
川陈皮素对MC3T3-E1细胞成骨分化影响的实验研究*
2019-03-14 08:47兰庭超
中华老年口腔医学杂志 2019年1期
李 强 兰庭超 卢 芸 陈 静
传统中药应用于骨代谢疾病治疗已有上千年历 史,目前认为,传统中药具有促进骨形成及抑制骨吸收的双重作用,其活性成分通过多靶点协同调控骨稳态关键转录因子发挥作用[1,2]。川陈皮素是传统中药陈皮中的主要活性物质,具有抗氧化、抗肿瘤、抗炎及调节免疫等多种生物活性[3,4]。有研究表明,川陈皮素能通过丝裂原活化蛋白激酶(mitogenactivated protein kinase,MAPK)信号通路抑制破骨细胞成熟,促进去卵巢小鼠骨形成,降低实验性牙周炎引起的牙槽骨骨吸收[5,6]。目前,川陈皮素对成骨细胞分化的作用及机制尚不清楚。本研究将以小鼠前体成骨细胞MC3T3-E1为细胞模型,探讨川陈皮素对成骨细胞分化的作用及相关机制。
1.材料与方法
1.1 主要试剂与设备 MC3T3-E1细胞(中科院细胞所,中国),α-MEM培养基(Hyclone,美国),胎牛血清(四季青,中国),CCK-8(同仁化学,日本),碱性磷酸酶活性检测试剂盒(碧云天,中国),RT-PCR试剂盒(TaKaRa,日本),BMP-2信号阻断剂noggin(CST,美国),川陈皮素(同田生物,中国),RT-PCR仪(Bio-rad,德国)。
1.2 MC3T3-E1细胞的培养及传代 细胞复苏后,使其接种于培养瓶中,放在37℃、5%CO2条件下的恒温培养箱中孵育,16~24h后换液,每2~3d更换培养液,待细胞生长融合至80%~90%,进行传代。α-MEM细胞培养液由α-MEM培养基、10%胎牛血清配制而成。成骨培养基由α-MEM培养基中添加10%胎牛血清、10mM β-甘油磷酸钠、50μg/ml L-抗坏血酸、100U/ml青霉素、100U/ml链霉素配制而成。
1.3 CCK-8检测 川陈皮素(纯度为99.5%)用DMSO溶解后,α-MEM培养基稀释至实验所需浓度,用于后续细胞实验(DMSO终浓度<0.5%)。将MC3T3-E1细胞以4×103细胞/孔密度接种于96孔板,培养24h待细胞贴壁后,分别加入川陈皮素终浓度为0,10-9,10-8,10-7,10-6,10-5M的细胞培养液,每组6个复孔。培养48h后,每孔加入10μl CCK-8检测液,孵育1h后,抽取150μl上清液用酶标仪测OD 450 nm波长吸光度值。
1.4 碱性磷酸酶(ALP)染色及活性定量检测将MC3T3-E1细胞以1.5×105细胞/孔密度接种于24孔板,培养24h待细胞贴壁后,分别加入川陈皮素终浓度为 0μM、0.1μM、1μM、10μM 的成骨培养液,每组6个复孔。分别培养10d后进行ALP染色及活性定量检测。细胞经过PBS漂洗及多聚甲醛固定,使用BCIP/NBT工作液染色后,镜下观察并拍照。ALP活性定量检测使用对硝基苯酚(p-nitrophenol,pNP)法,PBS漂洗细胞后,加入组织裂解液裂解细胞,离心收集细胞蛋白悬浮液,使用p-NP法,酶标仪于405nm波长测定吸光度计算各组样本的ALP活性值。采用BCA蛋白浓度测定法检测样品细胞总蛋白含量:采用倍比稀释法配制标准溶液,取样品及标准品测562nm波长OD值,根据标准曲线计算各组样品细胞总蛋白量。ALP单位活性=ALP活性值/总蛋白质量。
1.5 成骨分化相关基因检测 将MC3T3-E1细胞以1×105细胞/孔密度接种于24孔板,孵育24h,加入川陈皮素终浓度为0μM、0.1μM、1μM、10μM的成骨培养液,每组3个复孔。细胞培养7d后,细胞样品加入预冷的Trizol,提取总RNA后检测其纯度,根据RNA逆转录试剂说明书逆转录合成cDNA。将合成后样品进行实时定量PCR(RTPCR)检测成骨分化相关基因BMP-2、COL-I、OCN、Runx2的表达。每组设置3个复孔。反应条件:50℃,2min;95℃,10min;95℃变性15s,60℃退火20s,72℃延伸30s,共40个循环。以管家基因β-actin作为内参,目的基因表达通过2-ΔΔCt进行相对定量。各引物序列见表1。

表1 RT-qPCR检测的基因引物序列
1.6 BMP-2信号通路抑制剂作用检测 将MC3T3-E1细胞以1×105细胞/孔密度接种于24孔板,待细胞贴壁后,分别加入Noggin(100ng/ml)及川陈皮素(10μM),使其分为三组,A:对照组;B:川陈皮素(10μM)组;C:Noggin(100ng/ml)+川陈皮素(10μM)组,每组3个复孔。细胞成骨培养7d后,按照上述实验步骤将各组样品进行RT-PCR检测 BMP-2、COL-I、OCN、Runx2基因表达。
1.7 统计学分析 所有数据均表示为平均值±标准差,使用SPSS 22.0统计分析软件,用t检验和方差分析进行统计学分析。P<0.05被认为有统计学意义。
2.结果
2.1 川陈皮素对MC3T3-E1细胞增殖的作用CCK-8法测定结果显示:与0μM浓度组(对照组)相比,川陈皮素处理48h后,10-9、10-8M浓度组MC3T3-E1细胞的增殖活性上升,但与0μM浓度组之间无显著性差异;10-7,10-6,10-5M浓度组细胞的增殖活性上升,与0μM浓度组之间有统计学差异(P<0.05)(图1)。根据不同浓度川陈皮素溶液对MC3T3-E1细胞增殖活性的影响,可确定后续实验川陈皮素的浓度分别为:0.1μM、1μM、10μM。
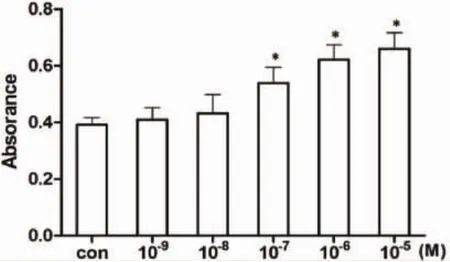
图1 川陈皮素对MC3T3-E1细胞增殖的作用
2.2 川陈皮素对MC3T3-E1细胞ALP活性的影响 MC3T3-E1细胞经不同浓度的川陈皮素处理10d后,进行ALP染色可见细胞被染成灰黑色,染色深度且呈药物浓度依赖性,川陈皮素溶液浓度越高,ALP染色结果越深(图2A)。ALP活性定量结果与川陈皮素溶液浓度呈递增效应,结果显示:川陈皮素处理10d后,0.1μM、1μM、10μM三个实验组结果与0μM对照组均有统计学差异(P<0.05)(图 2B)。

图2 川陈皮素作用下MC3T3-E1细胞的ALP染色及活性定量检测
2.3 川陈皮素对成骨标志基因表达的影响 诱导MC3T3-E1成骨分化7天后,RT-qPCR结果显示,与0μM对照组相比,0.1μM、1μM、10μM浓度的川陈皮素升高了成骨分化相关因子BMP-2、COL-I、Runx2、OCN 的基因表达(P< 0.05)(图 3)。
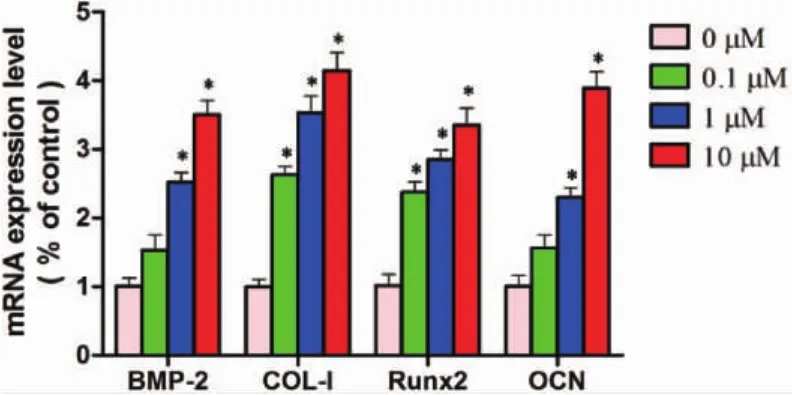
图3 川陈皮素对MC3T3-E1细胞成骨基因表达的影响
2.4 Noggin部分抑制了成骨标志基因的表达为研究川陈皮素是否通过BMP-2途径促进MC3T3-E1成骨分化,使用BMP-2信号抑制剂Noggin预处理细胞2 h,再检测川陈皮素对成骨基因表达的影响。根据ALP及成骨标志性基因检测结果,本部分实验采用了10μM川陈皮素作为药物处理对照组。结果发现,川陈皮素+Noggin组处理后,BMP-2、COL-I、Runx2、OCN基因的表达水平降低,与川陈皮素10μM处理组之间存在统计学差异(P<0.05)(图4),且成骨基因的表达水平均高于0μM对照组,并且与0μM组之间存在统计学差异(P< 0.05)(图 4)。

图4 川陈皮素及BMP-2通路抑制剂作用后成骨基因的表达
3.讨论
成骨细胞来源于骨髓间充质干细胞,在骨形成过程中,随着细胞外基质的不断分泌、矿化,最终成熟为骨细胞。因此,研究如何促进成骨细胞的分化及功能对于骨再生修复有着重要的临床意义。目前,一些蛋白类生长因子,如骨形成蛋白,确实在骨修复中被证实具有促进骨组织再生的优势。然而,由于该类生长因子的成本和存储要求均较高,使得其在骨再生修复中的广泛应用受到限制。近年来的研究发现,传统中药能够通过影响成骨细胞功能,促进成骨细胞分化,实现骨组织再生[2]。
本研究中,川陈皮素促进了MC3T3-E1细胞的增殖,升高了成骨分化标志物ALP的活性,并激活了成骨关键因子Runx2、COL-I及OCN的基因表达。Runx2是骨形成早期最重要的核心转录因子,Runx2不仅能被多种细胞因子、激素、力学刺激等因素所调控,还能够激活下游细胞外基质蛋白如OCN、COL-I的转录和表达[7-9]。我们的研究结果表明,川陈皮素促进了成骨前体细胞MC3T3-E1的成骨分化。川陈皮素属于母核结构为2-苯基色原酮的黄酮类化合物,诸多研究表明,黄酮类化合物如淫羊藿苷、槲皮素、金雀异黄酮等均能调节Runx2的表达,促进成骨细胞的增值与分化[10,11]。我们推测,由于化合物的生理活性与其化学结构特征密切相关,川陈皮素具有的黄酮母核结构以及羟基化模式可能是其促进成骨细胞增殖与分化的重要原因。
BMP家族信号分子能调控成骨细胞的分化及骨形成,作为BMP家族的重要成分之一,BMP-2在细胞及动物学实验中均证实有较强的促成骨作用[12]。BMP-2信号经配体与细胞膜表面的BMP受体结合,再激活BMP信号通路特异性调节蛋白Smad1/5/8磷酸化,Smad1/5/8与Smad4结合后,进入细胞核内,通过识别Runx2基因启动子区域,激活Runx2,从而促进基质分泌及骨形成[13]。本研究发现,川陈皮素能激活BMP-2基因表达,应用BMP-2通路阻断剂Noggin,发现Noggin能明显抑制川陈皮素激活的成骨标志性基因的表达下降。我们推测,BMP-2信号可能参与了川陈皮素调控MC3T3-E1成骨分化的过程。在成骨分化过程中,各种成骨信号通路与成骨信号因子彼此相互联系、相互影响,协同调节细胞的增殖与成骨分化。在本实验中,BMP-2、Runx2、OCN、COL-I基因水平较未加阻断剂时有显著下降,但仍高于0μM组水平,说明川陈皮素可能同时激活了其他成骨相关信号通路。
有研究表明,川陈皮素能通过MAPK信号通路,抑制转录因子NF-kB与AP-1活性,抑制破骨细胞成熟,降低骨质疏松及骨关节炎小鼠的骨吸收[5]。Tsukasa等的研究发现,川陈皮素能抑制脂多糖诱导的破骨细胞炎症反应,降低实验性牙周炎引起的牙槽骨骨吸收[6]。以往的研究表明川陈皮素能明显抑制破骨细胞功能,对炎症及雌激素缺乏引起的骨量丢失有明显抑制作用。本研究发现,川陈皮素能促进成骨细胞分化,结合以往的研究,我们推测,川陈皮素可能通过促进骨形成及抑制骨吸收的双重作用调控骨改建过程,实现骨组织再生修复,相关研究仍有待深入探讨。
猜你喜欢
中日友好医院学报(2022年1期)2022-11-28
中国药理学通报(2022年6期)2022-06-08
清华金融评论(2022年4期)2022-04-13
天津医药(2021年6期)2021-12-08
口腔医学(2021年10期)2021-12-02
中华骨与关节外科杂志(2021年12期)2021-08-31
国际放射医学核医学杂志(2021年10期)2021-02-28
山东医药(2020年36期)2020-12-31
房地产导刊(2020年7期)2020-08-24
中国外汇(2019年14期)2019-10-14
