
厦门地区低致病性禽流感病毒流行病学调查与分析
2019-03-07 08:43:06,,,,
中国人兽共患病学报 2019年2期
, ,, ,
禽流感病毒可以感染家禽、野禽甚至哺乳动物并致死,不仅给养殖业造成重大经济损失,亦危害公共卫生安全,是重要的人兽共患病病原。目前禽流感病毒有18个H亚型(H1 -H18)及11个N亚型(N1-N11),其中H1-H16亚型的流感病毒均可在水禽中分离到[1],而H17和H18自蝙蝠中检出[2]。野禽是禽流感病毒的天然宿主,并可通过迁徙,造成世界范围的大流行;猪和水禽是禽流感病毒的混合器,可以同时感染多种不同亚型的流感病毒,并在其体内发生重组,产生新的亚型;家禽活禽市场是禽流感病毒的重要储存库,大量不同种属、不同产地来源的禽类,高密度的集中在一起,加上管理和消毒不到位,造成禽流感病毒的长期存在及循环,并可借此途径感染人。
禽流感病毒分为高致病性(High Pathogenic Avian Influenza Viruses, HPAIV)和低致病性(Low Pathogenic Avian Influenza Viruses,LPAIV)两大类。高致病性禽流感因其在禽类中传播速度快、危害大、致死率高,被世界动物卫生组织列为A类动物疫病,成为疫病防控的重点。而LPAIV感染家禽后症状不明显,容易被忽视,因此得以长期、广泛的分布,周而复始的循环及变异使其不断适应当下养殖环境,进而演化成为具有耐药性及跨种属感染的新型毒株。有些低致病禽流感病毒如H7N9、H6N1、H10N8、H9N2,甚至可以突破宿主屏障感染人并致死[3-6],存在着极大的安全隐患。因此只有同时加大对LPAIV的监测力度,掌握本地区家禽禽流感病毒的真实带毒情况,才能科学有效的防控疫病。
1 材料与方法
1.1样品来源 选取2016年11月至2017年3月采集的厦门主要几家活禽交易市场的家禽喉、泄殖腔棉拭子及饲料、水等环境样品进行禽流感的检测及分型鉴定。同批次且相同来源的样品为1个群,共采集75群,534份棉拭子,包括鸡25群,265份;鸭24群,152份;环境26群,117份。
1.2样品的保存与运输 保存液PBS购自HyClone,头孢西丁钠购自致君(深圳)制药有限公司,甘油(分析纯)购自厦门绿茵试剂玻仪有限公司。PBS使用前,配置为头孢西丁钠浓度为0.2%、甘油浓度20%的工作液,分装在2 mL的EP管中,用于样品的保存。样品采集后立即置于含冰块的泡沫箱中,4 h内送实验室检测或-70 ℃保存,检测过程中避免样品的反复冻融。
1.3检测试剂及仪器 HA禽流感病毒通用诊断试剂盒购自QIAGEN,H1-H16亚型及N1-N9亚型的分型诊断试剂盒购自硕世生物科技有限公司和深圳澳东检验检测科技有限公司,核酸提取试剂(MagMAXTM-97Viral RNA)购自Thermo Fisher Scientific。荧光定量PCR仪(Cobas z 480)购自RoCho,全自动核酸提取仪购自瑞士TECAN。
1.4检测及分型鉴定 待检样品经全自动核酸提取仪提取核酸后,用HA禽流感病毒通用诊断试剂盒对其进行检测,检测结果处于临界值的可疑样品,经重复检测结果仍为可疑时判为阳性,用于禽流感病毒检出率的统计。从检出强阳性的群中,混合同群的强阳性核酸样品,用亚型分型诊断试剂盒进一步进行H1-H16及N1-N9的分型鉴定,计算不同亚型病毒的群检出率。
2 结 果
2.1禽流感病毒的检出率 2016年11月至2017年3月,在本地几家活禽交易市场共抽检75个群534份样品,检出禽流感病毒阳性的群33群,阳性样品216份,群阳性率为44.0%,样品阳性率为40.4%。其中抽检鸡群25群265份样品,检出阳性的鸡群15群,阳性样品111份,鸡群阳性率为60.0%,样品阳性率为41.9%;抽检鸭群24群152份样品,检出阳性的鸭群11群,阳性样品65份,鸭群阳性率为45.8%,样品阳性率为42.8%;抽检环境群26群117份样品,检出阳性的环境群7群,阳性样品40份,环境群阳性率为26.9%,样品阳性率为34.2%,详见表1。
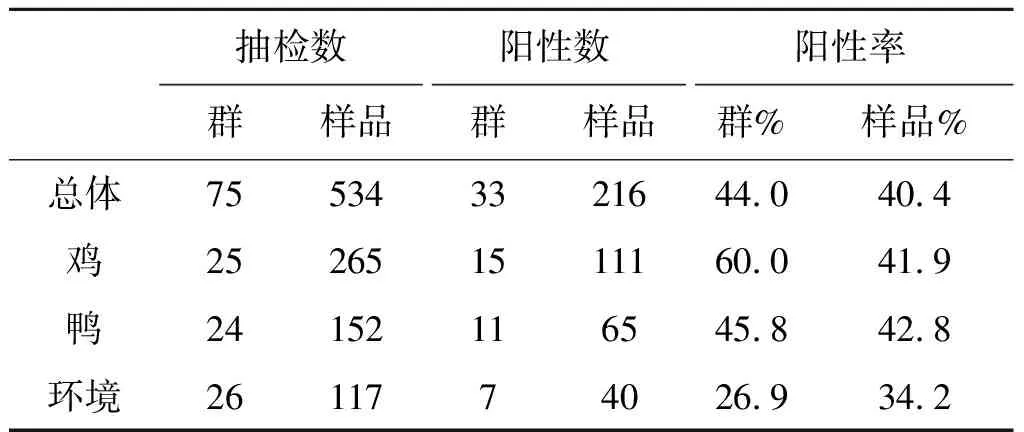
表1 活禽交易市场禽流感病毒检出率Tab.1 Detection rate of avian influenza virus in the live poultry market
2.2禽流感病毒亚型鉴定结果 从14个检出强阳性的群中,鉴定出6种HA亚型,不同亚型病毒群检出率由高到低依次为H9(100%,14/14)、H6(35.7%,5/14)、H11(28.6%,4/14)、H12(21.4%,3/14)、H1(14.3%,2/14)、H3(7.1%,1/14);检出5种NA亚型,不同亚型病毒群检出率由高到低依次为N2(92.9%,13/14)、N6(42.9%,6/14)、N3(35.7%,5/14)、N8(28.6%,4/14)、N9(21.4%,3/14);H9亚型,N2亚型检出率特别高,详见表2。
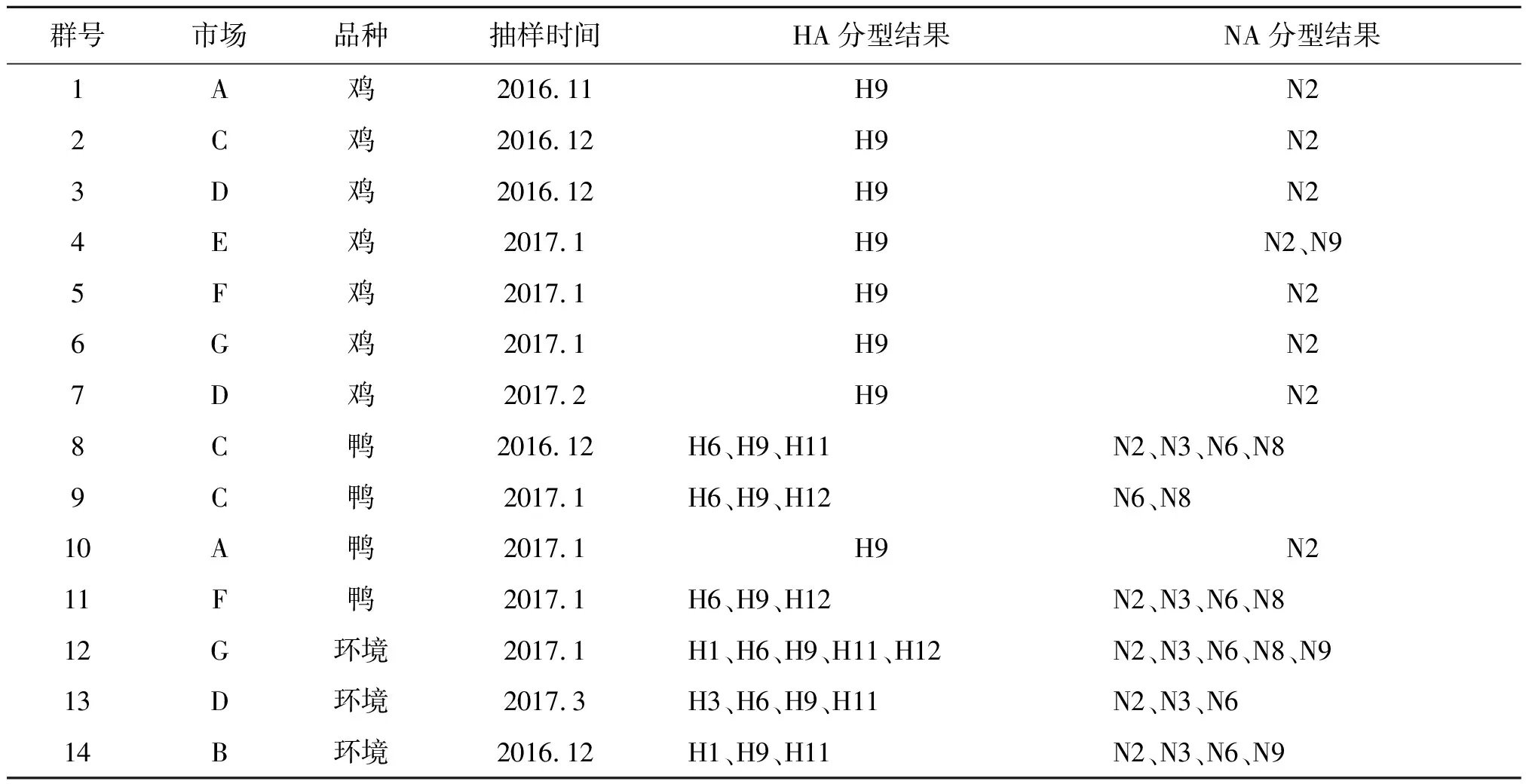
表2 活禽交易市场部分阳性样本HA、NA亚型分型结果Tab.2 Results of HA and NA subtypes typing of parts of sample in live poultry market
2.3禽流感病毒混合感染情况 交易市场中环境样品中可检出多种亚型的禽流感病毒。鸡群的混合感染不明显,7个禽流感阳性的鸡群中,有6个群体仅检出H9N2亚型病毒,混合感染率为14.3%(1/7);而鸭的混合感染情况较为严重,4个禽流感阳性的鸭群中有3个存在混合感染,混合感染率为75.0%(3/4),并且均是3种亚型以上的混合感染;3个禽流感阳性的环境群中均同时4种亚型以上的病毒。
3 讨 论
家禽的禽流感感染有很强的季节性,以冬季最高,春秋季次之,因此本研究选取2016年11月—2017年3月的时间段进行抽样检测,根据近几年来厦门市禽流感监测结果,这5个月中禽流感检出率最高。选择活禽交易市场作为研究对象,是因为活禽交易市场是禽流感病毒的重要储存库,大量不同种属、不同产地来源的禽类,高密度的集中在一起,加上管理和消毒不到位,易造成禽流感病毒的长期存在及循环,分离到的亚型会更丰富。目前尚未见到厦门市低致病性禽流感病毒亚型分型情况的报道,该研究弥补了数据的空白,为有效的防控提供科学依据。
研究亚型分型时,筛去检测结果中弱阳性和临界值的阳性样品,选择具有标准荧光曲线,Ct值小于30的阳性样本进行检测,有利于确保分型结果的准确性。结果提示该时期厦门市至少存在6种HA亚型和5种NA亚型的低致病性禽流感病毒,HA检出率由高到低依次为H9、H6、H11、H12、H1、H3;与华东地区2002-2010年检出的亚型相比存在较大的差异,华东地区有8种HA亚型,检出率由高到低依次为H3、H6、H11、H1、H4、H9、H10、H8,主要流行的亚型有H3、H6、H11、H1、H4几种,而H9、H10、H8较少检出[7-9]。两地亚型情况存在差异,一是厦门市H9亚型检出率最高,在所有检测的禽流感阳性禽群及环境样品中均检出H9亚型,而华东地区H9检出率不高;二是厦门市未检测出H4、H8和H10亚型;三是厦门市在3个禽群中检测到H12亚型,而这种亚型在华东地区尚未见报道。厦门市与华东地区H6亚型的检出率均高居第二,可见这种亚型广泛分布在华东和福建地区,仍是目前主要流行毒株。笔者认为亚型检出率差异大的原因主要有以下4点:一是研究的年份不同,不同亚型毒株有自身的流行规律,每隔几年就会出现一次大流行;二是不同的地域、气候、温湿度条件,都会影响流感病毒的进化及流行情况,有些毒株较适应当地的环境,就形成了当地的流行毒株;三是不同地区家禽的种群结构不同,陆禽和水禽的品种、比例存在差异,饲养模式不同,防疫条件不同,免疫压力不同,交易市场家禽来源不同等均会造成毒株之间的差异;四是不同地区候鸟的迁徙种类和数量亦存在差异,因此由候鸟传播给家禽的毒株亦存在差异。
研究发现,鸭的感染情况复杂,多以3种以上亚型的混合感染为主,这增加了防疫的难度。水禽被认为是禽流感病毒的储存库和传染源,可以通过公用水域与野鸟接触,隐形感染多种亚型病毒而不发病,为各亚型的基因重组提供了良好的载体,加快表面蛋白的变异,不断丰富内部基因的重配,最终又可以通过与陆禽的接触将变异的新型病毒传播给陆禽,引起新型病毒的流行与爆发。因此笔者认为水禽的防疫是禽流感防控的重点和难点,尽管水禽很少因感染禽流感病毒发生大规模的死亡,但因其在禽流感病毒传播和进化中的重要地位,该病的防控绝不能忽视对水禽的监测,同时应尽量避免野生水禽、家养水禽及其它野鸟与陆禽间的相互接触。
鸡的感染主要以H9N2亚型病毒为主,检出率位居首位,预示着未来防疫的艰巨性。一方面H9N2自身出现毒力增强的趋势。H9N2作为弱毒仅造成家禽呼吸道症状,通过进化变异成可致死禽类的强毒株,给养殖业造成巨大损失。另一方面该亚型毒株已突破宿主种间屏障,可感染人发病。早在1998-1999年香港、广州地区就发生4例人感染H9N2病例[10],随后江西、长沙、成都、云南、四川等地均出现H9N2感染人病例的报道[11-15]。人群H9N2血清学调查数据也验证了H9N2可感染人的结论[16]。不仅如此,H9N2还为1997年香港暴发的感染人H5N1、2013年国内暴发的感染人H7N9及2014年感染人H10N8等亚型流感病毒提供内部基因[17-19],说明潜在高致病的毒株可以与H9N2进行基因整合,并从H9N2的基因中获得更适合其生存和致病的基因,产生毒力更强,宿主范围更广的新型毒株,达到其进化的目的。因此笔者认为,在今后的防疫工作中,在兼顾高致病禽流感防控的同时,应加大对H9N2亚型的监控,养殖场应根据实际情况制定H9N2的免疫计划,而活禽市场应加强管理消毒,并细化禽类基本信息,以便溯源,防止该亚型的大流行和传染人事件的发生。
利益冲突:无
猜你喜欢
科学大观园(2022年2期)2022-01-23 11:05:15
汉语世界(The World of Chinese)(2017年3期)2017-10-09 10:58:52
启蒙(3-7岁)(2017年4期)2017-06-15 20:28:55
汉语世界(2017年3期)2017-06-05 15:01:42
动物医学进展(2015年10期)2015-12-07 05:46:18
家禽科学(2015年2期)2015-10-21 18:44:39
当代畜禽养殖业(2014年5期)2014-08-31 02:51:00
特产研究(2014年4期)2014-04-10 12:54:12
郑州大学学报(理学版)(2014年3期)2014-03-01 04:21:08
当代畜禽养殖业(2014年7期)2014-02-27 07:59:17
