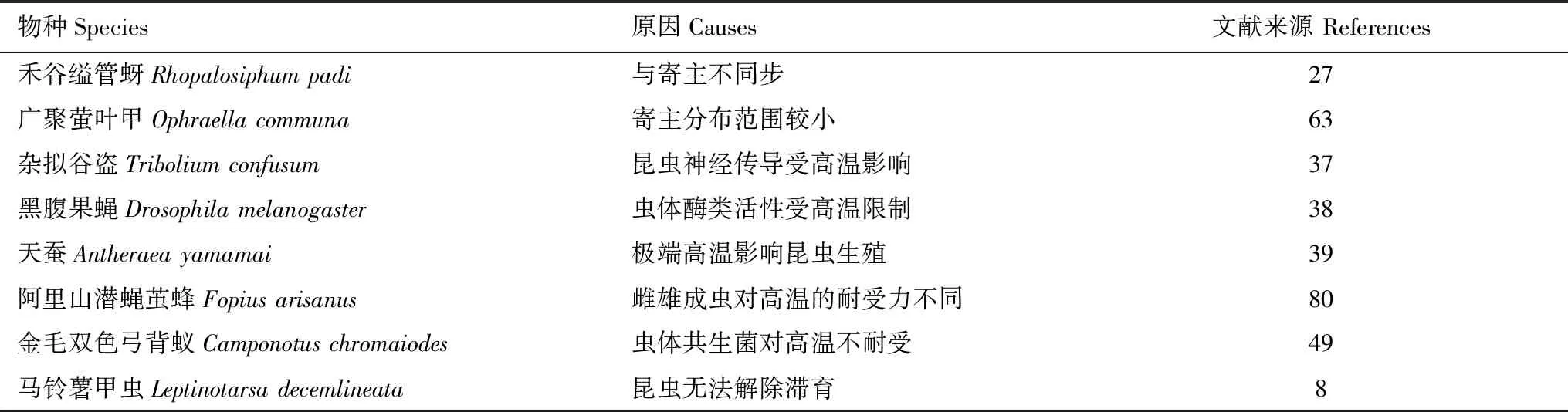
表2 一些昆虫分布南界形成的主要原因
4 结语与展望
昆虫的分布南界是指其在自然状况下的地理分布界限,但昆虫本身就具有扩散和迁移的能力,而后者会受环境、遗传、分子等多方面的调控[80],因此明确这一界限并不容易。目前大部分昆虫的分布南界尚不清楚,其形成原因除受外界环境条件影响外[14],如极端高温限制昆虫正常生殖发育[81],冬季低温较高导致昆虫无法打破滞育[8]等,寄主也是重要的影响因素。
随着全球气候逐渐变暖,大部分植物的分布范围扩大,导致一些植食性昆虫的分布南界发生相应改变[82]。对全球气候变化的预测分析认为,2100年地中海粉螟的分布将随气候变暖北移,分布面积收缩,日本西部的四国和九州不再适宜其定殖[53]。对“寄主植物—植食性昆虫—寄生性天敌”三级营养体系结构来说,气候变暖将使受低温限制的寄主植物和昆虫的分布范围向两极扩大,这些昆虫可能挤占另外一些原本处于分布南界的昆虫的生态位从而形成竞争,最终导致分布南界北移[82-83]。
在全球气候变化的背景下,昆虫的地理分布将随温度的升高发生一系列变化[82],正确估计气候变暖对昆虫分布南界的影响,分析害虫分布南界的变化趋势,可为评估害虫对低纬度地区生态环境和农林生产的危害提供理论依据。而研究昆虫分布南界形成的原因和影响因素,探明昆虫适宜生长发育的最适温区,结合寄主植物分布,也可以为合理造林提供科学指导。一些检疫害虫由于受到人为措施的影响,其分布范围较适生区小,而一旦突破检疫封锁,这些害虫将逐渐扩散至其他适生区。人类活动虽然不会改变昆虫的自然分布南界,但可以通过人工检疫等手段控制害虫的传播扩散[84],在害虫防治生产实践中发挥一定作用。
猜你喜欢
中国野生植物资源(2023年9期)2023-10-20 09:11:34
科普童话·百科探秘(2023年7期)2023-08-22 09:14:45
中国中医药信息杂志(2023年6期)2023-06-13 02:05:36
植物资源与环境学报(2020年2期)2020-04-20 07:18:02
上海农业学报(2020年1期)2020-03-25 08:44:36
竹子学报(2018年2期)2018-10-25 02:02:12
现代园艺·综合版(2017年1期)2017-07-14 16:37:59
江苏农业科学(2017年7期)2017-05-23 23:10:25
农业与技术(2016年19期)2016-12-12 02:23:50
数学大王·低年级(2016年12期)2016-09-10 07:22:44
